
A Whiff of Sulfur: One Wind a Day Keeps the Doctor Away
by
 , , and
, , and
Eduard Tiganescu
 ,
,
Markus Alexander Lämmermann
,
Yannick Ney
,
Ahmad Yaman Abdin
 ,
,
Muhammad Jawad Nasim
* and
Claus Jacob
* Division of Bioorganic Chemistry, School of Pharmacy, Saarland University, D-66123 Saarbruecken, Germany
*
Authors to whom correspondence should be addressed.
Antioxidants 2022, 11(6), 1036; https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11061036
Submission received: 18 April 2022
/
Revised: 12 May 2022
/
Accepted: 18 May 2022
/
Published: 24 May 2022
(This article belongs to the Special Issue Reactive Sulfur Species (RSS) in Physiological and Pathological Conditions and in Therapy)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Reactive Sulfur Species (RSS), such as allicin from garlic or sulforaphane from broccoli, are fre-quently associated with biological activities and possible health benefits in animals and humans. Among these Organic Sulfur Compounds (OSCs) found in many plants and fungi, the Volatile Sulfur Compounds (VSCs) feature prominently, not only because of their often-pungent smell, but also because they are able to access places which solids and solutions cannot reach that easily. Indeed, inorganic RSS such as hydrogen sulfide (H2S) and sulfur dioxide (SO2) can be used to lit-erally fumigate entire rooms and areas. Similarly, metabolites of garlic, such as allyl methyl sulfide (AMS), are formed metabolically in humans in lower concentrations and reach the airways from inside the body as part of one’s breath. Curiously, H2S is also formed in the gastrointestinal tract by gut bacteria, and the question of if and for which purpose this gas then crosses the barriers and enters the body is indeed a delicate matter for equally delicate studies. In any case, nature is surprisingly rich in such VSCs, as fruits (for instance, the infamous durian) demonstrate, and therefore these VSCs represent a promising group of compounds for further studies.
1. Introduction
Folk medicine has long been interested in plants and fungi rich in sulfur [1,2,3]. During the centuries and across the globe, it has ascribed several potential health benefits to the active ingredients contained in plants such as garlic, onions, mustard, and broccoli, and fungi such as Shiitake [2,4,5,6]. Indeed, at closer inspection, one finds many popular proverbs, for instance across Germany, which link garlic and onions to a healthy digestion and cardiovascular benefits. In Bavaria, home to the fairytale caste Neuschwanstein, for instance, there is an old saying that “if the arse is singing the heart is swinging”, whereas in the Saarland, home to the acclaimed Saarland Hurricanes (Canes) football team, the advice is “to have sunshine in the heart and onions in the belly, then you can breathe freely and smelly” (and not only via the lungs). Although some of this may sound rather offensive initially—just as the battle-cry of the Canes (Let’s go offense, let’s go)—we shall return to some of the possible health benefits associated with such (smelly) metabolites, for instance in the cardiovascular system, in the following sections.
Indeed, the natural products containing sulfur are rather special [7]. Different scientific disciplines have divided such sulfur compounds into specific groups—for instance, the Organic Sulfur Compounds (OSCs) in a more chemical context and the Reactive Sulfur Species (RSS) in a more biological one. Among these OSCs/RSS, the Volatile Sulfur Compounds (VSCs) stand out as they tend to combine high reactivity and hence biological activity with a unique air-bound bioavailability [8,9,10]. No doubt, there are quite a few anecdotes circulating within the scientific community about allicin from garlic fumigating entire incubators and thence affecting cell-based experiments in every corner of the instrument [11,12,13].
In this article, we shall therefore consider different activities and roles ascribed to volatile RSS. We will start with the more prominent activities of H2S as a gaseous transmitter in the human body on par with nitric oxide (●NO) and the uses of SO2 as fumigant and then turn to some of the more exotic and less(er) studied actions of natural substances, such as allicin and its metabolites.
2. Inorganic VSCs
In the periodic table, sulfur is found together with oxygen, selenium, tellurium, and polonium in Group 16, elements often referred to as the chalk generating chalcogens. Unlike most other elements, sulfur occurs in many different oxidation states ranging from −2 to +6, and indeed its inorganic chemistry alone may fill entire bookshelves and is quite colorful as far as reactivities and biological activities are concerned. As an appetizer for further reading, we would like to highlight topics such as S-thiolation and O-sulfation of proteins and enzymes, which may be considered as processes similar to phosphorylation. There is also a barrage of—less(er) studied—inorganic RSS, which contain highly reactive substances, such as hypothiocyanite (OSCN−) and thiocyanates (SCN−), and have been investigated recently by colleagues such as Michael T. Ashby [14,15,16].
The most prominent and indeed also simple volatile inorganic RSS are H2S (melting point −85.49 °C and boiling point −60.33 °C, at atmospheric pressure), with sulfur in its reduced form (oxidation state −2), and SO2 (melting point −75.5 °C, boiling point −10.05 °C) with sulfur in oxidized form (oxidation state +4). Please note that SO3 with sulfur in oxidation state +6 is not that volatile at room temperature (melting point 16.90 °C, boiling point 44.50 °C).
2.1. Hydrogen Sulfide (H2S)
Hydrogen sulfide may affect animals and humans via quite a few avenues and in quite a few ways. Firstly, H2S occurs in our environment, for instance near volcanos and in mineral waters, and as such may be inhaled and also taken as a bath or orally [17,18]. H2S is lethal at higher concentrations exceeding 500–1000 ppm (formally corresponding to 14.7–29.4 mM), whilst at concentrations of 10–500 ppm (0.3–14.7 mM), it affects the respiratory system and symptoms of exposure range from rhinitis to acute respiratory failure [19]. H2S is often considered beneficial to human health at lower concentrations and dissolved in water. Bad Nenndorf located in the district of Schaumburg in the State of Lower Saxony in Germany is representative of such a natural mineral sulfur spring with a total sulfur content of 137 ppm, including an H2S content ranging from 4–10 ppm (100–300 µM) [20]. Such sulfur-rich water has been employed extensively for the treatment of various types of rheumatic diseases, including degenerative, inflammatory, and soft tissue rheumatism (e.g., fibromyalgia), and also against inflammatory skin diseases, such as eczema and neurodermatitis [20]. Similarly, the Varna basin mineral water in the northern Black Sea region in Bulgaria, which among other sulfur species also contains dissolved H2S and soluble sulfides in concentrations ranging from 1.7 to 2.8 ppm (49.9–82.1 µM), has been reported to improve total glutathione and total thiol levels in healthy volunteers with increased expression of γ-glutamyl-cysteinyl ligase (GCL) and soluble intercellular adhesion molecule-1 (sICAM-1) genes [21,22].
Notably, H2S is not only of volcanic origin. It is also formed from organic sources, such as decomposing organic materials, and therefore poses a considerable danger in and around old-fashioned cesspits, where concentrations may reach up to 800 ppm in the air (23.5 mM) [23]. If, how, and where such organic sources of H2S have or are currently being used for healthy bathing has so far escaped our attention.
Talking about cesspits, H2S is also formed in the gastrointestinal tract of most animals and humans by the action of the gut microbiota. In this case, concentrations of H2S may reach up to 0.3–3.4 mM in the gut, which is quite impressive considering that H2S is already biologically active in micromolar (10–30 μM) concentrations [24,25,26]. In addition, whilst most of the H2S formed during digestion may escape like the Saarland Hurricanes on a high note through the backdoor, some of it may also cross the gut-blood barrier and exert its activity inside the body [27,28].
Notably, the production of H2S in gut involves microbes belonging to the family of dissimilatory sulfate reducers, such as Desulfovibrio and Bilophila species. H2S is therefore primarily produced in the colon by reduction and not via cleavage of amino acids as in mammalian cells. The concentrations of H2S produced by the gut bacteria range from 0.3 to 3.4 mM [29]. Regardless of the mode of production, H2S subsequently finds its way into the bloodstream with serum concentrations normally ranging from 34.0 to 36.4 µM in healthy individuals [29,30].
In the gut itself, H2S may have both a harmful and a beneficial impact on local microbiota depending upon its concentration. The higher concentrations (high micromolar to millimolar) may lead to the disruption of mucus and inflammation, whilst lower, i.e., nano- and low micromolar concentrations, have been reported to actually stabilize mucus layers, protect biofilm from fragmentation and encourage adherence of the microbiota biofilm to the epithelium, prohibit the escape of invasive pathobionts, and resolve inflammation and tissue injury [31,32,33].
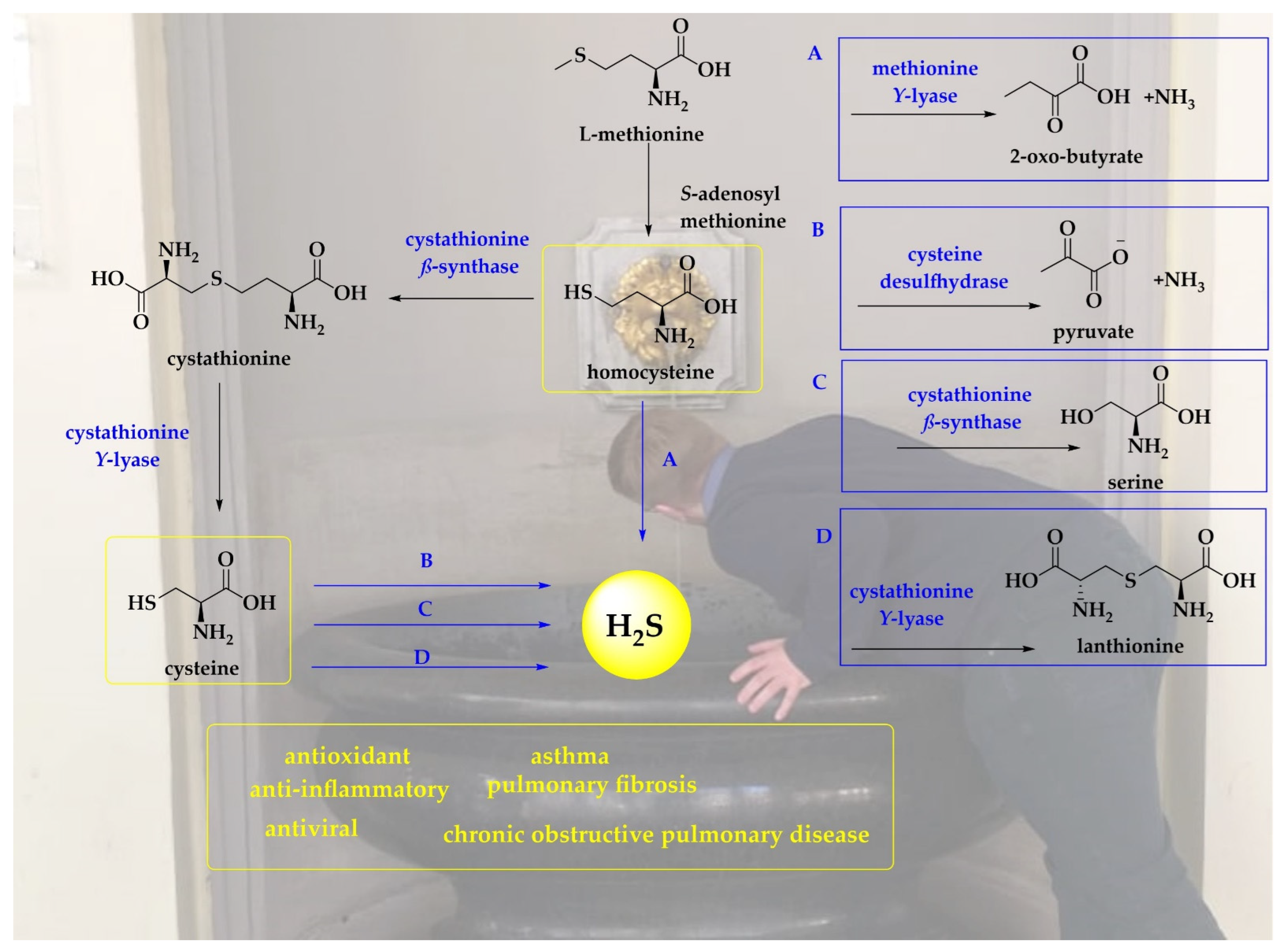
Indeed, H2S is no stranger to the human body, also in the hypothetical absence of bacteria. Human erythrocytes have been reported to generate H2S from both organic and inorganic polysulfides at constant concentrations reaching up to 170 µmol (L cells)−1 min−1 [34,35,36]. The conversion of garlic-derived polysulfides to H2S involve thiol- and glutathione-dependent reactions supported by glucose [36]. Moreover, the interactions of GSH or nicotinamide adenine dinucleotide (NADH) or nicotinamide adenine dinucleotide phosphate (NADPH) in the cell lysate also produce H2S [34,35,36]. The conversion of cysteine to cysteine per- and -polysulfides is catalyzed by cysteinyl tRNA synthetase (CARS2). Thioredoxin and GSH may reduce cysteine per- and -polysulfides employing NADPH as a source of electrons to produce H2S [37]. The non-enzymatic pathways involve the catalytic production of H2S from cysteine by iron and Vitamin B6 under physiological conditions [38]. Other major pathways for the production of H2S in animals and humans are presented in Figure 1.
H2S plays a role as a signaling molecule, similar to other gaseous transmitters, such as nitric oxide (●NO) and carbon monoxide (CO). Among these molecules, H2S is rather special as it is able to dissociate to hydrogen sulfide ions (HS−) with a dissociation constant pKa1 of 7.0, suggesting that at a physiological pH of 7.4, 28% of total hydrogen sulfide exists as H2S whilst 72% exists as HS− and very little as S2−. Indeed, the high pKa2 value of more than 12 suggests that S2− may not be formed in physiological systems [42]. H2S may therefore occur in considerable concentrations of up to 35–80 µM in the human bloodstream compared to 24.4–24.8 µM for ●NO [43,44].
Notably, animals and humans are unable to access H2S via the reductive avenue employed by bacteria in the notorious cesspits and the gut mentioned already. Therefore, sulfate (SO42−) and to a lesser extent sulfite (SO32−), which may be consumed directly or indirectly via daily nutrition, are not immediate sources of H2S, unless, of course, they are processed by gut bacteria.
As volatile H2S is highly soluble in aqueous solutions and in lipids, it is able to cross the plasma membranes and finds its way towards the lungs. Here, it may actually even cure a few respiratory diseases, such as obstructive respiratory disease, emphysema, pulmonary fibrosis, pulmonary inflammation, pancreatic inflammatory/respiratory lung injury, bronchiectasis, and bronchial asthma [45,46]. Intriguingly, serum concentrations of H2S vary significantly amongst various respiratory conditions including asthma (comparably low with 55.8 ± 13.6 μM vs. 75.2 ± 13.0 μM in healthy subjects) and chronic obstructive pulmonary disease (comparably high with 149 ± 77.6 μM vs. 90.6 ± 52.7 μM in healthy smokers and 91.0 ± 62.2 μM in healthy non-smokers), highlighting the possible application of H2S as a biomarker [47].
Furthermore, the biological activity of H2S and its physiological manifestations have recently been linked to the prevention and treatment of COVID-19. H2S may (a) block the entry of this virus into cells by interfering with the host receptors, (b) stop the viral replication by interfering with RNA-dependent RNA polymerase (RdRp), and (c) provoke the activation of a hyperinflammatory cytokine storm against the virus [48,49,50,51].
Intriguingly, H2S is also produced in the skin by both enzymatic and non-enzymatic pathways. Surprisingly, CSE, CBS, and 3-mercaptopyruvate sulfur transferase which are found in brain have also been detected in the epidermis [52]. The non-enzymatic pathways include reduction of elemental sulfur by GSH involving NADH or NADPH as electron donors or the release of H2S from iron–sulfur proteins containing Fe2S2, Fe3S4, or Fe4S4 clusters, such as ferredoxins and Rieske proteins [34,53]. Irrespective of the pathway of production, H2S plays several vital physiological roles in skin, such as promotion of cell proliferation, vasodilatation, apoptosis, and inflammation. As in the case of lung disorders, the amounts of H2S produced in various dermatological diseases such as psoriasis, melanoma, and other dermatoses vary [52,54].
2.2. Sulfur Dioxide (SO2)
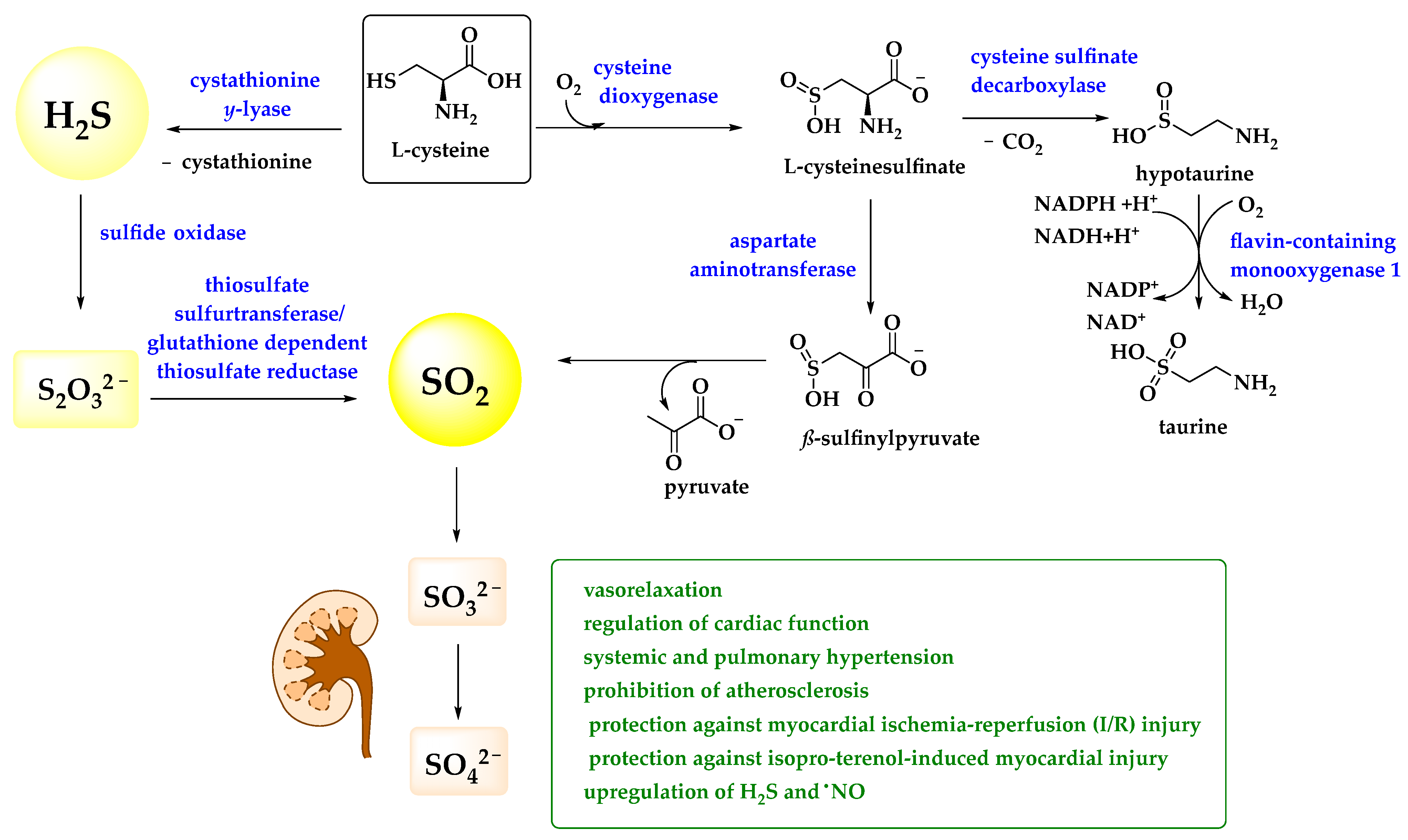
Compared to H2S, the biological roles of sulfur dioxide are less apparent and prominent. Admittedly, SO2 is a rather toxic sulfur compound, an environmental pollutant associated with acidic rain (containing sulfurous acid H2SO3) and severe damage to forests in the 1970s and 1980s [55]. It is also occasionally used to fumigate hospital wards and as a preservative for nuts and fruits and, of course, wine [56,57]. Nonetheless, Ji et al. have reported that ionic sulfite(s), derivatives of SO2, has been found in human serum at a concentration of up to 10 μM whilst Du et al. have detected an endogenous SO2 concentration of 15.54 ± 1.68 μM in rat plasma [58,59]. Under physiological conditions, SO2 is mostly found as sulfite SO32− ion, as it dissociates in water with first and second pKa values of 1.81 and 6.97, respectively, suggesting a 1:3 ratio for HSO3− and SO32− [60]. Indeed, similar to H2S, SO2—in the form of (hydrogen) sulfite—may be produced in the gastrointestinal tract by the reductive actions of bacteria, and even inside the human body by the transamination of sulfur-containing amino acids thanks to the activity of aspartate aminotransferase (AAT) (Figure 2) [61].
To date, a brief literature review on possible beneficial roles of SO2 in animals and humans only retrieves a few, yet quite interesting publications on a postulated role of the gas and its ions in cellular signaling, notably in the context of pathologies affecting the cardiovascular, nervous, and respiratory systems. In these studies, it looks as if SO2 may be involved in the normal physiological functions of the cardiovascular system, including vasorelaxation and regulation of cardiac function [62]. Moreover, SO2 may improve both systemic and pulmonary hypertension, prohibit atherosclerosis, and provide protection against myocardial ischemia-reperfusion (I/R) injury and isoproterenol-induced myocardial injury [62]. Intriguingly, SO2 is able to upregulate endogenous levels of other gasotransmitters, such as H2S or ●NO, in various cardiovascular disorders, such as systemic or pulmonary hypertension and atherosclerosis [62].
Mechanistically, endogenous SO2 manifests its biological activity in these studies through its involvement in the regulation of apoptosis. In this context, it has been reported that the improvements in the vascular remodeling in hypertension and pulmonary hypertension are achieved through increasing apoptosis of vascular smooth muscle cells (VSMCs) and prohibiting the apoptosis of vascular endothelial cells (ECs) [65]. Endogenous SO2 also provides cardio-protection by suppressing the apoptosis of cardiomyocytes [66]. Whether or not such initially beneficial actions are—at closer inspection—really desirable and may even be used in the context of therapy is a major and so far not really answered question.
Likewise, SO2 serves as a double-edged sword in the nervous system where on one hand it facilitates kainic acid induced hippocampal neuronal apoptosis leading to epileptic brain damage, whilst on the other hand, it decreases neuronal apoptosis and provides neuro-protection in recurrent febrile seizure and global brain I/R injury [67,68]. Furthermore, endogenous SO2 is closely linked with the pathogenesis of blinding retinal disease since the downregulation of endogenous SO2 has been reported to result in increased apoptosis of 661w retinal photoreceptor cells [69]. Then again, in the respiratory system, endogenous SO2 has been reported to assist the apoptosis of polymorphonuclear neutrophils (PMNs) and to inhibit the apoptosis of alveolar macrophages in order to provide protection against acute lung injury [63,70]. In any case, the relationship between SO2 and apoptosis is complicated, often dependent upon its exact concentration and the cell type exposed to it, and clearly requires further investigation.
3. Volatile Natural Products
Compared to structurally simple volatile inorganic sulfur compounds such as H2S and SO2, the field of organic sulfur compounds is considerably more diverse, and many of them are secondary metabolites serving as antioxidants or protectants against infection or predators [71]. The high reactivity often associated with such sulfur compounds then translates into powerful biological activities also outside the plants, bacteria, fungi, or animals of origin. Nonetheless, there are also quite a few limitations when it comes to volatile organic sulfur molecules. In general, these molecules need to be produced, stored, and/or distributed by the producer in a more or less controlled and stable manner, which is a challenge for any plant, bacterium, fungus, or animal. Secondly, as some of these molecules are rather active in biology, they should also not necessarily (re-)act prematurely, for instance by attacking the producer rather than the target. Furthermore, their distribution needs to be controlled, which is a clear challenge for a gas unless there are (de-)protonated ionic forms circumventing this issue. Fourthly, most of these molecules can be metabolized inside the target organism and one may therefore need to pay more attention to the more durable metabolites rather than the original volatile molecule itself.
3.1. Metabolites of H2S
H2S has the tendency to oxidize into inorganic H2Sx species with x ranging from 1–7 before further oxidation which produces elemental S8 and a precipitate [72,73]. The exact number of sulfur atoms present in H2Sx species under physiological conditions remains a matter of thorough investigation and may not only occur by spontaneous, random oxidation. There is speculation, for instance, that the production of H2Sx species could involve either the enzyme 3-mercaptopyruvate sulfurtransferase (3MST) and also interactions of H2S with ●NO. In contrast to H2S, which S-thiolates oxidized cysteine residues and reduces cysteine disulfide bonds, H2Sx can form per- and polysulfide motifs attached to cysteine residues of target proteins, thus modifying their activity [74]. Not surprisingly, biological activities of H2Sx include, among others, regulation of the activity of the tumor suppressor Phosphatase and Tensin Homolog deleted on Chromosome 10 (PTEN), facilitation of the translocation of Nuclear factor-erythroid factor 2-related factor 2 (Nrf2), and suppression of the activity of Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) [75,76,77].
In mammalian mitochondria, the conversion of H2S to the polysulfide is catalyzed by sulfide quinone oxidoreductase (SQR), and this polysulfide is subsequently transferred to sulfite to produce thiosulfate (S2O32−) or GSH persulfide (GSSH) [78,79]. Similarly, sulfide-producing enzymes, such as CSE and CBS are able to catalyze the beta-elimination reaction of cystine, to produce cysteine persulfide and polysulfide [80,81,82]. Sulfenic acids (RSOH) and S-nitrosothiols (RSNO) are also two very important players for the non-enzymatic production of persulfide. Protein thiols are subject to reversible oxidation to sulfenic acid by S-sulfenylation [83]. The pre-existing S-sulfenylation has been proposed to be responsible for the persulfidation of GAPDH and bovine serum albumin (BSA) [84,85]. S-nitrosothiol represents another form of oxidized thiol, which reacts with H2S to produce persulfides [86]. In contrast, sulfhydral radicals (●SH) are produced as a result of oxidation of H2S by metal centers, such as copper and iron, and may then react with free thiols and H2S to produce protein persulfides and polysulfides, respectively [87].
Besides oxidized metabolites of H2S, we should therefore turn our immediate attention to the metabolites of H2S, which is frequently methylated in animals and humans and therefore comes in concert with molecules such as methyl sulfide (MS, CH3SH), dimethyl sulfide (DMS, CH3SCH3), and the trimethyl sulfonium ion ((CH3)3S+) [27,88]. The trimethyl sulfonium ion is water-soluble and excreted via the kidneys. MS and DMS are volatile with boiling points of 6 °C and 37 °C, respectively. They circulate the human body and are released via the lung and skin. Concentrations of up to 178 ppb (3.78 µM) for MS and 35 ppb (0.56 µM) for DMS have been observed in the breath and therefore may induce some biological activities in the cells lining the airways [89].
Apart from that, the literature is vague on the biological impact of MS and DMS, especially at lower (micromolar) concentrations and prolonged exposure. Among the few studies addressing the biological activities of such metabolites, there are reports that the presence of VSCs, including the methylated metabolites of hydrogen sulfide, may be associated with chronic periodontitis and other oral conditions [90,91]. Then again, DMS has also been reported to provide an effective shield against oxidative stress and even to prolong the lifespan of mammals through a catalytic mechanism involving methionine sulfoxide reductase A [92].
3.2. Simple Alkyl Sulfides
Compounds such as dimethyl sulfide (CH3SCH3) and diethyl sulfide (C2H5SC2H5) may indeed not be the most active sulfides in biology, as the dialkyl monosulfide motif tends to be rather unreactive under physiological conditions. It is also present, for instance, in the proteinogenic amino acid methionine, and although its sulfur can be oxidized to a sulfoxide and theoretically also to a sulfone, this is a rare event and various methionine S-oxide reductases are at hand to reverse this modification [93].
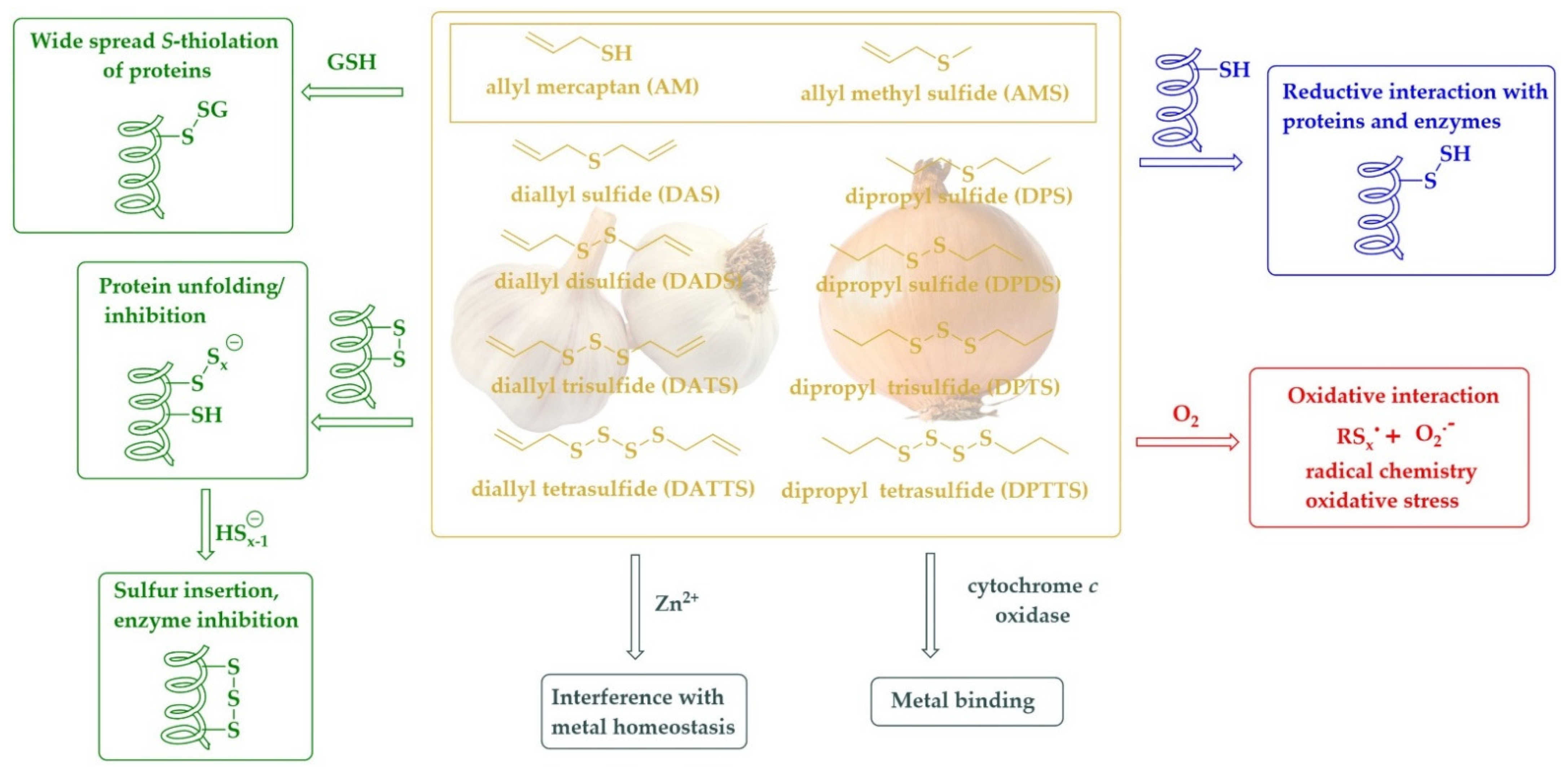
Moving from the mono- to the di-, tri-, or tetrasulfide changes this reactivity considerably [94]. Unlike the carbon-sulfur bond, the sulfur-sulfur bond is highly reactive. Besides oxidation of the sulfur to an S-oxide, in this case a thiosulfinate (RS(O)SR’) or thiosulfonate (RS(O)2SR’), it also allows reduction, for instance via an eloquent thiol/disulfide exchange mechanism [95,96]. Not surprisingly, the dialklyl disulfides and dialkyl polysulfides (RSxR’, x ≥ 2) also exhibit a range of pronounced biological activities, for instance as antioxidants and antimicrobial agents [97,98]. Among them, the dipropyl sulfides occur naturally in onions and show a palette of activities, mostly cytotoxic and antimicrobial, and mostly due to their ability to oxidatively modify certain target proteins and enzymes [99]. As a rule of thumb, the longer the sulfur-sulfur chain, the higher the reactivity and also the biological activity, with the caveat that longer sulfur-sulfur chains are also becoming more unstable. Most studies therefore consider the tri- and tetrasulfides as the most promising compounds for biological applications.
To underline the importance of this class of simple yet extraordinarily active compounds, we shall briefly mention dimethyl disulfide (DMDS, CH3SSCH3, melting point −85 °C, boiling point 110 °C) and dipropyl trisulfide (DPTS, C3H7SSSC3H7, boiling point 72°C) as prominent and perhaps also representative substances in this field. The naturally occurring DMDS shows antifungal activity against Sclerotinia minor by attacking the membrane of this fungus and also induces systemic resistance against white mold in host plants [100]. This simple disulfide also inhibits the two plant pathogens Agrobacterium tumefaciens and Agrobacterium vitis by functioning as a bacteriostatic agent [101,102]. Although DMDS has a boiling point of 110 °C, it has been applied air bound and, perhaps not surprisingly, is in use already as an innovative fumigant under the trade name PALADIN® to target soil-borne plant pathogens [103]. There are also studies indicating that DMDS may interfere with bacterial quorum sensing communication by significantly suppressing the transcription of N-acyl homoserine lactone synthase genes [102,104]. In any case, this and other simple volatile disulfides possess considerable potential for practical applications in the—literally—field of antimicrobial fumigation.
Dipropyl tetrasulfide (DPTTS, C3H7SSSSC3H7, boiling point 127 °C), on the other hand, is less volatile and has been studied extensively as a redox modulator in many disease models, among them a mouse model indicative of systemic sclerosis (scleroderma) [99]. In this model, DPTTS derived synthetically and also isolated from onions shows considerable beneficial properties by selectively killing the diseased fibroblasts and, thereby, counteracting this disease. When exposed to DPTTS, the concentration of H2O2 significantly increases in HOCl-fibroblasts as compared to normal fibroblasts. Moreover, DPTTS provides higher cytotoxic and pro-apoptotic activities in HOCl fibroblasts than in normal fibroblasts [99]. Once again, the mechanisms underlying this activity may be complex, and in any case related to the pronounced redox activity associated with the sulfur-sulfur bond and in need of further investigation.
The discussion of DPTTS from onions almost immediately also brings us to its relative in garlic, diallyl trisulfide (C3H5SSSC3H5, DATS). Indeed, DATS is equally, if not more active biologically compared to DPTTS and is also the source of centuries of medicinal inspiration and a plethora of publications [105,106,107]. Similar to DPTTS, DATS is volatile with a boiling point of 120 °C, and there are numerous anecdotes of DATS and its relatives diallyl disulfide (C3H5SSC3H5, DADS, boiling point 187.5 °C) and diallyl tetrasulfide (C3H5SSSSC3H5, DATTS, boiling point 45 °C) fuming out entire incubators when applied in Petri dish-based studies [11]. The biological activities associated with these diallyl sulfides range from antioxidant and protective to redox modulation and cytotoxic and are usually associated with the reactivity of the sulfur-sulfur bond acting in cahoots with the double bond of the allyl group. A brief overview of such compounds and their activities have been depicted in Figure 3. As mentioned already, the monosulfides, namely dipropyl sulfide (C3H7SC3H7, DPS, boiling point 142.9 °C) and diallyl sulfide (C3H5SC3H5, DAS, boiling point 138.6 °C) are also found in onions and garlic, respectively, although the biological activities associated with these less reactive compounds is also more restricted. DAS, for instance, has been reported to serve as a selective inhibitor of cytochrome P450 2E1 (CYP2E1), and therefore it is not only able to inhibit the cellular toxicities associated with alcohol and xenobiotic drugs; it also limits the cellular toxicities associated with HIV proteins and diabetes [108].
As for practical applications, the polysulfides from onions and garlic are employed already in the field of agriculture, in essence in the form of sprays and pellets [109]. To the best of our information, their possible use in fumigation has not been explored further so far. Notably, minute amounts of diallyl sulfides are present in so-called garlic breath after consumption of raw garlic [110]. The concentrations of DAS, DADS, and DATS increase from around 6 ppb (52.5 nM), 10 ppb (80.5 nM), and 3 ppb (16.8 nM) to 13 ppb (113.8 nM), 130 ppb (1.1 µM), and 5.5 ppb (30.9 nM), respectively, in the breath exhaled after ingestion of raw garlic [111]. Intriguingly, the concentration of DADS reaches its peak of 130 ppb in around two hours after ingestion of garlic followed by its rapid removal. The concentration of dimethylsulfide (DMS), in contrast, increases from 30 ppb (482.9 nM) to 60 ppb (965.7 nM) in 8 h and reaches the peak of 90 ppb (1.5 µM) in around 24 h after ingestion of garlic, possibly formed via the H2S pathway discussed already [111]. If and to which extent such low and sub-micromolar concentrations of DAS, DADS, and DATS in breath may exert any notable biological activity on cells lining the respiratory system is so far unclear, although not entirely impossible.
Indeed, such sulfur compounds are also excreted via the skin in the form of “human skin gas” and may therefore not only affect the respiratory system, but also the skin cells they encounter during their release. Sato et al. have recently reported that DADS presents a mean emission flux of 0.18 ng (1.23 pmol) cm−2 h−1 (n = 30) from the surface of human skin without ingestion of garlic. The consumption of 45 g of grilled garlic results in increased emission flux of DADS achieving a maximum of 4.3–5.2 ng (29.39–35.55 pmol) cm−2 h−1. Even after 8 h, the values of emission fluxes are higher than the base-line values [112]. As for breath, whether such small, albeit sustained, concentrations of these substances exert any major biological effects in and/or near the skin requires further investigations. It may, for instance, be feasible that some of these substances accumulate or enrich in the lung and skin cells and subsequently provide an antioxidant shield or affect microbes attached to these organs. In any case, the topic of exhalation of intact or metabolized sulfur compounds is rather stimulating as it brings the (active) sulfur compounds into intimate contact with tissues where they may have numerous benefits.
3.3. Metabolites
The discussion of exhaled rather than inhaled volatile and biologically active molecules essentially brings us to the various metabolites of natural sulfur compounds. In this context, it is essential to note that the metabolites, and not necessarily the original sulfur compounds, may themselves be volatile and/or active, as is the case, for instance, for several ingredients found in the Allium plants, such as alliin (S-allyl-L-cysteine) [8]. Equally notably, such metabolites are often less reactive than the original compounds as they have already lost some of their (re-)activity inside the body. Thus, the exhaled metabolites tend to have a different and generally lower biological activity when compared to the original substance taken up orally [113].
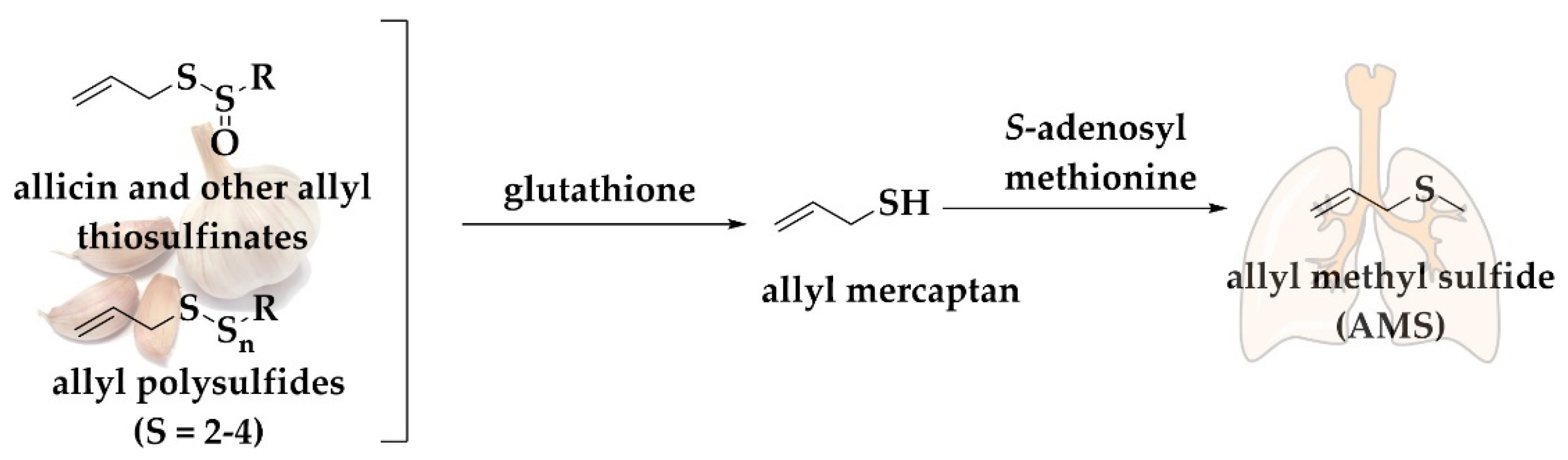
In the case of the infamous “garlic breath”, the simple alkyl sulfides discussed in Section 3.2, e.g., DMS, DAS, DADS, and DATS are still present, albeit in rather low concentrations [111]. This is partially due to release of some of these substances from the mouth and stomach, in essence before they have been taken up and metabolized. Excretion through the skin and lung differs, as mentioned already, and here one finds a different spectrum of—metabolized—volatile sulfur compounds, namely allyl methyl sulfide (C3H5SCH3, AMS, boiling point 92 °C).
Allicin is converted under physiological conditions to polysulfides and then to this volatile monosulfide, AMS, which is present as a major constituent of garlic breath (Figure 4). Studies have shown that after the consumption of raw garlic, the content of AMS in breath is linearly proportional to the amount of allicin consumed [114]. AMS represents the main metabolite of allicin exerted via the breath. Generally, 90% of the allicin consumed is turned into AMS, whilst allyl mercaptan (C3H5SH) is only a temporary intermediate in the formation of AMS. Allicin-derived DADS and DATS are also metabolized mainly to AMS. Furthermore, AMS is an active metabolite, responsible for the ability of allicin to increase the acetone levels in breath. Intriguingly, alveolar breath has been shown to contain 18% higher AMS concentrations compared to whole breath [115]. The release of AMS is not only confined to lungs since it has been recently reported that AMS presents a mean emission flux of 0.22 ng (2.50 pmol) cm−2 h−1 (n = 30) from the surface of human skin without ingestion of garlic. The consumption of 45 g of grilled garlic results in increased emission flux of AMS achieving a maximum of 2.8–3.0 ng (31.76–34.03 pmol) cm−2 h−1 [112].
AMS has been associated with several potential biological applications, especially in the treatment of lung cancer [116]. AMS, alongside DADS and DAS, has been reported to reduce the formation of cancerous lung nodules by 54.8% in the C57BL/6 mice model [116]. Furthermore, a recent study shows that AMS may act as an efficient antiviral agent. Based on a computational study, AMS may attach to the phospholipid surface of a virus, such as the SARS-CoV-2 virus, and thereby denature the virus [117]. Another study suggests that AMS might also be useful as a fumigation agent. Here, AMS shows a significant toxicity against Actinobacillus pleuropneumoniae, which causes Pleuropneumonia, a serious lung disease in pigs [118]. AMS may also be beneficial in the treatment of Alzheimer Disease (AD). In this case, 500 µM AMS inhibit β-secretase, acetyl choline esterase, and butyrylcholinesterase by 18.4%, 24.5%, and 21.0%, respectively [119].
3.4. The Durian Fruit—A Treasure Chest of Public Annoyance and Volatile Sulfur Compounds
Onions and garlic are not the sole producers of biologically active sulfur compounds. Broccoli has been mentioned already and mustard must also be on the rollcall, as these vegetables contain biologically active sulforaphane and allyl isothiocyanate, respectively (Figure 5a). Indeed, the less volatile sulforaphane (boiling point 368.2 °C) is often associated with antioxidant and preventive activities, whereas the more volatile mustard oil (boiling point 170 °C) shows pronounced anticancer, antioxidant, and antimicrobial activities in various in vitro and cell culture studies [120,121].
Compared to these popular culinary plants present in almost every cuisine across the globe, the durian fruit cultivated in Southeast Asia is more for lovers of extreme tastes [122]. Its pungent smell is often a matter of considerable annoyance and its transport and public consumption is frequently forbidden [123].
The durian fruit is rich in sugars, fat, fiber, and micronutrients, such as potassium. It is also home to a range of rather smelly volatile compounds, such as esters, sulfur, thioacetals, thioesters, thiolanes, and alcohols, some of them pictured in Figure 5b [124]. Thanks to the presence of these substances, the durian fruit is cherished in some quarters for its supposed anti-hyperglycemic, anti-atherosclerotic, probiotic and even anti-proliferative activities and its individual sulfur-containing ingredients lend themselves for further investigations [125].
4. Conclusions
Our brief encounter with volatile, biologically active sulfur agents has shown that quite a palette of such substances is present as natural products in plants, fungi, animals, and microbes. Many of these compounds serve as antioxidants or as part of host defense and therefore may also be of interest for potential uses in the fields of agriculture and medicine. Among the inorganic substances, H2S clearly stands out as an exogenous toxic gas at higher concentrations and an endogenous signaling molecule in humans and animals at lower concentrations. If and how its oxidized companion SO2 may also serve as a signaling molecule is still a matter of research and debate. More eloquent sulfur-containing agents are found in plants such as garlic and onions, and besides possible uses as culinary delights may also produce metabolites during (in-)digestion, which themselves may be active biologically, inside the body and during excretion. As usual, reactivity and activity in vitro alone are not sufficient, and concentrations—and periods of exposure—also matter if one tries to turn such sulfur compounds into good antioxidants or antimicrobial agents. The field of volatile sulfur compounds, especially the ones of natural origin, is therefore one worth considering and cultivating, as such compounds may have rather valuable applications in agriculture, medicine, and also as a culinary highlight in our daily nutrition. So, let’s go research, let’s go.
Author Contributions
Conceptualization, C.J. and M.J.N.; methodology, E.T., M.A.L., Y.N. and A.Y.A.; investigation, E.T., M.A.L., Y.N. and A.Y.A.; data curation, M.J.N.; writing—original draft preparation, C.J. and M.J.N.; writing—review and editing, E.T., M.A.L., Y.N. and A.Y.A.; supervision, C.J. and M.J.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors would like to express their explicit gratitude to all the members of the EU COST Action 16112 “NutRedOx” for the many stimulating discussions on nutrition and health. The authors would also like to thank the University of Saarland, the Landesforschungsförderungsprogramm of the State of Saarland (Grant No: WT/2—LFFP 16/01), and the INTERREG VA GR program (BIOVAL, Grant No: 4-09-21). The authors express special thanks to Ken Rory and many other colleagues of the Academiacs International (www.academiacs.eu) and Pharmasophy networks for helpful discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Milito, A.; Brancaccio, M.; D’Argenio, G.; Castellano, I. Natural Sulfur-Containing Compounds: An Alternative Therapeutic Strategy against Liver Fibrosis. Cells 2019, 8, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miękus, N.; Marszałek, K.; Podlacha, M.; Iqbal, A.; Puchalski, C.; Świergiel, A.H. Health Benefits of Plant-Derived Sulfur Compounds, Glucosinolates, and Organosulfur Compounds. Molecules 2020, 25, 3804. [Google Scholar] [CrossRef] [PubMed]
- Kalaras, M.D.; Richie, J.P.; Calcagnotto, A.; Beelman, R.B. Mushrooms: A Rich Source of the Antioxidants Ergothioneine and Glutathione. Food. Chem. 2017, 233, 429–433. [Google Scholar] [CrossRef]
- Ruhee, R.T.; Roberts, L.A.; Ma, S.; Suzuki, K. Organosulfur Compounds: A Review of Their Anti-Inflammatory Effects in Human Health. Front. Nutr. 2020, 7, 64. [Google Scholar] [CrossRef] [PubMed]
- Doleman, J.F.; Grisar, K.; Van Liedekerke, L.; Saha, S.; Roe, M.; Tapp, H.S.; Mithen, R.F. The Contribution of Alliaceous and Cruciferous Vegetables to Dietary Sulphur Intake. Food. Chem. 2017, 234, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-L.; Lin, S.-Y.; Du, H.-T.; Qin, L.; Lei, L.-M.; Chen, D. An Insight by Molecular Sensory Science Approaches to Contributions and Variations of the Key Odorants in Shiitake Mushrooms. Foods 2021, 10, 622. [Google Scholar] [CrossRef]
- Cao, X.; Cao, L.; Zhang, W.; Lu, R.; Bian, J.-S.; Nie, X. Therapeutic Potential of Sulfur-Containing Natural Products in Inflammatory Diseases. Pharmacol. Ther. 2020, 216, 107687. [Google Scholar] [CrossRef]
- Borlinghaus, J.; Foerster (née Reiter), J.; Kappler, U.; Antelmann, H.; Noll, U.; Gruhlke, M.C.H.; Slusarenko, A.J. Allicin, the Odor of Freshly Crushed Garlic: A Review of Recent Progress in Understanding Allicin’s Effects on Cells. Molecules 2021, 26, 1505. [Google Scholar] [CrossRef]
- Gruhlke, M.C.H.; Nicco, C.; Batteux, F.; Slusarenko, A.J. The Effects of Allicin, a Reactive Sulfur Species from Garlic, on a Selection of Mammalian Cell Lines. Antioxidants 2016, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Gruhlke, M.C.H.; Antelmann, H.; Bernhardt, J.; Kloubert, V.; Rink, L.; Slusarenko, A.J. The Human Allicin-Proteome: S-Thioallylation of Proteins by the Garlic Defence Substance Allicin and Its Biological Effects. Free Radic. Biol. Med. 2019, 131, 144–153. [Google Scholar] [CrossRef]
- Leontiev, R.; Hohaus, N.; Jacob, C.; Gruhlke, M.C.H.; Slusarenko, A.J. A Comparison of the Antibacterial and Antifungal Activities of Thiosulfinate Analogues of Allicin. Sci. Rep. 2018, 8, 6763. [Google Scholar] [CrossRef] [PubMed]
- Bhatwalkar, S.B.; Shukla, P.; Srivastava, R.K.; Mondal, R.; Anupam, R. Validation of Environmental Disinfection Efficiency of Traditional Ayurvedic Fumigation Practices. J. Ayurveda Integr. Med. 2019, 10, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.; Gould, E.; Tinson, R.; Groom, M.; Hamilton, C.J. Think Yellow and Keep Green—Role of Sulfanes from Garlic in Agriculture. Antioxidants 2016, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Hoffhines, A.J.; Moore, K.L.; Leary, J.A. Determination of the Sites of Tyrosine O-Sulfation in Peptides and Proteins. Nat. Methods 2007, 4, 583–588. [Google Scholar] [CrossRef]
- Nagy, P.; Beal, J.L.; Ashby, M.T. Thiocyanate Is an Efficient Endogenous Scavenger of the Phagocytic Killing Agent Hypobromous Acid. Chem. Res. Toxicol. 2006, 19, 587–593. [Google Scholar] [CrossRef]
- Bonifay, V.; Barrett, T.J.; Pattison, D.I.; Davies, M.J.; Hawkins, C.L.; Ashby, M.T. Tryptophan Oxidation in Proteins Exposed to Thiocyanate-Derived Oxidants. Arch. Biochem. Biophys. 2014, 564, 1–11. [Google Scholar] [CrossRef]
- Raciti, L.; Calabrò, R.S. Can Volcanic Trace Elements Facilitate Covid-19 Diffusion? A Hypothesis Stemming from the Mount Etna Area, Sicily. Med. Hypotheses 2020, 144, 110058. [Google Scholar] [CrossRef]
- Karagülle, M.Z.; Karagülle, M. Effects of Drinking Natural Hydrogen Sulfide (H2S) Waters: A Systematic Review of in Vivo Animal Studies. Int. J. Biometeorol. 2020, 64, 1011–1022. [Google Scholar] [CrossRef]
- Doujaiji, B.; Al-Tawfiq, J.A. Hydrogen Sulfide Exposure in an Adult Male. Ann. Saudi Med. 2010, 30, 76–80. [Google Scholar] [CrossRef]
- Bad Nenndorfer Schwefelquelle-Eine Der Stärksten in Europa Bad Nenndorf. Available online: https://www.badnenndorf.de/kur-and-gesundheit/unsere-heilmittel-moor-schwefel-sole/schwefel/ (accessed on 16 March 2022).
- Sokrateva, T.D.; Roussev, B.H.; Nashar, M.A.; Kiselova-Kaneva, Y.D.; Mihaylova, G.M.; Todorova, M.N.; Pasheva, M.G.; Tasinov, O.B.; Nazifova-Tasinova, N.F.; Vankova, D.G.; et al. Effects of Sulphur-Containing Mineral Water Intake on Oxidative Status and Markers for Inflammation in Healthy Subjects. Arch. Physiol. Biochem. 2021, 127, 327–336. [Google Scholar] [CrossRef]
- Physicochemical Analysis of Varna Basin Mineral Water-ProQuest. Available online: https://0-www-proquest-com.brum.beds.ac.uk/openview/4b6fab94d5ade3ac7a79409f75fcde9d/1?pq-origsite=gscholar&cbl=1536338 (accessed on 16 March 2022).
- Li, X.; O’Moore, L.; Song, Y.; Bond, P.L.; Yuan, Z.; Wilkie, S.; Hanzic, L.; Jiang, G. The Rapid Chemically Induced Corrosion of Concrete Sewers at High H2S Concentration. Water Res. 2019, 162, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Braccia, D.J.; Jiang, X.; Pop, M.; Hall, A.B. The Capacity to Produce Hydrogen Sulfide (H2S) via Cysteine Degradation Is Ubiquitous in the Human Gut Microbiome. Front. Microbiol. 2021, 12, 705583. [Google Scholar] [CrossRef] [PubMed]
- Dordević, D.; Jančíková, S.; Vítězová, M.; Kushkevych, I. Hydrogen Sulfide Toxicity in the Gut Environment: Meta-Analysis of Sulfate-Reducing and Lactic Acid Bacteria in Inflammatory Processes. J. Adv. Res. 2021, 27, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Kolluru, G.K.; Shen, X.; Bir, S.C.; Kevil, C.G. Hydrogen Sulfide Chemical Biology: Pathophysiological Roles and Detection. Nitric Oxide 2013, 35, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Ding, L.; Xie, Z.; Yang, Y.; Whiteman, M.; Moore, P.K.; Bian, J.-S. A Review of Hydrogen Sulfide Synthesis, Metabolism, and Measurement: Is Modulation of Hydrogen Sulfide a Novel Therapeutic for Cancer? Antioxid. Redox Sign. 2019, 31, 1–38. [Google Scholar] [CrossRef]
- Tomasova, L.; Konopelski, P.; Ufnal, M. Gut Bacteria and Hydrogen Sulfide: The New Old Players in Circulatory System Homeostasis. Molecules 2016, 21, 1558. [Google Scholar] [CrossRef] [PubMed]
- Suarez, F.; Furne, J.; Springfield, J.; Levitt, M. Insights into Human Colonic Physiology Obtained from the Study of Flatus Composition. Am. J. Physiol. Gastroint Liver Physiol. 1997, 272, G1028–G1033. [Google Scholar] [CrossRef]
- Furne, J.; Saeed, A.; Levitt, M.D. Whole Tissue Hydrogen Sulfide Concentrations Are Orders of Magnitude Lower than Presently Accepted Values. Am. J. Physiol. Reg. Integ. Comp. Physiol. 2008, 295, R1479–R1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buret, A.G.; Allain, T.; Motta, J.-P.; Wallace, J.L. Effects of Hydrogen Sulfide on the Microbiome: From Toxicity to Therapy. Antioxid. Redox Sign. 2021, 36, 211–219. [Google Scholar] [CrossRef]
- Wallace, J.L.; Wang, R. Hydrogen Sulfide-Based Therapeutics: Exploiting a Unique but Ubiquitous Gasotransmitter. Nat. Rev. Drug Discov. 2015, 14, 329–345. [Google Scholar] [CrossRef]
- Blachier, F.; Beaumont, M.; Kim, E. Cysteine-Derived Hydrogen Sulfide and Gut Health: A Matter of Endogenous or Bacterial Origin. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Searcy, D.G.; Lee, S.H. Sulfur Reduction by Human Erythrocytes. J. Exp. Zool. 1998, 282, 310–322, Wiley Online Library. Available online: https://0-onlinelibrary-wiley-com.brum.beds.ac.uk/doi/10.1002/(SICI)1097-010X(19981015)282:3%3C310::AID-JEZ4%3E3.0.CO;2-P (accessed on 21 March 2022). [CrossRef]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen Sulfide Mediates the Vasoactivity of Garlic. Proc. Nat. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- di Masi, A.; Ascenzi, P. H2S: A “Double Face” Molecule in Health and Disease. BioFactors 2013, 39, 186–196. [Google Scholar] [CrossRef]
- Dóka, É.; Pader, I.; Bíró, A.; Johansson, K.; Cheng, Q.; Ballagó, K.; Prigge, J.R.; Pastor-Flores, D.; Dick, T.P.; Schmidt, E.E.; et al. A Novel Persulfide Detection Method Reveals Protein Persulfide- and Polysulfide-Reducing Functions of Thioredoxin and Glutathione Systems. Sci. Adv. 2016, 2, e1500968. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Minkler, P.; Grove, D.; Wang, R.; Willard, B.; Dweik, R.; Hine, C. Non-Enzymatic Hydrogen Sulfide Production from Cysteine in Blood Is Catalyzed by Iron and Vitamin B6. Commun. Biol. 2019, 2, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Banerjee, R. PLP-Dependent H2S Biogenesis. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2011, 1814, 1518–1527. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Liu, H.; Shi, X.; Prokosch, V. Hydrogen Sulfide: Novel Endogenous and Exogenous Modulator of Oxidative Stress in Retinal Degeneration Diseases. Molecules 2021, 26, 2411. [Google Scholar] [CrossRef]
- Instagram, E.F.-W.-P. 52062 A.D.F. Elisenbrunnen. Available online: https://www.aachen-tourismus.de/en/discover/sights/details/elisenbrunnen/ (accessed on 4 April 2022).
- Li, Q.; Lancaster, J.R. Chemical Foundations of Hydrogen Sulfide Biology. Nitric Oxide 2013, 35, 21–24. [Google Scholar] [CrossRef] [Green Version]
- Hyšpler, R.; Tichá, A.; Indrová, M.; Zadák, Z.; Hyšplerová, L.; Gasparič, J.; Churáček, J. A Simple, Optimized Method for the Determination of Sulphide in Whole Blood by GC–MS as a Marker of Bowel Fermentation Processes. J. Chromatog. B 2002, 770, 255–259. [Google Scholar] [CrossRef]
- Ghasemi, A.; Zahedi Asl, S.; Mehrabi, Y.; Saadat, N.; Azizi, F. Serum Nitric Oxide Metabolite Levels in a General Healthy Population: Relation to Sex and Age. Life Sci. 2008, 83, 326–331. [Google Scholar] [CrossRef]
- Wang, R. Physiological Implications of Hydrogen Sulfide: A Whiff Exploration That Blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khattak, S.; Zhang, Q.-Q.; Sarfraz, M.; Muhammad, P.; Ngowi, E.E.; Khan, N.H.; Rauf, S.; Wang, Y.-Z.; Qi, H.-W.; Wang, D.; et al. The Role of Hydrogen Sulfide in Respiratory Diseases. Biomolecules 2021, 11, 682. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Saito, J.; Munakata, M.; Shibata, Y. Hydrogen Sulfide as a Novel Biomarker of Asthma and Chronic Obstructive Pulmonary Disease. Allergol. Int. 2021, 70, 181–189. [Google Scholar] [CrossRef]
- Bourgonje, A.R.; Offringa, A.K.; van Eijk, L.E.; Abdulle, A.E.; Hillebrands, J.-L.; van der Voort, P.H.J.; van Goor, H.; van Hezik, E.J. N-Acetylcysteine and Hydrogen Sulfide in Coronavirus Disease 2019. Antioxid. Redox Signal. 2021, 35, 1207–1225. [Google Scholar] [CrossRef]
- Citi, V.; Martelli, A.; Brancaleone, V.; Brogi, S.; Gojon, G.; Montanaro, R.; Morales, G.; Testai, L.; Calderone, V. Anti-Inflammatory and Antiviral Roles of Hydrogen Sulfide: Rationale for Considering H2S Donors in COVID-19 Therapy. Br. J. Pharmacol. 2020, 177, 4931–4941. [Google Scholar] [CrossRef]
- Yang, G. H2S as a Potential Defense against COVID-19? Am. J. Physiol. Cell Physiol. 2020, 319, C244–C249. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Teng, X.; Jin, S.; Wu, Y. The Antiviral Roles of Hydrogen Sulfide by Blocking the Interaction between SARS-CoV-2 and Its Potential Cell Surface Receptors. Oxid. Med. Cell Longev. 2021, 2021, e7866992. [Google Scholar] [CrossRef]
- Xiao, Q.; Xiong, L.; Tang, J.; Li, L.; Li, L. Hydrogen Sulfide in Skin Diseases: A Novel Mediator and Therapeutic Target. Oxid. Med. Cell Longev. 2021, 2021, e6652086. [Google Scholar] [CrossRef]
- Beinert, H.; Holm, R.H.; Münck, E. Iron-Sulfur Clusters: Nature’s Modular, Multipurpose Structures. Science 1997, 277, 653–659. [Google Scholar] [CrossRef]
- Coavoy-Sánchez, S.A.; Costa, S.K.P.; Muscará, M.N. Hydrogen Sulfide and Dermatological Diseases. Br. J. Pharmacol. 2020, 177, 857–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grennfelt, P.; Engleryd, A.; Forsius, M.; Hov, Ø.; Rodhe, H.; Cowling, E. Acid Rain and Air Pollution: 50 Years of Progress in Environmental Science and Policy. Ambio 2020, 49, 849–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, S.-M.; Xu, J.; Bai, Y.-J.; Ding, Y.; Kong, M.; Liu, H.-H.; Li, X.-Y.; Zhang, Q.-S.; Chen, H.-B.; Liu, L.-F.; et al. Sulfur Dioxide Residue in Sulfur-Fumigated Edible Herbs: The Fewer, the Safer? Food Chem. 2016, 192, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Arapitsas, P.; Guella, G.; Mattivi, F. The Impact of SO2 on Wine Flavanols and Indoles in Relation to Wine Style and Age. Sci. Rep. 2018, 8, 858. [Google Scholar] [CrossRef] [Green Version]
- Du, S.; Jin, H.; Bu, D.; Zhao, X.; Geng, B.; Tang, C.; Du, J. Endogenously Generated Sulfur Dioxide and Its Vasorelaxant Effect in Rats. Acta Pharmacol. Sin. 2008, 29, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Ji, A.J.; Savon, S.R.; Jacobsen, D.W. Determination of Total Serum Sulfite by HPLC with Fluorescence Detection. Clin. Chem. 1995, 41, 897–903. [Google Scholar] [CrossRef]
- Wang, W.; Wang, B. SO2 Donors and Prodrugs, and Their Possible Applications: A Review. Front. Chem. 2018, 6, 559. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Jin, H.; Song, Y.; Chen, S.Y.; Wang, Y.; Sun, Y.; Tang, C.; Du, J.; Huang, Y. Endogenous Sulfur Dioxide Is a Novel Inhibitor of Hypoxia-Induced Mast Cell Degranulation. J. Adv. Res. 2021, 29, 55–65. [Google Scholar] [CrossRef]
- Huang, Y.; Tang, C.; Du, J.; Jin, H. Endogenous Sulfur Dioxide: A New Member of Gasotransmitter Family in the Cardiovascular System. Oxid. Med. Cell Longev. 2016, 2016, 8961951. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Feng, Y.; Ye, X.; Peng, H.; Du, J.; Yao, X.; Huang, Y.; Jin, H.; Du, J. Endogenous SO2 Controls Cell Apoptosis: The State-of-the-Art. Front. Cell Dev. Biol. 2021, 9, 729728. [Google Scholar] [CrossRef]
- Veeravalli, S.; Phillips, I.R.; Freire, R.T.; Varshavi, D.; Everett, J.R.; Shephard, E.A. Flavin-Containing Monooxygenase 1 Catalyzes the Production of Taurine from Hypotaurine. Drug Metab. Dispos. 2020, 48, 378–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Huang, Y.; Bu, D.; Liu, A.D.; Holmberg, L.; Jia, Y.; Tang, C.; Du, J.; Jin, H. Sulfur Dioxide Inhibits Vascular Smooth Muscle Cell Proliferation via Suppressing the Erk/MAP Kinase Pathway Mediated by CAMP/PKA Signaling. Cell Death Dis. 2014, 5, e1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Xu, Z.; Xia, J.; Qin, G.; Sang, N. Sulfur Dioxide Induces Apoptosis via Reactive Oxygen Species Generation in Rat Cardiomyocytes. Environ. Sci. Pollut. Res. Int. 2019, 26, 8758–8767. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Han, Y.; Li, Q.; Zhang, J. Endogenous Sulfur Dioxide Regulates Hippocampal Neuron Apoptosis in Developing Epileptic Rats and Is Associated with the PERK Signaling Pathway. Neurosci. Lett. 2018, 665, 22–28. [Google Scholar] [CrossRef]
- Han, Y.; Yi, W.; Qin, J.; Zhao, Y.; Zhang, J.; Chang, X. Dose-Dependent Effect of Sulfur Dioxide on Brain Damage Induced by Recurrent Febrile Seizures in Rats. Neurosci. Lett. 2014, 563, 149–154. [Google Scholar] [CrossRef]
- Wenzel, A.; Grimm, C.; Samardzija, M.; Remé, C.E. Molecular Mechanisms of Light-Induced Photoreceptor Apoptosis and Neuroprotection for Retinal Degeneration. Progr. Retin. Eye Res. 2005, 24, 275–306. [Google Scholar] [CrossRef]
- Ma, H.; Huang, X.; Liu, Y.; Fan, Y. Sulfur Dioxide Attenuates LPS-Induced Acute Lung Injury via Enhancing Polymorphonuclear Neutrophil Apoptosis. Acta Pharmacol. Sin. 2012, 33, 983–990. [Google Scholar] [CrossRef] [Green Version]
- Giles, G.I.; Nasim, M.J.; Ali, W.; Jacob, C. The Reactive Sulfur Species Concept: 15 Years On. Antioxidants 2017, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Toohey, J.I. Sulfur Signaling: Is the Agent Sulfide or Sulfane? Anal. Biochem. 2011, 413, 1–7. [Google Scholar] [CrossRef]
- Nagy, P.; Winterbourn, C.C. Rapid Reaction of Hydrogen Sulfide with the Neutrophil Oxidant Hypochlorous Acid to Generate Polysulfides. Available online: https://0-pubs-acs-org.brum.beds.ac.uk/doi/full/10.1021/tx100266a (accessed on 9 May 2022).
- Kimura, H. Hydrogen Polysulfide (H2Sn) Signaling along with Hydrogen Sulfide (H2S) and Nitric Oxide (NO). J. Neural. Transm. 2016, 123, 1235–1245. [Google Scholar] [CrossRef]
- Greiner, R.; Pálinkás, Z.; Bäsell, K.; Becher, D.; Antelmann, H.; Nagy, P.; Dick, T.P. Polysulfides Link H2S to Protein Thiol Oxidation. Antioxid. Redox Signal. 2013, 19, 1749–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, S.; Ogasawara, Y.; Shibuya, N.; Kimura, H.; Ishii, K. Polysulfide Exerts a Protective Effect against Cytotoxicity Caused by T-Buthylhydroperoxide through Nrf2 Signaling in Neuroblastoma Cells. FEBS Lett. 2013, 587, 3548–3555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarosz, A.P.; Wei, W.; Gauld, J.W.; Auld, J.; Özcan, F.; Aslan, M.; Mutus, B. Glyceraldehyde 3-Phosphate Dehydrogenase (GAPDH) Is Inactivated by S-Sulfuration In Vitro. Free Radic. Biol. Med. 2015, 89, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.R.; Melideo, S.L.; Jorns, M.S. Human Sulfide:Quinone Oxidoreductase Catalyzes the First Step in Hydrogen Sulfide Metabolism and Produces a Sulfane Sulfur Metabolite. Biochemistry 2012, 51, 6804–6815. [Google Scholar] [CrossRef]
- Libiad, M.; Yadav, P.K.; Vitvitsky, V.; Martinov, M.; Banerjee, R. Organization of the Human Mitochondrial Hydrogen Sulfide Oxidation Pathway. J. Biol. Chem. 2014, 289, 30901–30910. [Google Scholar] [CrossRef] [Green Version]
- Yadav, P.K.; Martinov, M.; Vitvitsky, V.; Seravalli, J.; Wedmann, R.; Filipovic, M.R.; Banerjee, R. Biosynthesis and Reactivity of Cysteine Persulfides in Signaling. J. Am. Chem. Soc. 2016, 138, 289–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, K.; Akaike, T.; Sawa, T.; Kumagai, Y.; Wink, D.A.; Tantillo, D.J.; Hobbs, A.J.; Nagy, P.; Xian, M.; Lin, J.; et al. Redox Chemistry and Chemical Biology of H2S, Hydropersulfides, and Derived Species: Implications of Their Possible Biological Activity and Utility. Free Radic. Biol. Med. 2014, 77, 82–94. [Google Scholar] [CrossRef] [Green Version]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive Cysteine Persulfides and S-Polythiolation Regulate Oxidative Stress and Redox Signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, C.E.; Carroll, K.S. Cysteine-Mediated Redox Signaling: Chemistry, Biology, and Tools for Discovery. Chem. Rev. 2013, 113, 4633–4679. [Google Scholar] [CrossRef]
- Carballal, S.; Radi, R.; Kirk, M.C.; Barnes, S.; Freeman, B.A.; Alvarez, B. Sulfenic Acid Formation in Human Serum Albumin by Hydrogen Peroxide and Peroxynitrite. Biochemistry 2003, 42, 9906–9914. [Google Scholar] [CrossRef]
- Zhang, D.; Macinkovic, I.; Devarie-Baez, N.O.; Pan, J.; Park, C.-M.; Carroll, K.S.; Filipovic, M.R.; Xian, M. Detection of Protein S-Sulfhydration by a Tag-Switch Technique. Angew. Chem. Int. Ed. Engl. 2014, 53, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talipov, M.R.; Timerghazin, Q.K. Protein Control of S-Nitrosothiol Reactivity: Interplay of Antagonistic Resonance Structures. J. Phys. Chem. B 2013, 117, 1827–1837. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic, J.L.; Kenkel, I.; Ivanović-Burmazović, I.; Filipovic, M.R. Generation of HNO and HSNO from Nitrite by Heme-Iron-Catalyzed Metabolism with H2S. Angew. Chem. Int. Ed. Engl. 2013, 52, 12061–12064. [Google Scholar] [CrossRef] [PubMed]
- Lajin, B.; Francesconi, K.A. The Hydrogen Sulfide Metabolite Trimethylsulfonium Is Found in Human Urine. Sci. Rep. 2016, 6, 27038. [Google Scholar] [CrossRef] [Green Version]
- Snel, J.; Burgering, M.; Smit, B.; Noordman, W.; Tangerman, A.; Winkel, E.G.; Kleerebezem, M. Volatile Sulphur Compounds in Morning Breath of Human Volunteers. Arch. Oral. Biol. 2011, 56, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, M.A.; Alasqah, M.; Sanaa, M.S.; Gufran, K. The Relationship between Volatile Sulfur Compounds and the Severity of Chronic Periodontitis: A Cross-Sectional Study. J. Pharm. Bioallied Sci. 2020, 12, S268–S273. [Google Scholar] [CrossRef]
- Yoo, S.H.; Jung, H.S.; Sohn, W.S.; Kim, B.H.; Ku, B.H.; Kim, Y.S.; Park, S.W.; Hahm, K.-B. Volatile Sulfur Compounds as a Predictor for Esophagogastroduodenal Mucosal Injury. Gut Liver 2008, 2, 113–118. [Google Scholar] [CrossRef]
- Guan, X.; Wu, P.; Wang, S.; Zhang, J.; Shen, Z.; Luo, H.; Chen, H.; Long, L.; Chen, J.; Wang, F. Dimethyl Sulfide Protects against Oxidative Stress and Extends Lifespan via a Methionine Sulfoxide Reductase A-dependent Catalytic Mechanism. Aging Cell 2017, 16, 226–236. [Google Scholar] [CrossRef]
- Lim, J.C.; You, Z.; Kim, G.; Levine, R.L. Methionine Sulfoxide Reductase A Is a Stereospecific Methionine Oxidase. Proc. Nat. Acad. Sci. USA 2011, 108, 10472–10477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anwar, A.; Burkholz, T.; Scherer, C.; Abbas, M.; Lehr, C.-M.; Diederich, M.; Jacob, C. Naturally Occurring Reactive Sulfur Species, Their Activity against Caco-2 Cells, and Possible Modes of Biochemical Action. J. Sulfur. Chem. 2008, 29, 251–268. [Google Scholar] [CrossRef]
- Grman, M.; Nasim, M.J.; Leontiev, R.; Misak, A.; Jakusova, V.; Ondrias, K.; Jacob, C. Inorganic Reactive Sulfur-Nitrogen Species: Intricate Release Mechanisms or Cacophony in Yellow, Blue and Red? Antioxidants 2017, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasim, M.J.; Denezhkin, P.; Sarfraz, M.; Leontiev, R.; Ney, Y.; Kharma, A.; Griffin, S.; Masood, M.I.; Jacob, C. The Small Matter of a Red Ox, a Particularly Sensitive Pink Cat, and the Quest for the Yellow Stone of Wisdom. Curr. Pharmacol. Rep. 2018, 4, 380–396. [Google Scholar] [CrossRef]
- Allah, D.R.; Schwind, L.; Asali, I.A.; Nasim, J.; Jacob, C.; Götz, C.; Montenarh, M. A Scent of Therapy: Synthetic Polysulfanes with Improved Physico-Chemical Properties Induce Apoptosis in Human Cancer Cells. Int. J. Oncol. 2015, 47, 991–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czepukojc, B.; Leroch, M.; Salm, F.; Viswanathan, U.M.; Burkholz, T.; Hahn, M.; Jacob, C. Antifungal Activity of Tetrasulfanes against Botrytis Cinerea. Nat. Prod. Commun. 2013, 8, 1599–1603. [Google Scholar] [CrossRef] [Green Version]
- Marut, W.; Jamier, V.; Kavian, N.; Servettaz, A.; Winyard, P.G.; Eggleton, P.; Anwar, A.; Nicco, C.; Jacob, C.; Chéreau, C.; et al. The Natural Organosulfur Compound Dipropyltetrasulfide Prevents HOCl-Induced Systemic Sclerosis in the Mouse. Arthritis Res. Ther. 2013, 15, R167. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, S.; Lee, K.-J.; Shukla, P.; Chae, J.-C. Dimethyl Disulfide Exerts Antifungal Activity against Sclerotinia Minor by Damaging Its Membrane and Induces Systemic Resistance in Host Plants. Sci. Rep. 2020, 10, 6547. [Google Scholar] [CrossRef] [Green Version]
- Dandurishvili, N.; Toklikishvili, N.; Ovadis, M.; Eliashvili, P.; Giorgobiani, N.; Keshelava, R.; Tediashvili, M.; Vainstein, A.; Khmel, I.; Szegedi, E.; et al. Broad-Range Antagonistic Rhizobacteria Pseudomonas Fluorescens and Serratia Plymuthica Suppress Agrobacterium Crown Gall Tumours on Tomato Plants. J. Appl. Microbiol. 2011, 110, 341–352. [Google Scholar] [CrossRef]
- Lammers, A.; Lalk, M.; Garbeva, P. Air Ambulance: Antimicrobial Power of Bacterial Volatiles. Antibiotics 2022, 11, 109. [Google Scholar] [CrossRef]
- Paladin®EC Soil Fumigant, Active Ingredient Dimethyl Disulfide (DMDS), Drip Application. Available online: https://www.arkema.com/global/en/products/product-finder/product-range/thiochemicals/dmds-for-agricultural-soil-fumig/ (accessed on 23 March 2022).
- Chernin, L.; Toklikishvili, N.; Ovadis, M.; Kim, S.; Ben-Ari, J.; Khmel, I.; Vainstein, A. Quorum-Sensing Quenching by Rhizobacterial Volatiles. Environ. Microbiol. Rep. 2011, 3, 698–704. [Google Scholar] [CrossRef]
- Tang, Y.; Li, F.; Gu, D.; Wang, W.; Huang, J.; Jiao, X. Antimicrobial Effect and the Mechanism of Diallyl Trisulfide against Campylobacter Jejuni. Antibiotics 2021, 10, 246. [Google Scholar] [CrossRef]
- Gong, X.; Su, X.; Liu, H. Diallyl Trisulfide, the Antifungal Component of Garlic Essential Oil and the Bioactivity of Its Nanoemulsions Formed by Spontaneous Emulsification. Molecules 2021, 26, 7186. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.-L.; Ma, Z.-Y.; Wang, B.; Gao, F.; Yi, C.-G.; Zhou, X.-X.; Guo, S.-Y.; Zhou, L. In Vitro Anti-Synovial Sarcoma Effect of Diallyl Trisulfide and MRNA Profiling. Gene 2022, 816, 146172. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.; Midde, N.M.; Miller, D.D.; Chauhan, S.; Kumar, A.; Kumar, S. Diallyl Sulfide: Potential Use in Novel Therapeutic Interventions in Alcohol, Drugs, and Disease Mediated Cellular Toxicity by Targeting Cytochrome P450 2E1. Curr. Drug Metab. 2015, 16, 486–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ecospray|Ecospray Is the Pioneer of Internationally Approved and Patented Crop Protection Products Formulated Using a 100% Safe, Natural and Sustainable Technology Derived from Garlic. Available online: https://ecospray.com/ (accessed on 23 March 2022).
- Zhou, W.; Zou, X.; Lu, Y.; Xia, L.; Huang, C.; Shen, C.; Chen, X.; Chu, Y. Characterization of Volatiles in Garlic and in Exhaled Breath after Garlic Ingestion by On-Line Atmospheric Pressure Photoionization Quadrupole Time-of-Flight Mass Spectrometry. Food Sci. Technol. Res. 2017, 23, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Taucher, J.; Hansel, A.; Jordan, A.; Lindinger, W. Analysis of Compounds in Human Breath after Ingestion of Garlic Using Proton-Transfer-Reaction Mass Spectrometry. J. Agric. Food Chem. 1996, 44, 3778–3782. [Google Scholar] [CrossRef]
- Sato, S.; Sekine, Y.; Kakumu, Y.; Hiramoto, T. Measurement of Diallyl Disulfide and Allyl Methyl Sulfide Emanating from Human Skin Surface and Influence of Ingestion of Grilled Garlic. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Laakso, I.; Seppänen-Laakso, T.; Hiltunen, R.; Müller, B.; Jansen, H.; Knobloch, K. Volatile Garlic Odor Components: Gas Phases and Adsorbed Exhaled Air Analysed by Headspace Gas Chromatography-Mass Spectrometry. Planta Med. 1989, 55, 257–261. [Google Scholar] [CrossRef]
- Lawson, L.D.; Wang, Z.J. Allicin and Allicin-Derived Garlic Compounds Increase Breath Acetone through Allyl Methyl Sulfide: Use in Measuring Allicin Bioavailability. J. Agric. Food Chem. 2005, 53, 1974–1983. [Google Scholar] [CrossRef]
- Lawson, L.D.; Hunsaker, S.M. Allicin Bioavailability and Bioequivalence from Garlic Supplements and Garlic Foods. Nutrients 2018, 10, 812. [Google Scholar] [CrossRef] [Green Version]
- Kuttan, G.; Kuttan, R. Effect of Diallyl Sulphide, Diallyl Disulphide, and Allyl Methyl Sulphide on the Inhibition of Lung Metastasis of B16F-10 Melanoma Cells in Mice. J. Clin. Biochem. Nutr. 1999, 27, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Sinha, A.K.; Farooqui, S.A.; Sharma, A.; Mishra, A.; Verma, V. Reactivity of Allyl Methyl Sulphide, the in-Vitro Metabolite of Garlic, with Some Amino Acids and with Phospholipid Involved in Viral Infections. J. Biomol. Struct. Dyn. 2022, 40, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Becker, P.M.; van Wikselaar, P.G.; Mul, M.F.; Pol, A.; Engel, B.; Wijdenes, J.W.; van der Peet-Schwering, C.M.C.; Wisselink, H.J.; Stockhofe-Zurwieden, N. Actinobacillus Pleuropneumoniae Is Impaired by the Garlic Volatile Allyl Methyl Sulfide (AMS) in Vitro and in-Feed Garlic Alleviates Pleuropneumonia in a Pig Model. Vet Microbiol. 2012, 154, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, Y.; Matsumura, S.; Morimoto, M.; Takemoto, Y.; Kishi, C.; Moriyama, T.; Zaima, N. Inhibitory Activities of Sulfur Compounds in Garlic Essential Oil against Alzheimer’s Disease-Related Enzymes and Their Distribution in the Mouse Brain. J. Agric. Food Chem. 2021, 69, 10163–10173. [Google Scholar] [CrossRef] [PubMed]
- Lietzow, J. Biologically Active Compounds in Mustard Seeds: A Toxicological Perspective. Foods 2021, 10, 2089. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Khatun, A.; Liu, L.; Barkla, B.J. Brassicaceae Mustards: Traditional and Agronomic Uses in Australia and New Zealand. Molecules 2018, 23, 231. [Google Scholar] [CrossRef] [Green Version]
- Striegel, L.; Chebib, S.; Dumler, C.; Lu, Y.; Huang, D.; Rychlik, M. Durian Fruits Discovered as Superior Folate Sources. Front. Nutr. 2018, 5, 114. [Google Scholar] [CrossRef] [Green Version]
- Husin, N.A.; Rahman, S.; Karunakaran, R.; Bhore, S.J. A Review on the Nutritional, Medicinal, Molecular and Genome Attributes of Durian (Durio zibethinus L.), the King of Fruits in Malaysia. Bioinformation 2018, 14, 265–270. [Google Scholar] [CrossRef]
- Weenen, H.; Koolhaas, W.E.; Apriyantono, A. Sulfur-Containing Volatiles of Durian Fruits (Durio zibethinus Murr.). J. Agric. Food Chem. 1996, 44, 3291–3293. [Google Scholar] [CrossRef]
- Aziz, N.A.A.; Mhd Jalil, A.M. Bioactive Compounds, Nutritional Value, and Potential Health Benefits of Indigenous Durian (Durio zibethinus Murr.): A Review. Foods 2019, 8, 96. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
In mammals, H2S is produced via several pathways. Cystathionine-β-synthase (CBS) and cystathionine-γ-lyase (CSE) interact with cysteine to produce H2S following the reverse transsulfuration pathway and α, β-elimination, respectively [39,40]. The production of H2S also involves enzymes such as methionine-γ-lyase and cysteine desulfhydrase [24]. The scheme is superimposed on a photo of the Elisenbrunnen in the German city of Aachen, a popular source of sulfur-rich water drunk frequently by the local population (Photo credit to Muhammad Jawad Nasim) [41].
Figure 1.
In mammals, H2S is produced via several pathways. Cystathionine-β-synthase (CBS) and cystathionine-γ-lyase (CSE) interact with cysteine to produce H2S following the reverse transsulfuration pathway and α, β-elimination, respectively [39,40]. The production of H2S also involves enzymes such as methionine-γ-lyase and cysteine desulfhydrase [24]. The scheme is superimposed on a photo of the Elisenbrunnen in the German city of Aachen, a popular source of sulfur-rich water drunk frequently by the local population (Photo credit to Muhammad Jawad Nasim) [41].

Figure 2.
In humans, SO2 is produced from L-cysteine and metabolized to SO32− and SO42−, which are excreted via the urine. A brief list of biological activities is provided and more details can be found in the text [62,63,64].

Figure 3.
Allyl and propyl mono- and polysulfides found in garlic and onion may serve as multifunctional agents able to interact with thiol residues of proteins and enzymes, interfere with metal homeostasis, bind to metals, and induce oxidative stress via oxidative interactions.
Figure 3.
Allyl and propyl mono- and polysulfides found in garlic and onion may serve as multifunctional agents able to interact with thiol residues of proteins and enzymes, interfere with metal homeostasis, bind to metals, and induce oxidative stress via oxidative interactions.

Figure 4.
Generic pathway for the conversion of allyl thiosulfinates and polysulfanes to AMS. Allyl polysulfides are metabolized mainly to allyl methyl sulfide with allyl mercaptan as an intermediate, see text for details.
Figure 4.
Generic pathway for the conversion of allyl thiosulfinates and polysulfanes to AMS. Allyl polysulfides are metabolized mainly to allyl methyl sulfide with allyl mercaptan as an intermediate, see text for details.

Figure 5.
A brief overview of volatile sulfur species found in natural products such as mustard, cruciferous vegetables, coffee, and shiitake mushroom (a), and in durian fruit (b).
Figure 5.
A brief overview of volatile sulfur species found in natural products such as mustard, cruciferous vegetables, coffee, and shiitake mushroom (a), and in durian fruit (b).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tiganescu, E.; Lämmermann, M.A.; Ney, Y.; Abdin, A.Y.; Nasim, M.J.; Jacob, C. A Whiff of Sulfur: One Wind a Day Keeps the Doctor Away. Antioxidants 2022, 11, 1036. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11061036
AMA Style
Tiganescu E, Lämmermann MA, Ney Y, Abdin AY, Nasim MJ, Jacob C. A Whiff of Sulfur: One Wind a Day Keeps the Doctor Away. Antioxidants. 2022; 11(6):1036. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11061036
Chicago/Turabian StyleTiganescu, Eduard, Markus Alexander Lämmermann, Yannick Ney, Ahmad Yaman Abdin, Muhammad Jawad Nasim, and Claus Jacob. 2022. "A Whiff of Sulfur: One Wind a Day Keeps the Doctor Away" Antioxidants 11, no. 6: 1036. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11061036
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.