
The Butterfly Effect: Mild Soil Pollution with Heavy Metals Elicits Major Biological Consequences in Cobalt-Sensitized Broad Bean Model Plants

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sample Characteristics
2.2. Experimental Design
2.3. Induction of Chlorophyll Morphosis in Vicia faba Plants with Co(NO3)2
2.4. Evaluation of Oxidative Stress in Plants Using Biochemical Markers
2.5. DNA Extraction and CDDP Analysis
2.6. Transcript Profiling Using Differential Display Technique
2.7. CDDP- and TDF-Based Dendrogram Construction
2.8. Statistical Analysis
3. Results
3.1. Chemical Characteristics of the Polluted Soil Samples
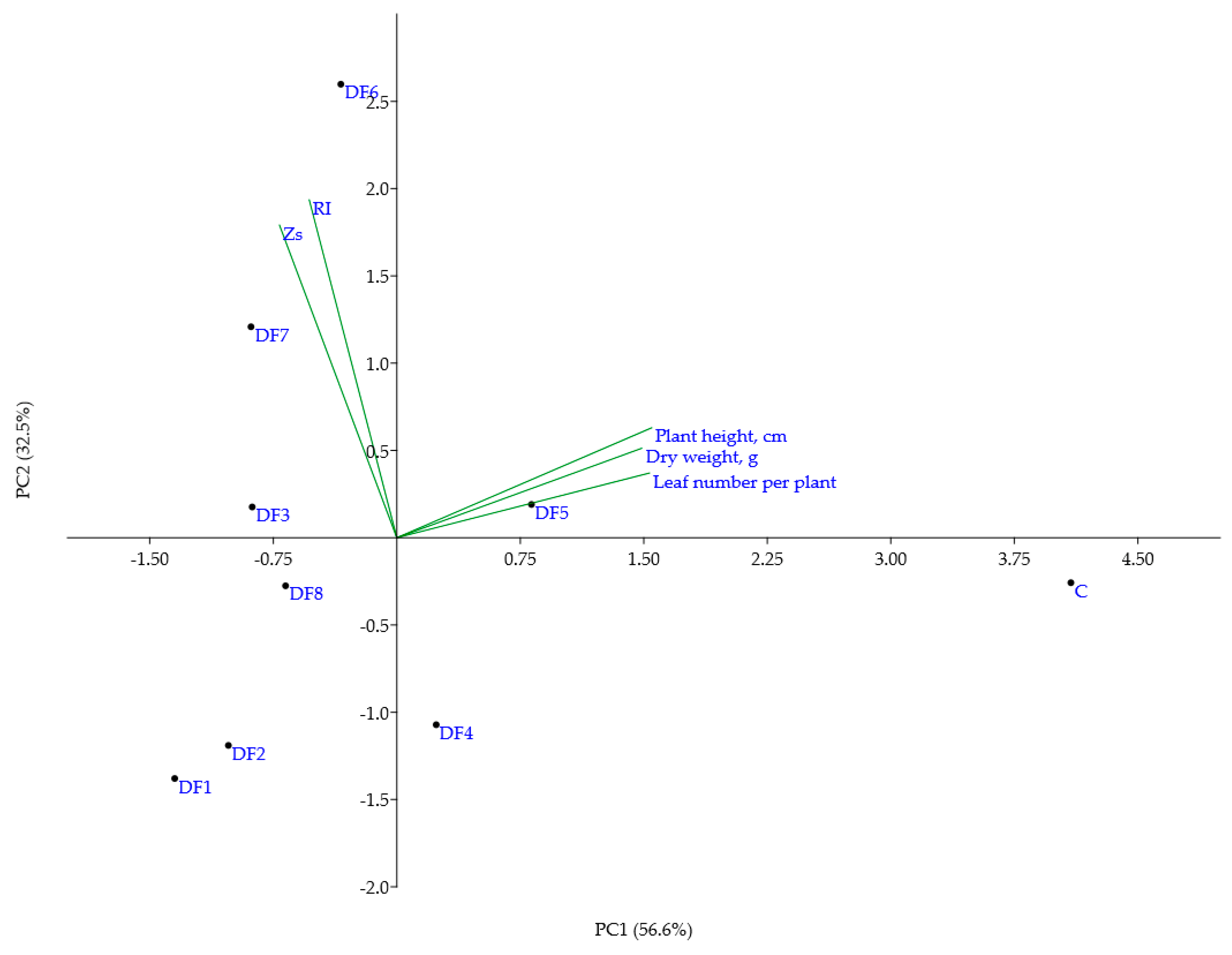
3.2. The Effect of Polluted Soils on Quantitative Parameters of V. faba Plants
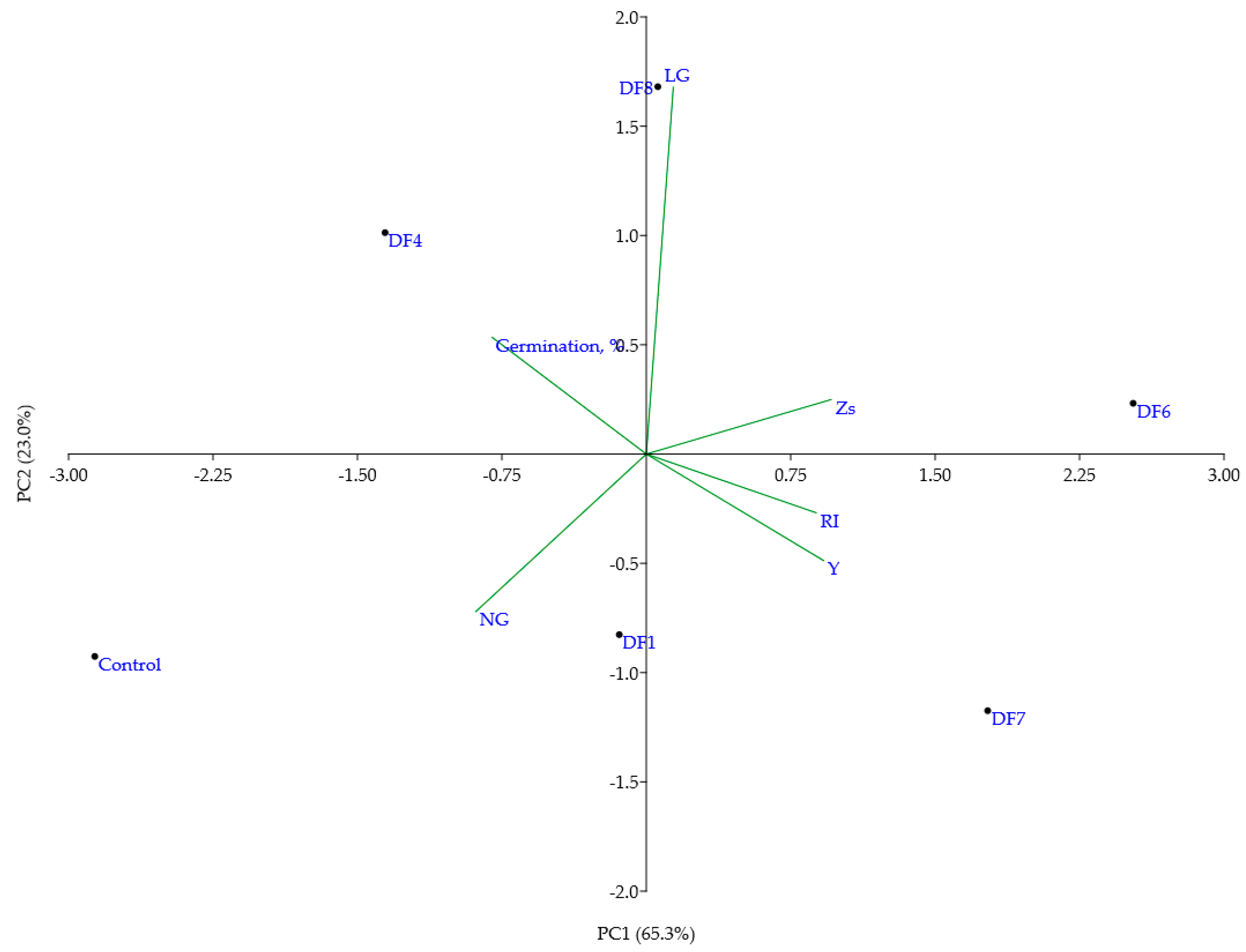
3.3. The Use of Co-Induced Chlorophyll Morphoses for the Evaluation of Soil Pollution
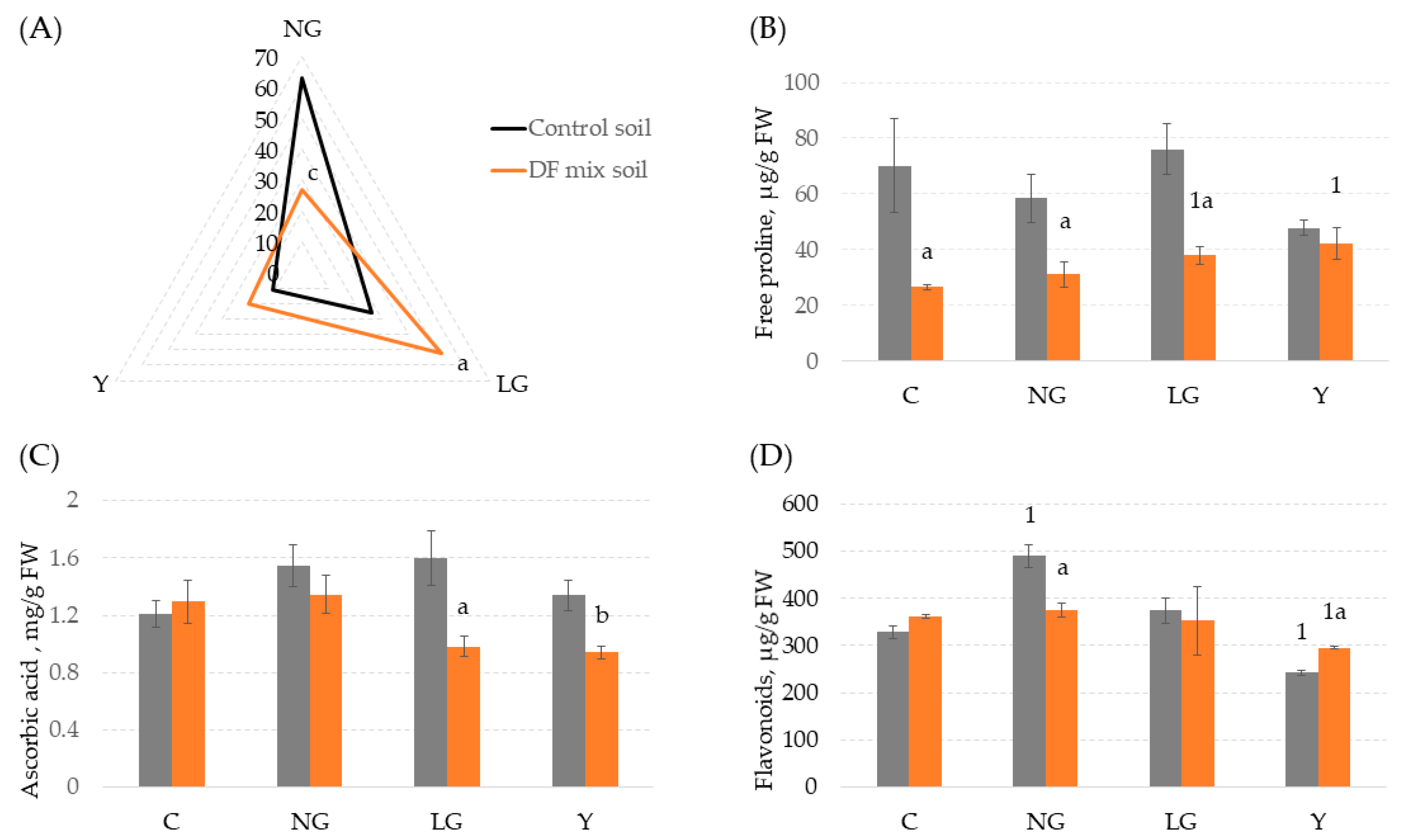
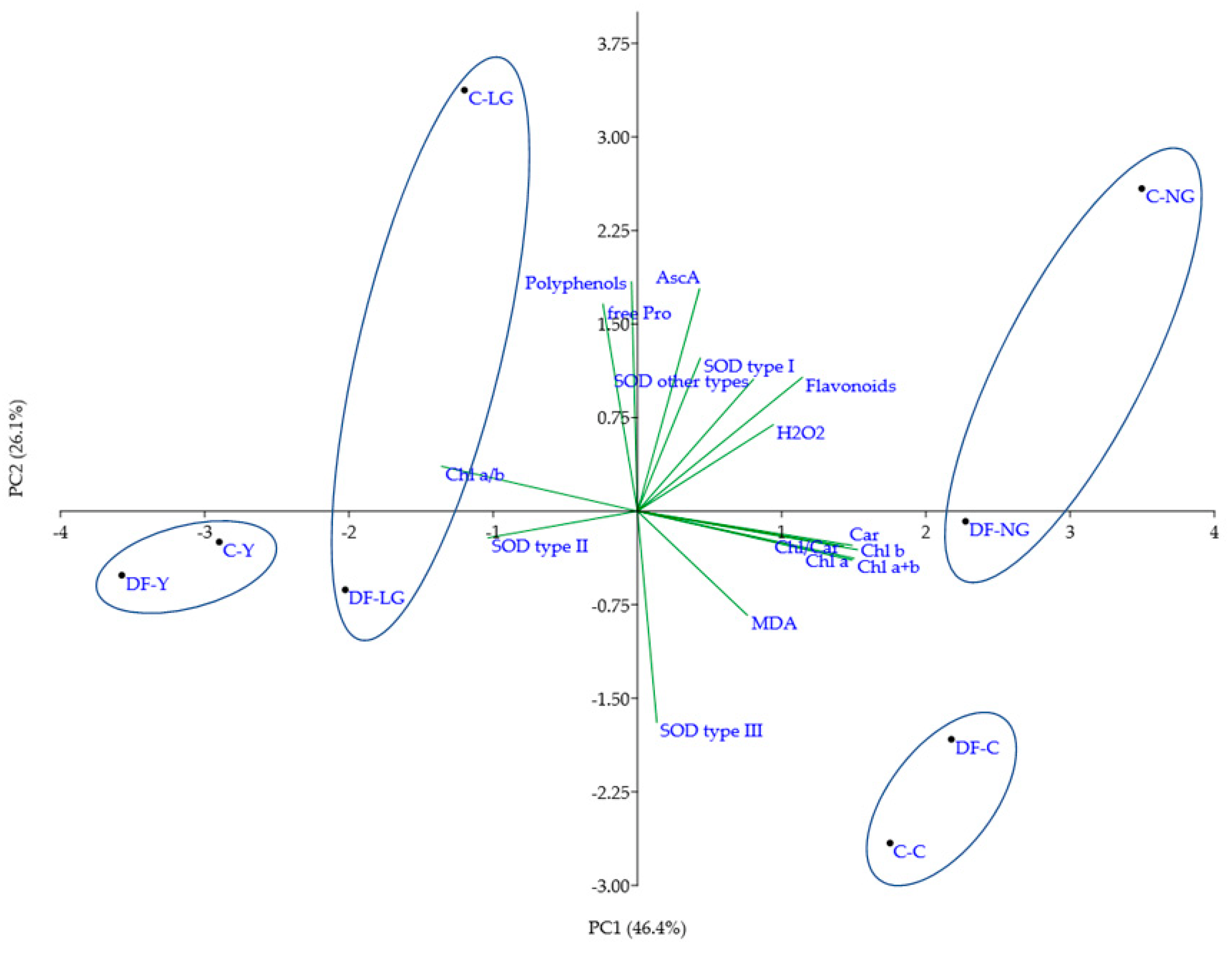
3.4. The Biochemical Response of Co-Treated V. faba Plants to Mild Heavy Metal-Induced Stress
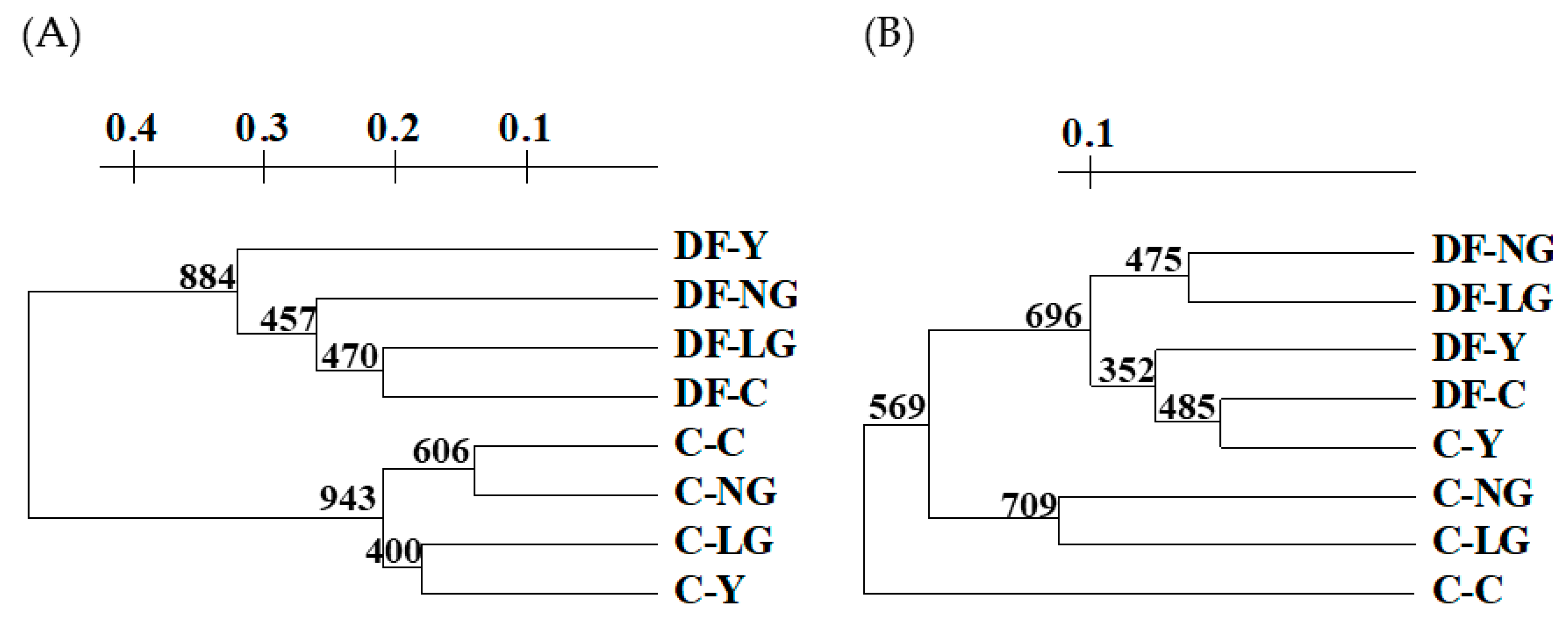
3.5. Evaluation of Heavy Metal-Induced Changes in V. faba Plants Using Molecular Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kosheleva, N.E.; Nikiforova, E.M. Long-Term Dynamics of Urban Soil Pollution with Heavy Metals in Moscow. Appl. Environ. Soil Sci. 2016, 2016, 5602795. [Google Scholar] [CrossRef] [Green Version]
- Manisalidis, I.; Stavropoulou, E.; Stavropoulos, A.; Bezirtzoglou, E. Environmental and Health Impacts of Air Pollution: A Review. Front. Public Health 2020, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieczorek, K.; Turek, A.; Szczesio, M.; Wolf, W.M. Comprehensive Evaluation of Metal Pollution in Urban Soils of a Post-Industrial City—A Case of Łódź, Poland. Molecules 2020, 25, 4350. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Khan, S. Heavy Metal Phytotoxicity: DNA Damage. In Cellular and Molecular Phytotoxicity of Heavy Metals; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Ghori, N.H.; Ghori, T.; Hayat, M.Q.; Imadi, S.R.; Gul, A.; Altay, V.; Ozturk, M. Heavy Metal Stress and Responses in Plants. Int. J. Environ. Sci. Technol. 2019, 16, 1807–1828. [Google Scholar] [CrossRef]
- Rehman, A.; Nazir, S.; Irchad, R.; Tahir, K.; Rehman, K.; Islam, R.; Wahab, Z. Toxicity of Heavy Metals in Plants and Animals and Their Uptake by Magnetic Iron Oxide Nanoparticles. J. Mol. Liq. 2021, 321, 114455. [Google Scholar] [CrossRef]
- Oka, G.A.; Thomas, L.; Lavkulich, L.M. Soil Assessment for Urban Agriculture: A Vancouver Case Study. J. Soil Sci. Plant Nutr. 2014, 14, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.H.; Mustafa, A.R.A.; El-Sheikh, A.A. Geochemistry and Spatial Distribution of Selected Heavy Metals in Surface Soil of Sohag, Egypt: A Multivariate Statistical and GIS Approach. Environ. Earth Sci. 2016, 75, 1257. [Google Scholar] [CrossRef]
- Khalid, S.; Shahid, M.; Natasha; Bibi, I.; Sarwar, T.; Shah, A.H.; Niazi, N.K. A Review of Environmental Contamination and Health Risk Assessment of Wastewater Use for Crop Irrigation with a Focus on Low and High-Income Countries. Int. J. Environ. Res. Public Health 2018, 15, 895. [Google Scholar] [CrossRef] [Green Version]
- Järup, L. Hazards of Heavy Metal Contamination. Br. Med. Bull. 2003, 68, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.; Yuan, L.; Jin, P.; Ding, B.; Qin, N.; Li, L.; Liu, X.; Wu, Z.; Zhao, G.; Deng, Y. A Clinical Study of the Effects of Lead Poisoning on the Intelligence and Neurobehavioral Abilities of Children. Theor. Biol. Med. Model 2013, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, Mechanism and Health Effects of Some Heavy Metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariani, R.L.; Martins Jorge, M.P.; Pereira, S.S.; Melione, L.P.; Carvalho-Oliveira, R.; Ma, T.H.; Nascimento Saldiva, P.H. Association between Micronuclei Frequency in Pollen Mother Cells of Tradescantia and Mortality Due to Cancer and Cardiovascular Diseases: A Preliminary Study in Sao José Dos Campos, Brazil. Environ. Pollut. 2009, 157, 1767–1770. [Google Scholar] [CrossRef] [PubMed]
- Korosteleva, N.v.; Rastyapina, O.A. Integration of Industrial Territories into the Urban Environment. IOP Conf. Ser. Mater. Sci. Eng. 2019, 687, 055017. [Google Scholar] [CrossRef]
- Khosrovyan, A.; DelValls, T.A.; Riba, I. Effects of Simulated CO2 Escape from Sediments on the Development of Midge Chironomus riparius. Aquat. Toxicol. 2014, 156, 230–239. [Google Scholar] [CrossRef]
- Bardina, T.V.; Chugunova, M.V.; Kapelkina, L.P.; Bardina, V.I.; Gerasimov, A.O. Ecological State Assessment of Urban Soils by Bioassay. Water Air Soil Pollut. 2022, 233, 7. [Google Scholar] [CrossRef]
- Fontanetti, C.S.; Nogarol, L.R.; de Souza, R.B.; Perez, D.G.; Maziviero, G.T. Bioindicators and Biomarkers in the Assessment of Soil Toxicity. In Soil Contamination; IntechOpen: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Khosrovyan, A.; Aghajanyan, E.; Avalyan, R.; Atoyants, A.; Sahakyan, L.; Gabrielyan, B.; Aroutiounian, R. Assessment of the Mutagenic Potential of the Water of an Urban River by Means of Two Tradescantia-Based Test Systems. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2022, 876–877, 503449. [Google Scholar] [CrossRef]
- Knasmüller, S.; Gottmann, E.; Steinkellner, H.; Fomin, A.; Pickl, C.; Paschke, A.; Göd, R.; Kundi, M. Detection of Genotoxic Effects of Heavy Metal Contaminated Soils with Plant Bioassays. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1998, 420, 37–48. [Google Scholar] [CrossRef]
- Maluszynska, J.; Juchimiuk, J. Plant Genotoxicity: A Molecular Cytogenetic Approach in Plant Bioassays. Arh. Hig. Rada. Toksikol. 2005, 56, 177–184. [Google Scholar]
- Meireles, J.R.C.; de Moraes Marcílio Cerqueira, E. Use of the Micronucleus Test on Tradescantia (Trad-MCN) to Evaluate the Genotoxic Effects of Air Pollution. In Air Pollution–New Developments; IntechOpen: London, UK, 2011. [Google Scholar] [CrossRef]
- Ma, T.H. Vicia Cytogenetic Tests for Environmental Mutagens. A report of the U.S. Environmental Protection Agency Gene-Tox Program. Mutat. Res. 1982, 99, 257–271. [Google Scholar] [CrossRef]
- Ma, T.H.; Xu, Z.; McConnell, H.; Rabago, E.V.; Arreola, G.A.; Zhang, H. The Improved Allium/Vicia Root Tip Micronucleus Assay for Clastogenicity of Environmental Pollutants. Mutat. Res. 1995, 334, 185–195. [Google Scholar] [CrossRef]
- Arya, S.K.; Mukherjee, A. Sensitivity of Allium cepa and Vicia faba towards Cadmium Toxicity. J. Soil Sci. Plant Nutr. 2014, 14, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yuan, S.; Liu, X.; Zhou, X.; Zhou, Y.; Song, Y. Genotoxicity Response of Vicia faba Seedlings to Cadmium in Soils as Characterized by Direct Soil Exposure and Micronucleus Test. Ecotoxicology 2020, 29, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Romero-Freire, A.; González, V.; Groenenberg, J.E.; Qiu, H.; Auffan, M.; Cotelle, S.; Giamberini, L. Cytotoxicity and Genotoxicity of Lanthanides for Vicia faba L. Are Mediated by Their Chemical Speciation in Different Exposure Media. Sci. Total Environ. 2021, 790, 148223. [Google Scholar] [CrossRef]
- Patlolla, A.K.; Berry, A.; May, L.; Tchounwou, P.B. Genotoxicity of Silver Nanoparticles in Vicia faba: A Pilot Study on the Environmental Monitoring of Nanoparticles. Int. J. Environ. Res. Public Health 2012, 9, 1649–1662. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Chen, H.; Liao, Y.; Ye, Z.; Li, M.; Klobučar, G. Ecotoxicity and Genotoxicity of Polystyrene Microplastics on Higher Plant Vicia faba. Environ. Pollut. 2019, 250, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Sta, C.; Ledoigt, G.; Ferjani, E.; Goupil, P. Exposure of Vicia faba to Sulcotrione Pesticide Induced Genotoxicity. Pestic. Biochem. Physiol. 2012, 103, 9–14. [Google Scholar] [CrossRef]
- Horozal, A.; Aksoy, Ö. Evaluation of the Genotoxicity of Some Standart and Eco-Friendly Detergents with Vicia faba. Caryologia 2020, 73, 129–139. [Google Scholar] [CrossRef]
- Rancelis, V.; Cesniene, T.; Zvingila, D.; Barysas, D.; Balciuniene, L.; Dapkuniene, S. Polymorphism of Response to Cobalt Excess in Individual Vicia faba Plants. Environ. Exp. Bot. 2006, 55, 221–234. [Google Scholar] [CrossRef]
- Rancelis, V.; Cesniene, T.; Kleizaite, V.; Zvingila, D.; Balciuniene, L. Influence of Cobalt Uptake by Vicia faba Seeds on Chlorophyll Morphosis Induction, SOD Polymorphism, and DNA Methylation. Environ. Toxicol. 2012, 27, 32–41. [Google Scholar] [CrossRef]
- Čėsnienė, T.; Kleizaitė, V.; Bondzinskaitė, S.; Taraškevičius, R.; Žvingila, D.; Šiukšta, R.; Rančelis, V. Metal Bioaccumulation and Mutagenesis in a Tradescantia Clone Following Long-Term Exposure to Soils from Urban Industrial Areas and Closed Landfills. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2017, 823, 65–72. [Google Scholar] [CrossRef]
- Velikova, V.; Loreto, F. On the Relationship between Isoprene Emission and Thermotolerance in Phragmites australis Leaves Exposed to High Temperatures and during the Recovery from a Heat Stress. Plant Cell Environ. 2005, 28, 318–327. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the Thiobarbituric Acid-Reactive-Substances Assay for Estimating Lipid Peroxidation in Plant Tissues Containing Anthocyanin and Other Interfering Compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Mitsui, A.; Ohta, T. Photooxidative Consumption and Photoreductive Formation of Ascorbic Acid in Green Leaves. Plant Cell Physiol. 1961, 2, 31–44. [Google Scholar] [CrossRef]
- Ábrahám, E.; Hourton-Cabassa, C.; Erdei, L.; Szabados, L. Methods for Determination of Proline in Plants. In Plant Stress Tolerance. Methods in Molecular Biology (Methods and Protocols); Humana Press: Totowa, NJ, USA, 2010; Volume 639, pp. 317–331. [Google Scholar] [CrossRef]
- Chew, Y.L.; Goh, J.K.; Lim, Y.Y. Assessment of in Vitro Antioxidant Capacity and Polyphenolic Composition of Selected Medicinal Herbs from Leguminosae Family in Peninsular Malaysia. Food Chem. 2009, 116, 13–18. [Google Scholar] [CrossRef]
- Minocha, R.; Martinez, G.; Lyons, B.; Long, S. Development of a Standardized Methodology for Quantifying Total Chlorophyll and Carotenoids from Foliage of Hardwood and Conifer Tree Species. Can. J. For. Res. 2009, 39, 849–861. [Google Scholar] [CrossRef]
- Kleizaite, V.; Cesniene, T.; Rancelis, V. The Use of Cobalt-Induced Chlorophyll Morphoses for Studying Co2+ Interactions with Cysteine and SOD. Plant Sci. 2004, 167, 1249–1256. [Google Scholar] [CrossRef]
- Doyle, J.; Doyle, J. Isolation of Plant DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Collard, B.C.Y.; Mackill, D.J. Conserved DNA-Derived Polymorphism (CDDP): A Simple and Novel Method for Generating DNA Markers in Plants. Plant Mol. Biol. Rep. 2009, 27, 558–562. [Google Scholar] [CrossRef]
- Šiukšta, R.; Vaitkūnienė, V.; Rančelis, V. Is Auxin Involved in the Induction of Genetic Instability in Barley Homeotic Double Mutants? Planta 2018, 247, 483–498. [Google Scholar] [CrossRef]
- Benbouza, H.; Jacquemin, J.-M.; Baudoin, J.-P.; Guy, M. Optimization of a Reliable, Fast, Cheap and Sensitive Silver Staining Method to Detect SSR Markers in Polyacrylamide Gels. Biotechnol. Agron. Soc. Environ. 2006, 10, 77–81. [Google Scholar]
- Nei, M.; Li, W.H. Mathematical Model for Studying Genetic Variation in Terms of Restriction Endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Peer, Y.; de Wachter, R. TREECON for Windows: A Software Package for the Construction and Drawing of Evolutionary Trees for the Microsoft Windows Environment. Bioinformatics 1994, 10, 569–570. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Chaves, L.H.G.; Estrela, M.A.; De Souza, R.S. Effect on Plant Growth and Heavy Metal Accumulation by Sunflower. J. Phytol. 2011, 3, 4–9. [Google Scholar]
- Kobaissi, A.N.; Kanso, A.A.; Kanbar, H.J. Translocation of Heavy Metals in Zea mays L. Treated with Wastewater and Consequences on Morphophysiological Aspects. Rev. Int. Contam. Ambient. 2014, 30, 297–305. [Google Scholar]
- Arif, N.; Yadav, V.; Singh, S.; Singh, S.; Ahmad, P.; Mishra, R.K.; Sharma, S.; Tripathi, D.K.; Dubey, N.K.; Chauhan, D.K. Influence of High and Low Levels of Plant-Beneficial Heavy Metal Ions on Plant Growth and Development. Front. Environ. Sci. 2016, 4, 69. [Google Scholar] [CrossRef]
- Ullah, S.; Khan, J.; Hayat, K.; Elateeq, A.A.; Salam, U.; Yu, B.; Ma, Y.; Wang, H.; Tang, Z.H. Comparative Study of Growth, Cadmium Accumulation and Tolerance of Three Chickpea (Cicer arietinum L.) Cultivars. Plants 2020, 9, 310. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Guan, J.; Liang, Q.; Zhang, X.; Hu, H.; Zhang, J. Effects of Cadmium Stress on Growth and Physiological Characteristics of Sassafras Seedlings. Sci. Rep. 2021, 11, 9913. [Google Scholar] [CrossRef]
- Majer, B.J.; Tscherko, D.; Paschke, A.; Wennrich, R.; Kundi, M.; Kandeler, E.; Knasmüller, S. Effects of Heavy Metal Contamination of Soils on Micronucleus Induction in Tradescantia and on Microbial Enzyme Activities: A Comparative Investigation. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2002, 515, 111–124. [Google Scholar] [CrossRef]
- Stapulionytė, A.; Kleizaitė, V.; Šiukšta, R.; Žvingila, D.; Taraškevičius, R.; Čėsnienė, T. Cyto/Genotoxicological Evaluation of Hot Spots of Soil Pollution Using Allium Bioassays in Relation to Geochemistry. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2019, 842, 102–110. [Google Scholar] [CrossRef]
- Šiukšta, R.; Bondzinskaitė, S.; Kleizaitė, V.; Žvingila, D.; Taraškevičius, R.; Mockeliūnas, L.; Stapulionytė, A.; Mak, K.; Čėsnienė, T. Response of Tradescantia Plants to Oxidative Stress Induced by Heavy Metal Pollution of Soils from Industrial Areas. Environ. Sci. Pollut. Res. 2019, 26, 44–61. [Google Scholar] [CrossRef] [PubMed]
- An, Y.J.; Kim, Y.M.; Kwon, T.I.; Jeong, S.W. Combined Effect of Copper, Cadmium, and Lead upon Cucumis sativus Growth and Bioaccumulation. Sci. Total Environ. 2004, 326, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, K.; Gill, R.A.; Islam, F.; Farooq, M.A.; Wang, J.; Zhou, W. Ecotoxicological and Interactive Effects of Copper and Chromium on Physiochemical, Ultrastructural, and Molecular Profiling in Brassica napus L. Biomed. Res. Int. 2018, 2018, 9248123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lwalaba, J.L.W.; Louis, L.T.; Zvobgo, G.; Fu, L.; Mwamba, T.M.; Mukobo Mundende, R.P.; Zhang, G. Copper Alleviates Cobalt Toxicity in Barley by Antagonistic Interaction of the Two Metals. Ecotoxicol. Environ. Saf. 2019, 180, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Małecka, A.; Konkolewska, A.; Hanć, A.; Ciszewska, L.; Staszak, A.M.; Jarmuszkiewicz, W.; Ratajczak, E. Activation of Antioxidative and Detoxificative Systems in Brassica juncea L. Plants against the Toxicity of Heavy Metals. Sci. Rep. 2021, 11, 22345. [Google Scholar] [CrossRef] [PubMed]
- Küpper, H.; Andresen, E. Mechanisms of Metal Toxicity in Plants. Metallomics 2016, 8, 269–285. [Google Scholar] [CrossRef]
- White, P.A.; Claxton, L.D. Mutagens in Contaminated Soil: A Review. Mutat. Res. Rev. Mutat. Res. 2004, 567, 227–345. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.K. Heavy Metals Toxicity in Plants: An Overview on the Role of Glutathione and Phytochelatins in Heavy Metal Stress Tolerance of Plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy Metal Stress and Some Mechanisms of Plant Defense Response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M. Dual Role of Metallic Trace Elements in Stress Biology—From Negative to Beneficial Impact on Plants. Int. J. Mol. Sci. 2019, 20, 3117. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Wei, X.; Ling, J.; Chen, J. Cobalt: An Essential Micronutrient for Plant Growth? Front. Plant Sci. 2021, 12, 2370. [Google Scholar] [CrossRef] [PubMed]
- Daulta, R.; Prakash, M.; Goyal, S. Metal Content in Soils of Northern India and Crop Response: A Review. Int. J. Environ. Sci. Technol. 2022. [Google Scholar] [CrossRef]
- Ul Hassan, Z.; Ali, S.; Rizwan, M.; Hussain, A.; Akbar, Z.; Rasool, N.; Abbas, F. Role of Zinc in Alleviating Heavy Metal Stress. In Essential Plant Nutrients: Uptake, Use Efficiency, and Management; Springer: Cham, Switzerland, 2017; pp. 351–366. [Google Scholar] [CrossRef]
- Elazab, D.S.; Abdel-Wahab, D.A.; El-Mahdy, M.T. Iron and Zinc Supplies Mitigate Cadmium Toxicity in Micropropagated Banana (Musa spp.). Plant Cell Tissue Organ Cult. 2021, 145, 367–377. [Google Scholar] [CrossRef]
- Ince, N.H.; Dirilgen, N.; Apikyan, I.G.; Tezcanli, G.; Üstün, B. Assessment of Toxic Interactions of Heavy Metals in Binary Mixtures: A Statistical Approach. Arch. Environ. Contam. Toxicol. 1999, 36, 365–372. [Google Scholar] [CrossRef]
- Palit, S.; Sharma, A.; Talukder, G. Effects of Cobalt on Plants. Bot. Rev. 1994, 60, 149–181. [Google Scholar] [CrossRef]
- Salinitro, M.; Hoogerwerf, S.; Casolari, S.; Zappi, A.; Melucci, D.; Tassoni, A. Production of Antioxidant Molecules in Polygonum aviculare (L.) and Senecio vulgaris (L.) under Metal Stress: A Possible Tool in the Evaluation of Plant Metal Tolerance. Int. J. Mol. Sci. 2020, 21, 7317. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, Y.; Xu, Z.; Huang, H.; Zhou, J.; Yang, G. Morphological and Physiological Changes of Broussonetia papyrifera Seedlings in Cadmium Contaminated Soil. Plants 2020, 9, 1698. [Google Scholar] [CrossRef]
- Alsherif, E.A.; Al-Shaikh, T.M.; Abdelgawad, H. Heavy Metal Effects on Biodiversity and Stress Responses of Plants Inhabiting Contaminated Soil in Khulais, Saudi Arabia. Biology 2022, 11, 164. [Google Scholar] [CrossRef]
- Sgobba, A.; Paradiso, A.; Dipierro, S.; de Gara, L.; de Pinto, M.C. Changes in Antioxidants Are Critical in Determining Cell Responses to Short- and Long-Term Heat Stress. Physiol. Plant. 2015, 153, 68–78. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, H.; Yu, F.; Hu, B.; Jia, Y.; Sha, H.; Zhao, H. Differential Activity of the Antioxidant Defence System and Alterations in the Accumulation of Osmolyte and Reactive Oxygen Species under Drought Stress and Recovery in Rice (Oryza sativa L.) Tillering. Sci. Rep. 2019, 9, 8543. [Google Scholar] [CrossRef] [PubMed]
- Lau, O.L.; Yang, S.F. Inhibition of Ethylene Production by Cobaltous Ion. Plant Physiol. 1976, 58, 114–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thao, N.P.; Khan, M.I.R.; Anh Thu, N.B.; Thi Hoang, X.L.; Asgher, M.; Khan, N.A.; Tran, L.S.P. Role of Ethylene and Its Cross Talk with Other Signaling Molecules in Plant Responses to Heavy Metal Stress. Plant Physiol. 2015, 169, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keunen, E.; Schellingen, K.; Vangronsveld, J.; Cuypers, A. Ethylene and Metal Stress: Small Molecule, Big Impact. Front. Plant Sci. 2016, 7, 23. [Google Scholar] [CrossRef] [Green Version]
- Saini, S.; Kaur, N.; Pati, P.K. Phytohormones: Key Players in the Modulation of Heavy Metal Stress Tolerance in Plants. Ecotoxicol. Environ. Saf. 2021, 223, 112578. [Google Scholar] [CrossRef]
- Sun, P.; Tian, Q.Y.; Chen, J.; Zhang, W.H. Aluminium-Induced Inhibition of Root Elongation in Arabidopsis Is Mediated by Ethylene and Auxin. J. Exp. Bot. 2010, 61, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Chmielowska-Bąk, J.; Gzyl, J.; Rucińska-Sobkowiak, R.; Arasimowicz-Jelonek, M.; Deckert, J. The New Insights into Cadmium Sensing. Front. Plant Sci. 2014, 5, 245. [Google Scholar] [CrossRef]
- Tamás, L.; Mistrík, I.; Huttová, J.; Halušková, L.; Valentovičová, K.; Zelinová, V. Role of Reactive Oxygen Species-Generating Enzymes and Hydrogen Peroxide during Cadmium, Mercury and Osmotic Stresses in Barley Root Tip. Planta 2010, 231, 221–231. [Google Scholar] [CrossRef]
- De Silva, N.D.G.; Cholewa, E.; Ryser, P. Effects of Combined Drought and Heavy Metal Stresses on Xylem Structure and Hydraulic Conductivity in Red Maple (Acer rubrum L.). J. Exp. Bot. 2012, 63, 5957–5966. [Google Scholar] [CrossRef] [Green Version]
- Rucińska-Sobkowiak, R. Water Relations in Plants Subjected to Heavy Metal Stresses. Acta Physiol. Plant. 2016, 38, 257. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cai, X.; Xu, C.; Wang, Q.; Dai, S. Drought-Responsive Mechanisms in Plant Leaves Revealed by Proteomics. Int. J. Mol. Sci. 2016, 17, 1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, F.; Zhang, F. Cell Cycle Regulation in the Plant Response to Stress. Front. Plant Sci. 2020, 10, 1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umate, P.; Schwenkert, S.; Karbat, I.; Dal Bosco, C.; Mlcòchová, L.; Volz, S.; Zer, H.; Herrmann, R.G.; Ohad, I.; Meurer, J. Deletion of PsbM in Tobacco Alters the QB Site Properties and the Electron Flow within Photosystem II. J. Biol. Chem. 2018, 293, 11649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nan, H.; Ludlow, R.A.; Lu, M.; An, H. Genome-Wide Analysis of Dof Genes and Their Response to Abiotic Stress in Rose (Rosa chinensis). Front. Genet. 2021, 12, 168. [Google Scholar] [CrossRef]
- Ghatak, A.; Chaturvedi, P.; Weckwerth, W. Cereal Crop Proteomics: Systemic Analysis of Crop Drought Stress Responses towards Marker-Assisted Selection Breeding. Front. Plant Sci. 2017, 8, 757. [Google Scholar] [CrossRef] [Green Version]
- Ru, J.N.; Hou, Z.H.; Zheng, L.; Zhao, Q.; Wang, F.Z.; Chen, J.; Zhou, Y.-B.; Chen, M.; Ma, Y.Z.; Xi, Y.J.; et al. Genome-Wide Analysis of DEAD-Box RNA Helicase Family in Wheat (Triticum aestivum) and Functional Identification of TaDEAD-Box57 in Abiotic Stress Responses. Front. Plant Sci. 2021, 12, 2714. [Google Scholar] [CrossRef]
- Acosta-Pérez, P.; Camacho-Zamora, B.D.; Espinoza-Sánchez, E.A.; Gutiérrez-Soto, G.; Zavala-García, F.; Abraham-Juárez, M.J.; Sinagawa-García, S.R. Characterization of Trehalose-6-Phosphate Synthase and Trehalose-6-Phosphate Phosphatase Genes and Analysis of Its Differential Expression in Maize (Zea mays) Seedlings under Drought Stress. Plants 2020, 9, 315. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Hu, Y.; Jin, G.; Lei, P.; Sang, L.; Luo, Q.; Liu, Z.; Guan, F.; Meng, F.; Zhao, X. Physiological and Proteomic Responses to Drought in Leaves of Amygdalus mira (Koehne) Yü et Lu. Front. Plant Sci. 2021, 12, 876. [Google Scholar] [CrossRef]
- Islam, A.; Leung, S.; Nikmatullah, A.; Dijkwel, P.P.; McManus, M.T. Kunitz Proteinase Inhibitors Limit Water Stress Responses in White Clover (Trifolium repens L.) Plants. Front. Plant Sci. 2017, 8, 1683. [Google Scholar] [CrossRef] [Green Version]
- Xu, C. Physiological and Molecular Response in Soybean to Drought and Heat Stresses. Ph.D. Thesis, Purdue University, West Lafayette, IN, USA, 2018. [Google Scholar]
- Fu, J.; Momčilović, I.; Vara Prasad, P.V. Roles of Protein Synthesis Elongation Factor EF-Tu in Heat Tolerance in Plants. J. Bot 2012, 2012, 835836. [Google Scholar] [CrossRef] [Green Version]
- Fan, T.; Lv, T.; Xie, C.; Zhou, Y.; Tian, C. Genome-Wide Analysis of the IQM Gene Family in Rice (Oryza sativa L.). Plants 2021, 10, 1949. [Google Scholar] [CrossRef] [PubMed]
- Atienzar, F.A.; Venier, P.; Jha, A.N.; Depledge, M.H. Evaluation of the Random Amplified Polymorphic DNA (RAPD) Assay for the Detection of DNA Damage and Mutations. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2002, 521, 151–163. [Google Scholar] [CrossRef]
- Mohsenzadeh, S.; Shahrtash, M.; Mohabatkar, H. Cadmium-Induced Genotoxicity Detected by the Random Amplification of Polymorphism DNA in the Maize Seedling Roots. J. Cell Mol. Res. 2010, 2, 42–48. [Google Scholar]
- Correia, S.; Matos, M.; Ferreira, V.; Martins, N.; Gonçalves, S.; Romano, A.; Pinto-Carnide, O. Molecular Instability Induced by Aluminum Stress in Plantago Species. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 770, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Ozyigit, I.I.; Dogan, I.; Igdelioglu, S.; Filiz, E.; Karadeniz, S.; Uzunova, Z. Screening of Damage Induced by Lead (Pb) in Rye (Secale cereale L.)—A Genetic and Physiological Approach. Biotechnol. Biotechnol. Equip. 2016, 30, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Taspinar, M.S.; Aydin, M.; Sigmaz, B.; Yagci, S.; Arslan, E.; Agar, G. Aluminum-Induced Changes on DNA Damage, DNA Methylation and LTR Retrotransposon Polymorphism in Maize. Arab. J. Sci. Eng. 2017, 43, 123–131. [Google Scholar] [CrossRef]
- Ozyigit, I.I.; Kaval, A.; Altundag Cakir, E.; Vardar, F. DNA Fingerprinting and Assessment of Some Physiological Changes in Al-Induced Bryophyllum daigremontianum Clones. Mol. Biol. Rep. 2019, 46, 2703–2711. [Google Scholar] [CrossRef]
- Poczai, P.; Varga, I.; Laos, M.; Cseh, A.; Bell, N.; Valkonen, J.P.T.; Hyvönen, J. Advances in Plant Gene-Targeted and Functional Markers: A Review. Plant Methods 2013, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Lau, W.C.P.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.; Latif, M.A.; Ramli, A. Review of Functional Markers for Improving Cooking, Eating, and the Nutritional Qualities of Rice. Front. Plant Sci. 2015, 6, 832. [Google Scholar] [CrossRef] [Green Version]
- Beyersmann, D.; Hartwig, A. The Genetic Toxicology of Cobalt. Toxicol. Appl. Pharmacol. 1992, 115, 137–145. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Sample | Zs a | RI b | Elements Which Exceed Limits According to Lithuanian Hygienic Norm HN 60:2015 c | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Value | Hazard Level | Value | Risk | |||||||
| C | - | - | 2 | low | ||||||
| DF1 | 26 | average hazard | 12 | low | Mo1.1 | Pb1.2 | ||||
| DF2 | 30 | average hazard | 11 | low | Mo1.5 | Zn1.1 | ||||
| DF3 | 37 | hazard | 80 | low | Cd1.9 | |||||
| DF4 | 17 | average hazard | 9 | low | ||||||
| DF5 | 65 | hazard | 14 | low | Cr1.1 | Mo6.1 | ||||
| DF6 | 104 | hazard | 129 | low | Cr1.2 | Hg2.8 | Mo7.2 | Pb1.3 | Zn2.0 | |
| DF7 | 83 | hazard | 83 | low | Cd1.6 | Cr1.1 | Mo4.6 | Pb1.2 | Zn1.7 | |
| DF8 | 67 | hazard | 14 | low | Cr1.0 | Mo6.6 | ||||
| DF soil mix | 67 | hazard | 14 | low | Cr1.2 | Zn1.1 | ||||
| Group | TDF | TDF Length, nt | Intensity of Polymorphic TDF Bands | Homologous Gene | Sequence Homology, % | E value | Accession Number | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control Soil | DF Soil Mix | |||||||||||||
| C | NG | LG | Y | C | NG | LG | Y | |||||||
| I | N4 | 350 | 1 | 1 ↓ | 1 | 1 ↓ | 0 | 0 | 0 | 0 | Medicago truncatula carboxy-terminal region remorin | 96.00 | 2 × 10−9 | XM_013589416.1 |
| N13 | 300 | 1 | 1 ↓↓ | 1 | 1 ↓ | 1 | 0 | 0 | 0 | PREDICTED: Medicago truncatula cell division control protein 2 homolog | 87.15 | 2 × 10−52 | XM_013601820.2 | |
| N18 | 200 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | Lupinus angustifolius cultivar Tanjil chromosome LG-04 | 77.59 | 0.44 | CP023116.1 | |
| N29 | 220 | 1 ↓ | 1 ↓ | 1 ↓ | 1 | 0 | 0 | 0 | 0 | Chionochloa rigida subsp. amara PsbM (psbM) gene | 90.91 | 0.053 | GQ305171.1 | |
| N37/N41 | 300 | 1 | 1 ↓ | 1 ↓ | 1 | 0 | 0 | 0 | 0 | Pisum sativum PSI light-harvesting antenna chlorophyll a/b-binding protein (lhcA-P4) | 91.44 | 6 × 10−65 | AF002248.3 | |
| N49 | 150 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | PREDICTED: Medicago truncatula cyclic dof factor 2 | 91.67 | 2 × 10−9 | XM_003618459.3 | |
| N33 | 240 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | Pisum sativum DEAD box RNA helicase | 90.34 | 1 × 10−55 | AY167671.1 | |
| II | N17 | 370 | 0 | 0 | 0 | 0 | 1 | 1 | 1 ↓ | 1 ↓ | Vicia villosa chloroplast NADH dehydrogenase subunit 4-like | 96.92 | 4 × 10−133 | KT457043.1 |
| N47 | 320 | 1 ↓ | 0 | 0 | 0 | 1 | 1 | 1 | 1 ↓ | Medicago truncatula trehalose-6-phosphate synthase domain protein | 84.42 | 4 × 10−32 | XM_003630929.3 | |
| N54 | 180 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | PREDICTED: Cicer arietinum uncharacterized | 86.96 | 3 × 10−21 | XM_004504383.3 | |
| III | N35 | 440 | 1 | 1 | 1 | 1 | 1 ↓ | 1 ↓ | 1 ↓ | 1 ↓ | PREDICTED: Medicago truncatula kunitz trypsin inhibitor 2 | 84.09 | 1 × 10−83 | XM_003620121.3 |
| N36 | 400 | 1 ↓ | 1 ↓ | 1 ↓ | 1 ↓ | 1 | 1 | 1 | 1 | PREDICTED: Medicago truncatula pentatricopeptide repeat-containing protein At1g71460, chloroplastic | 82.20 | 4 × 10−78 | XM_003604187.3 | |
| IV | N32 | 250 | 1 | 0 | 0 | 0 | 1 ↓ | 0 | 0 | 0 | PREDICTED: Medicago truncatula elongation factor Tu, chloroplastic | 77.91 | 0.001 | XM_003601112.3 |
| N40 | 350 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | PREDICTED: Medicago truncatula IQ domain-containing protein IQM1 | 84.62 | 7 × 10−8 | XM_013603017.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šiukšta, R.; Pukenytė, V.; Kleizaitė, V.; Bondzinskaitė, S.; Čėsnienė, T. The Butterfly Effect: Mild Soil Pollution with Heavy Metals Elicits Major Biological Consequences in Cobalt-Sensitized Broad Bean Model Plants. Antioxidants 2022, 11, 793. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11040793
Šiukšta R, Pukenytė V, Kleizaitė V, Bondzinskaitė S, Čėsnienė T. The Butterfly Effect: Mild Soil Pollution with Heavy Metals Elicits Major Biological Consequences in Cobalt-Sensitized Broad Bean Model Plants. Antioxidants. 2022; 11(4):793. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11040793
Chicago/Turabian StyleŠiukšta, Raimondas, Vėjūnė Pukenytė, Violeta Kleizaitė, Skaistė Bondzinskaitė, and Tatjana Čėsnienė. 2022. "The Butterfly Effect: Mild Soil Pollution with Heavy Metals Elicits Major Biological Consequences in Cobalt-Sensitized Broad Bean Model Plants" Antioxidants 11, no. 4: 793. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11040793