
Antioxidant Effect of Tyr-Ala Extracted from Zein on INS-1 Cells and Type 2 Diabetes High-Fat-Diet-Induced Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. MTT Reduction Assay
2.4. Reactive Oxygen Species (ROS) Assay
2.5. Insulin Secretion Assay
2.6. Western Blot
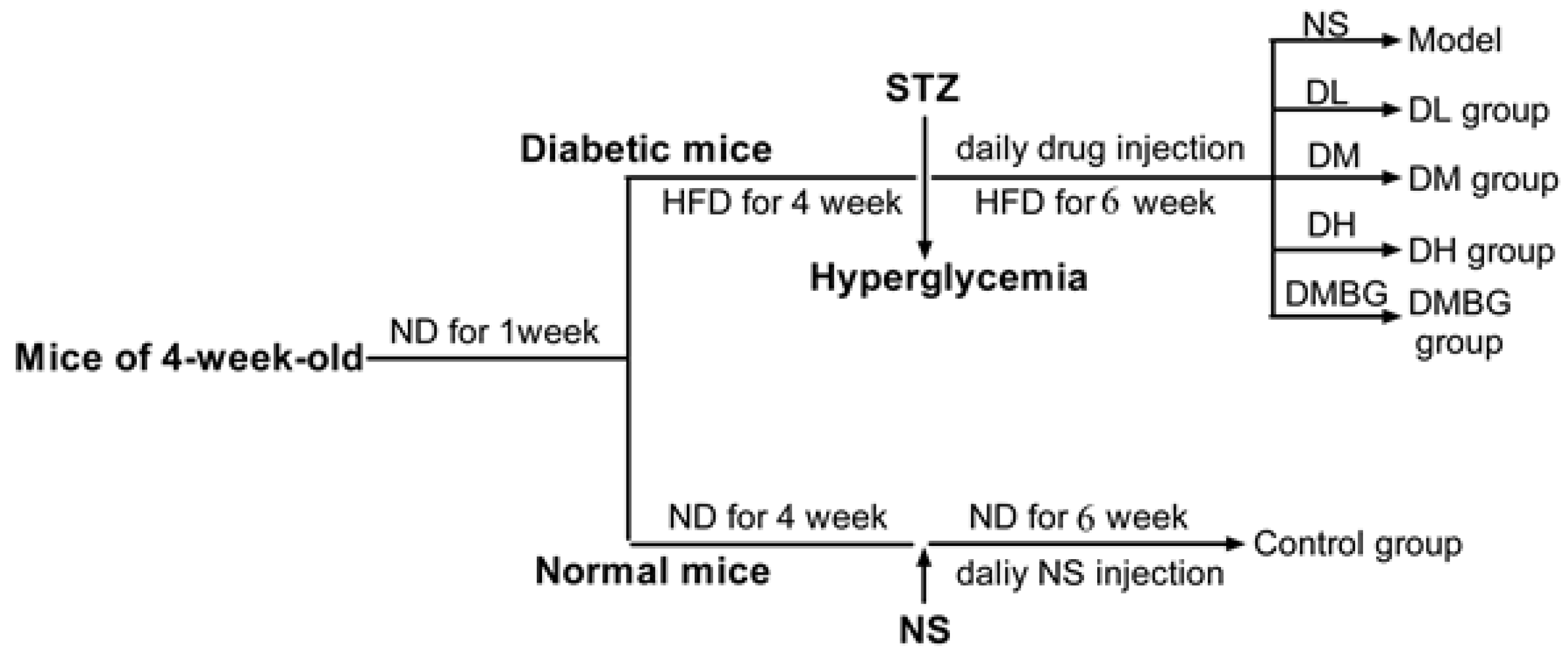
2.7. Experimental Animals
2.8. Body Weight and Fasting Blood Glucose Assay
2.9. Intraperitoneal Glucose Tolerance Test (IPGTT)
2.10. Biochemical Assay
2.11. Histopathological Examination
2.12. Statistical Analysis
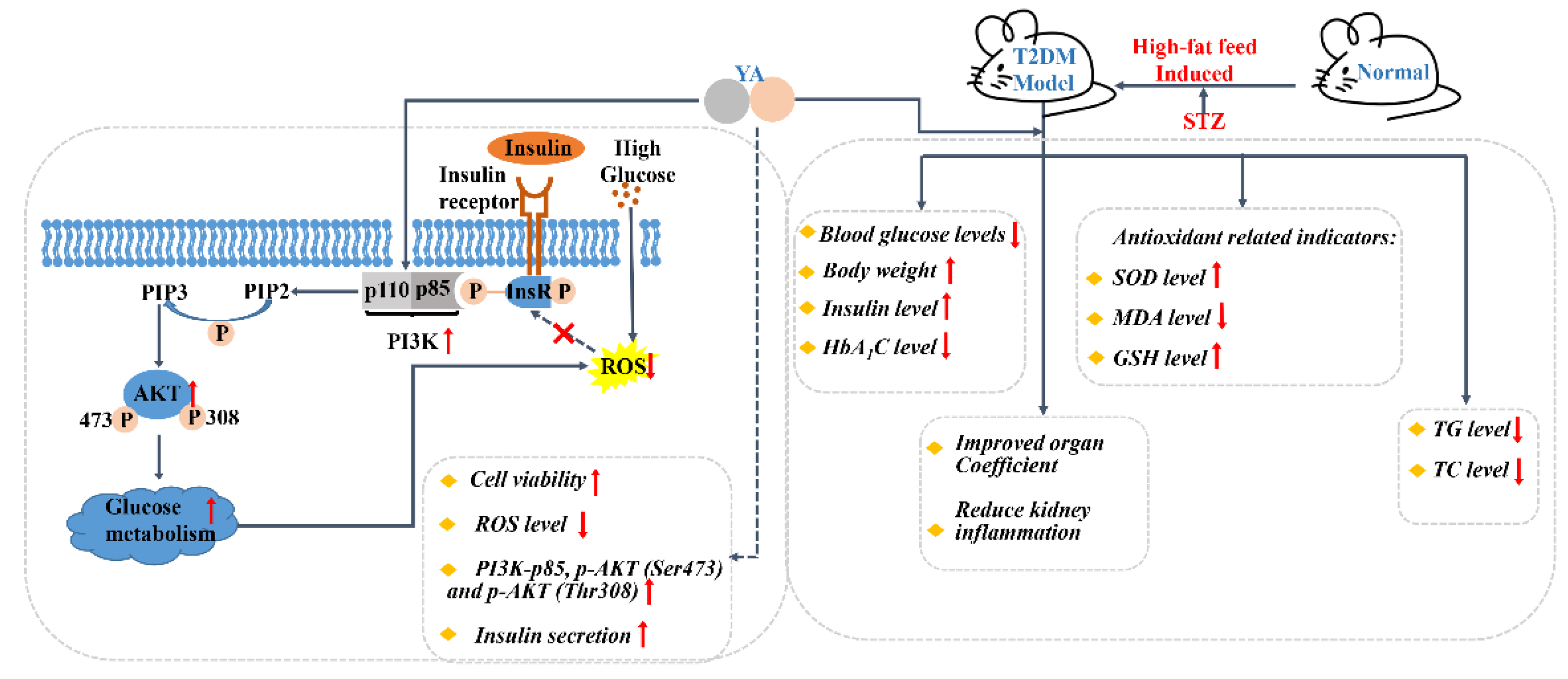
3. Results
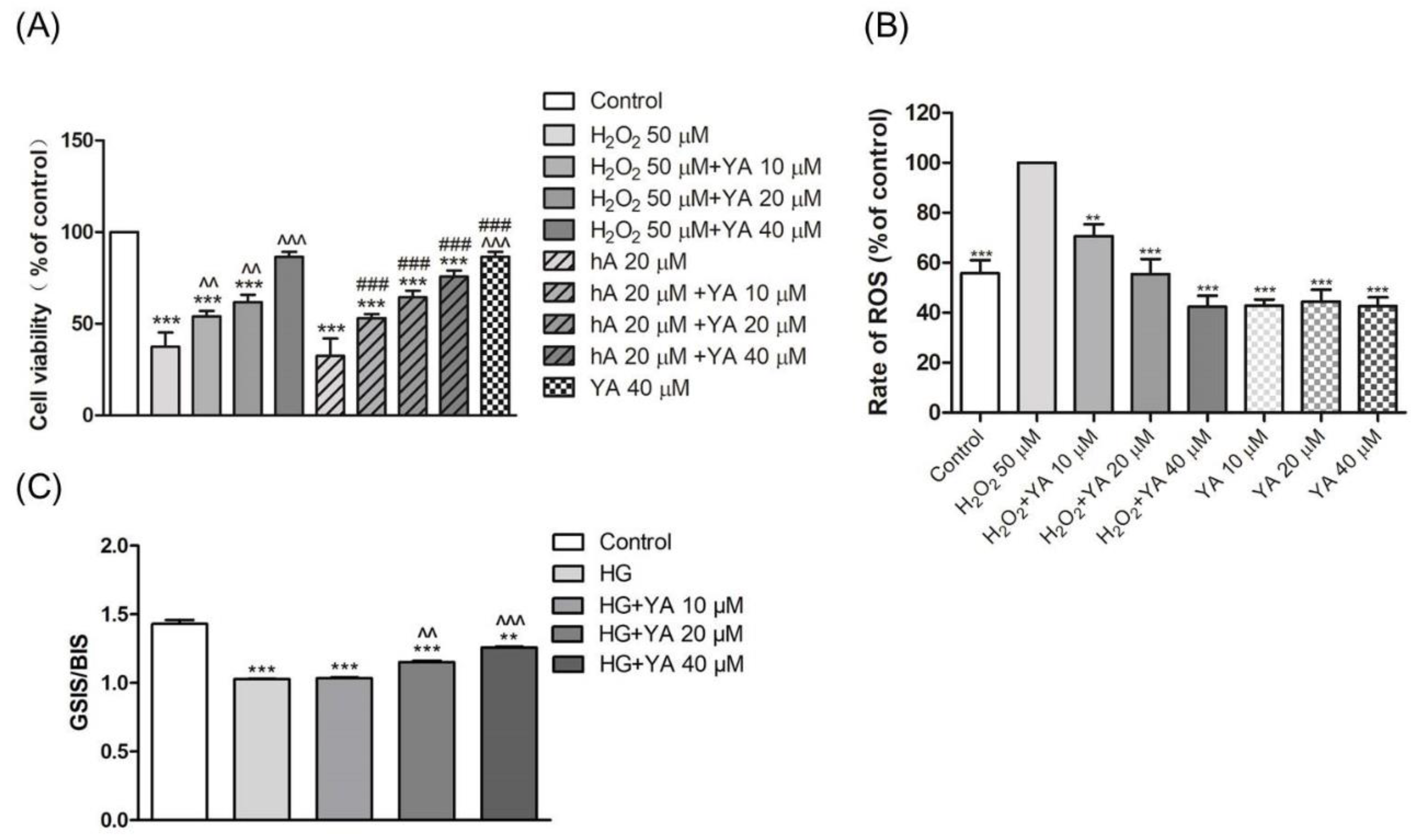
3.1. Effect of YA on INS-1 Cell Viability
3.2. Effect of YA on ROS Accumulation in INS-1 Cells
3.3. Effect of YA on Insulin Secretion
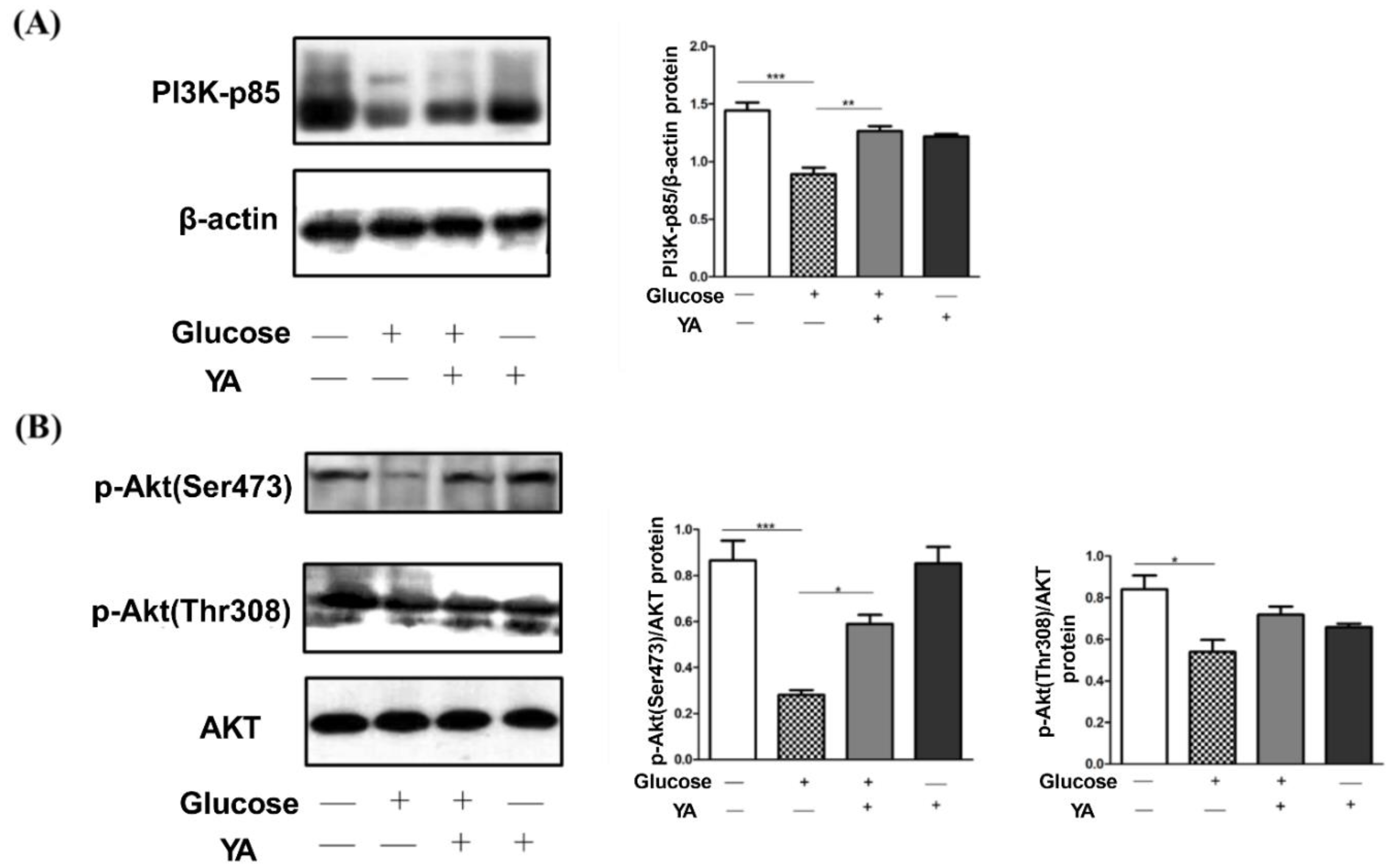
3.4. Regulation of YA on PI3K/Akt Signal Pathway
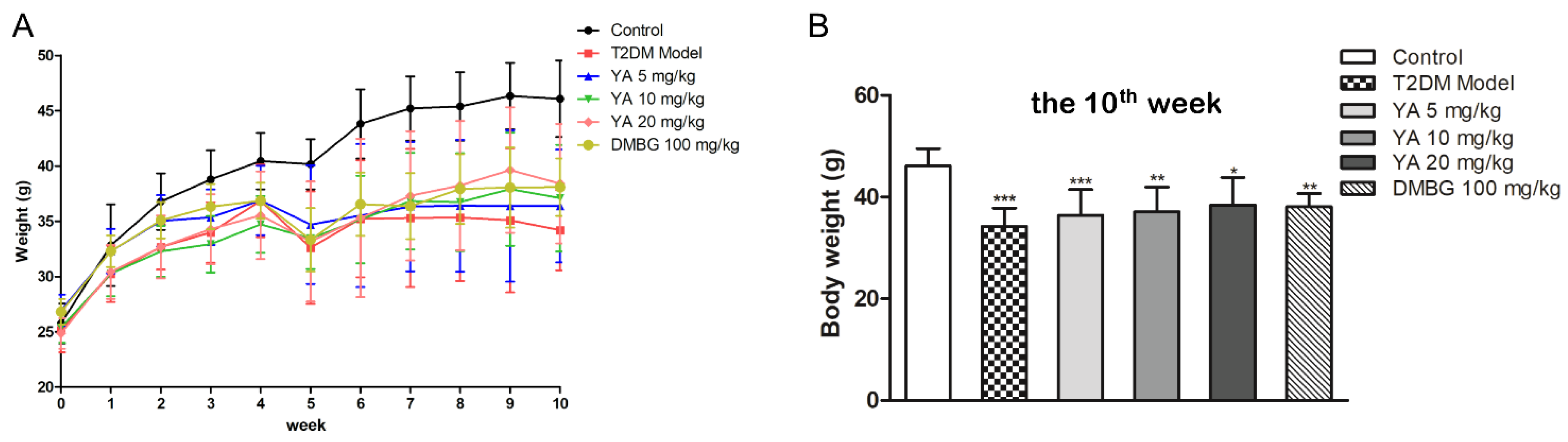
3.5. Effect of YA on General Condition and Weight of Mice
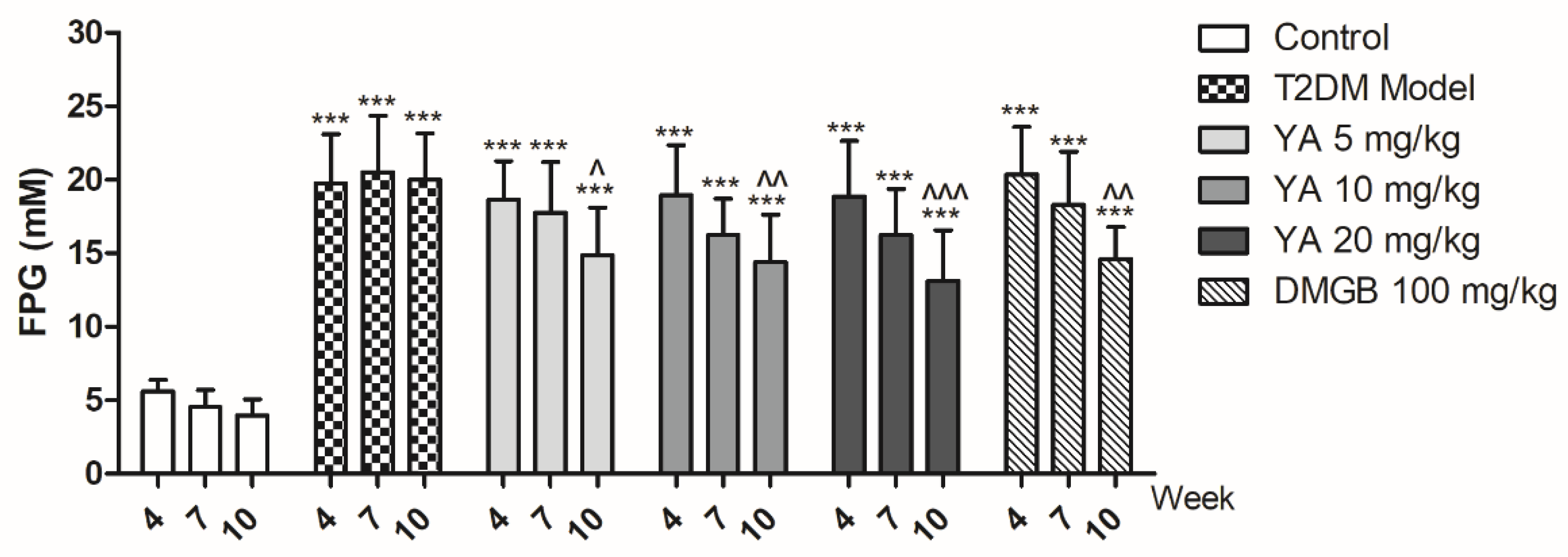
3.6. YA Decrease Blood Glucose Levels in T2DM Mice
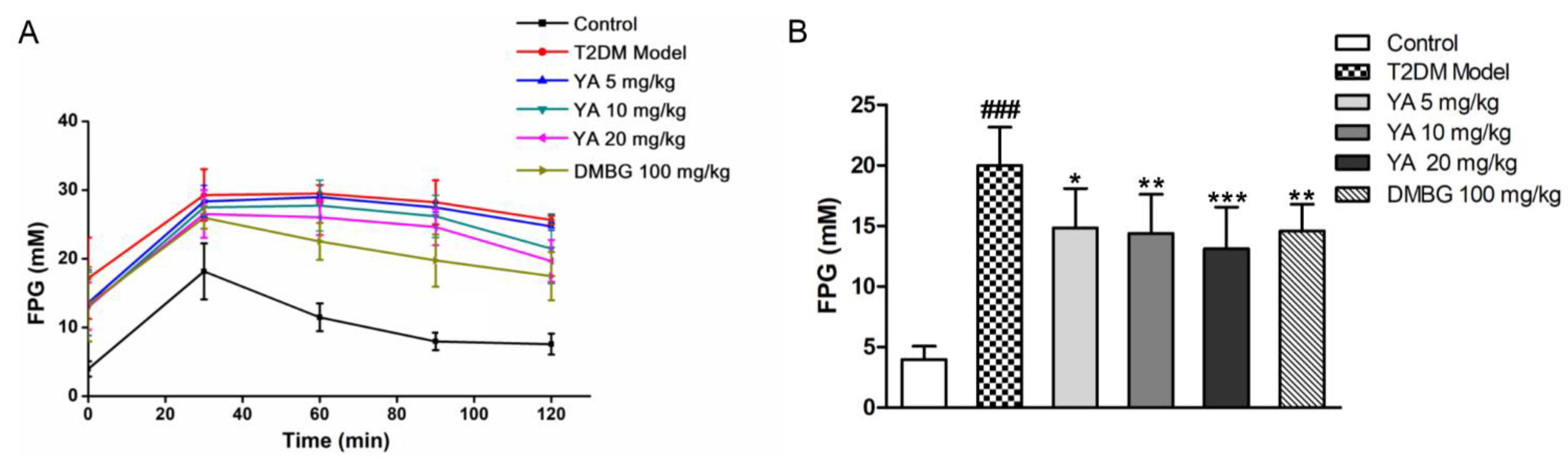
3.7. Effect of YA on Glucose Tolerance of T2DM Mice
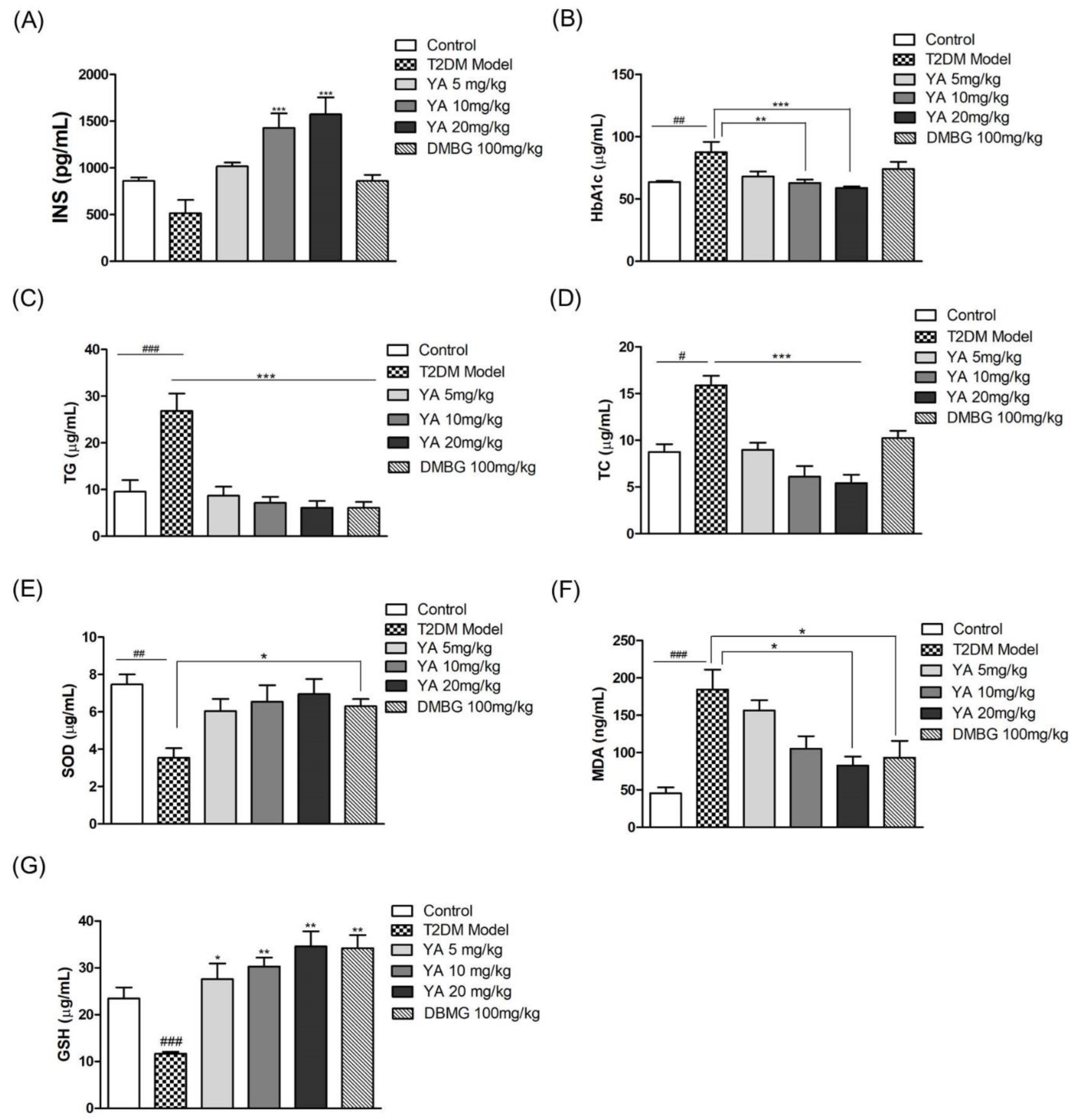
3.8. Effect of YA on Insulin and HbA1c Levels
3.9. Effect of YA on TG and TC Levels
3.10. Effect of YA on Levels of SOD, MDA, and GSH
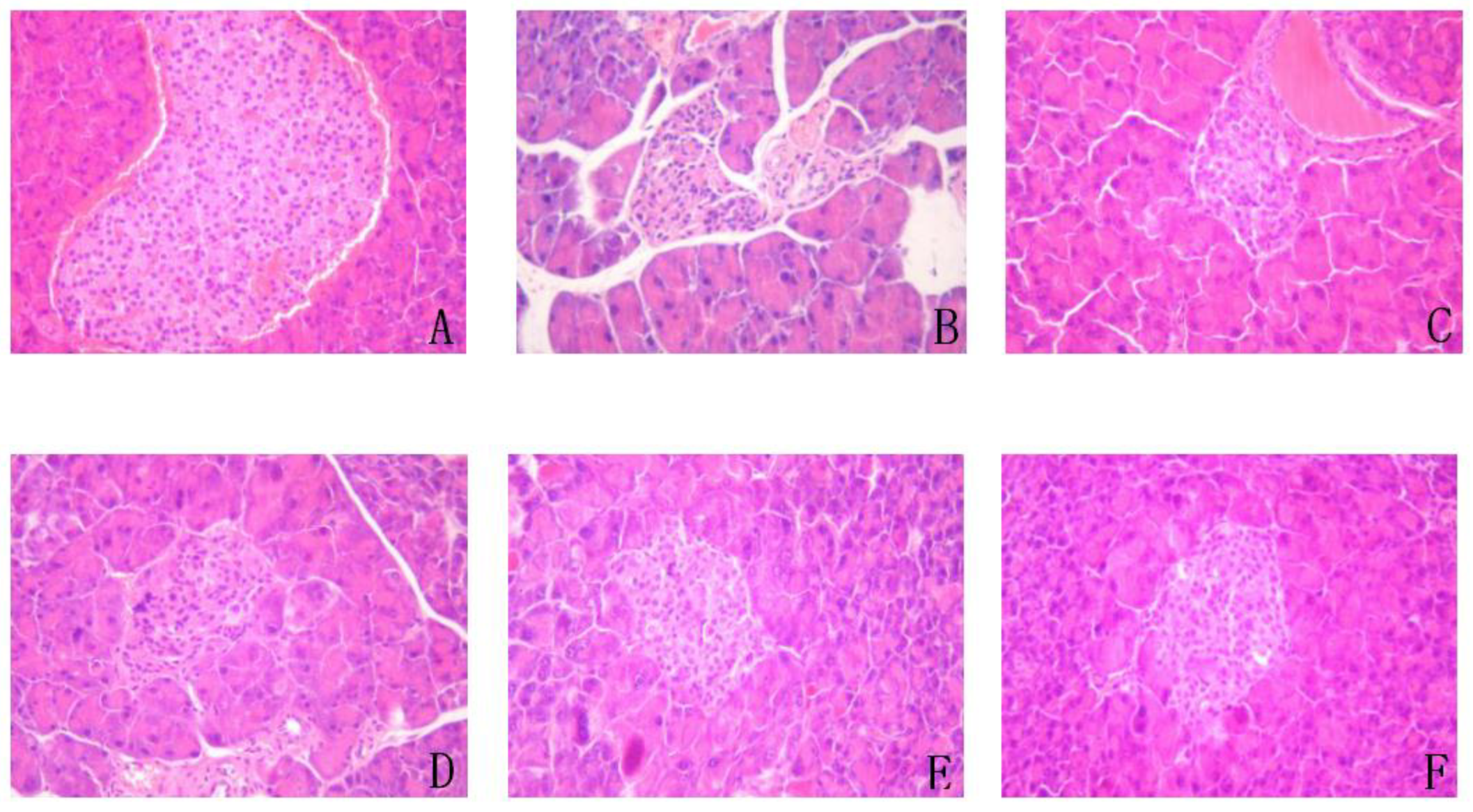
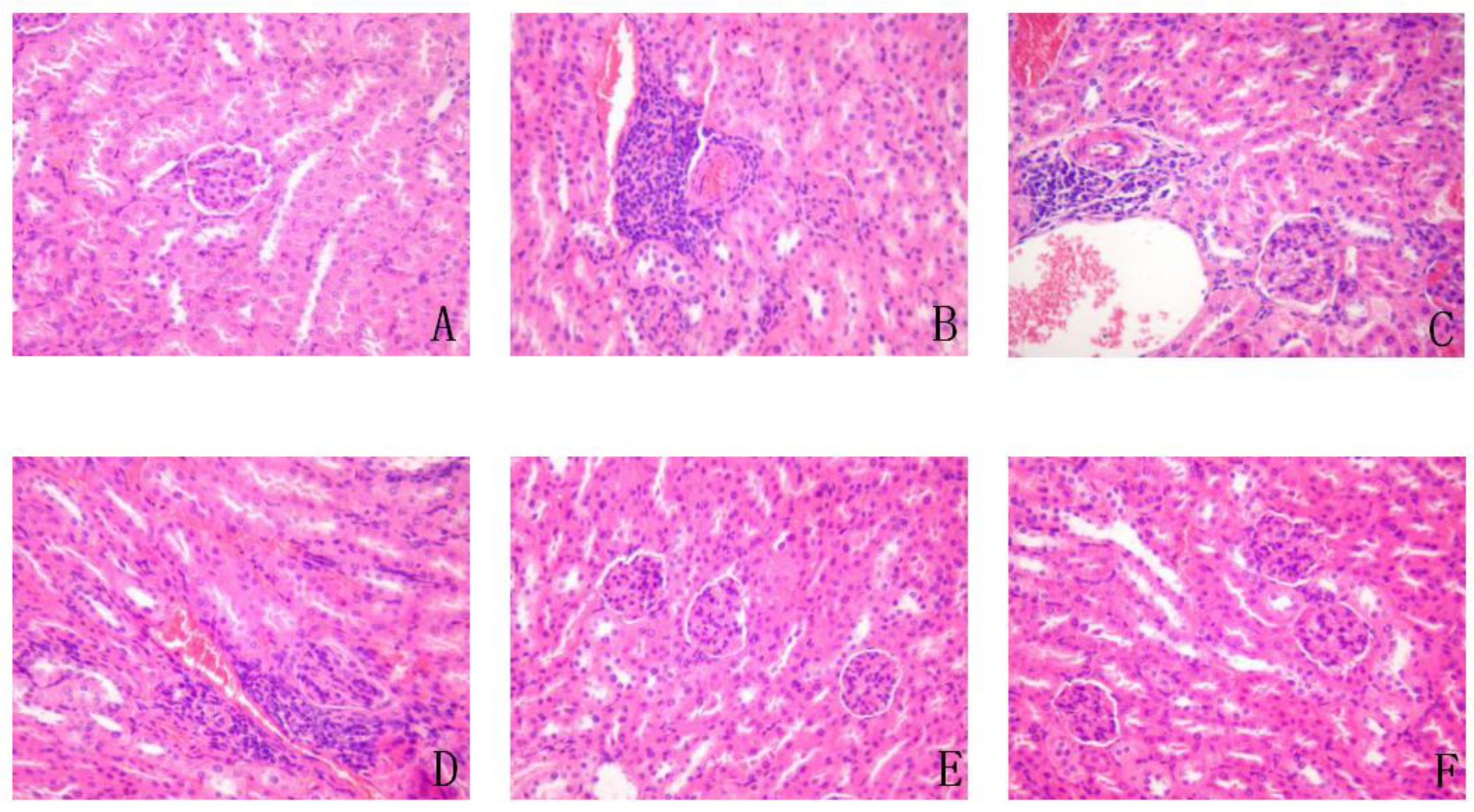
3.11. Effect on Liver, Spleen, Pancreas, and Kidney
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malecki, M.T. Type 2 Diabetes Mellitus and Its Complications: From the Molecular Biology to the Clinical Practice. Rev. Diabet. Stud. 2004, 1, 5–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juarez-Flores, D.L.; Ezquerra, M.; Gonzalez-Casacuberta, I.; Ormazabal, A.; Moren, C.; Tolosa, E.; Fucho, R.; Guitart-Mampel, M.; Casado, M.; Valldeoriola, F.; et al. Disrupted Mitochondrial and Metabolic Plasticity Underlie Comorbidity between Age-Related and Degenerative Disorders as Parkinson Disease and Type 2 Diabetes Mellitus. Antioxidants 2020, 9, 1063. [Google Scholar] [CrossRef] [PubMed]
- Dowlaty, N.; Yoon, A.; Galassetti, P. Monitoring States of Altered Carbohydrate Metabolism via Breath Analysis: Are Times Ripe for Transition from Potential to Reality? Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 466–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etsassala, N.G.E.R.; Badmus, J.A.; Marnewick, J.L.; Egieyeh, S.; Iwuoha, E.I.; Nchu, F.; Hussein, A.A. Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities, Molecular Docking, and Antioxidant Capacities of Plectranthus ecklonii Constituents. Antioxidants 2022, 11, 378. [Google Scholar] [CrossRef]
- Chen, H.Y.; Chiang, Y.F.; Wang, K.L.; Huang, T.C.; Ali, M.; Shieh, T.M.; Chang, H.Y.; Hong, Y.H.; Hsia, S.M. Rice Husk Silica Liquid Protects Pancreatic beta Cells from Streptozotocin-Induced Oxidative Damage. Antioxidants 2021, 10, 1080. [Google Scholar] [CrossRef]
- Perry, R.J.; Samuel, V.T.; Petersen, K.F.; Shulman, G.I. The Role of Hepatic Lipids in Hepatic Insulin Resistance and type 2 Diabetes. Nature 2014, 510, 84–91. [Google Scholar] [CrossRef]
- Lee, M.S.; Chyau, C.C.; Wang, C.P.; Wang, T.H.; Chen, J.H.; Lin, H.H. Flavonoids Identification and Pancreatic Beta-Cell Protective Effect of Lotus Seedpod. Antioxidants 2020, 9, 658. [Google Scholar] [CrossRef]
- Simmons, R.A.; Suponitsky-Kroyter, I.; Selak, M.A. Progressive Accumulation of Mitochondrial DNA Mutations and Decline in Mitochondrial Function Lead to β-Cell Failure. J. Biol. Chem. 2005, 280, 28785–28791. [Google Scholar] [CrossRef] [Green Version]
- Martens, G.A.; Cai, Y.; Hinke, S.; Stangé, G.; Van de Casteele, M.; Pipeleers, D. Glucose Suppresses Superoxide Generation in Metabolically Responsive Pancreatic Beta Cells. J. Biol. Chem. 2005, 280, 20389–20396. [Google Scholar] [CrossRef] [Green Version]
- Maestre, I.; Jordán, J.; Calvo, S.; Reig, J.A.; Ceña, V.; Soria, B.; Prentki, M.; Roche, E. Mitochondrial Dysfunction Is Involved in Apoptosis Induced by Serum Withdrawal and Fatty Acids in the Beta-Cell Line INS-1. Endocrinology 2003, 144, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee Bhowmick, D.; Burnett, L.; Kudaibergenova, Z.; Jeremic, A.M. FoxA2 and RNA Pol II mediate human islet amyloid polypeptide turnover in ER-stressed pancreatic beta-cells. Biochem. J. 2021, 478, 1261–1282. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.A.; Lewin, T.M.; Van Horn, C.G.; Gonzalez-Baró, M.R. Do Long-Chain Acyl-CoA Synthetases Regulate Fatty Acid Entry Into Synthetic Versus Degradative Pathways? J. Nutr. 2002, 132, 2123–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoval, H.; Weiner, L.; Gazit, E.; Levy, M.; Pinchuk, I.; Lichtenberg, D. Polyphenol-Induced Dissociation of Various Amyloid Fibrils Results in a Methionine-Independent Formation of ROS. Biochim. Biophys. Acta 2008, 1784, 1570–1577. [Google Scholar] [CrossRef]
- Kahn, S.E.; Andrikopoulos, S.; Verchere, C.B. Islet Amyloid: A Long-Recognized but Underappreciated Pathological Feature of type 2 Diabetes. Diabetes 1999, 48, 241–253. [Google Scholar] [CrossRef]
- Ye, X.; Chen, W.; Tu, P.; Jia, R.; Liu, Y.; Li, Y.; Tang, Q.; Zheng, X.; Chu, Q. Food-derived cyanidin-3-O-glucoside alleviates oxidative stress: Evidence from the islet cell line and diabetic db/db mice. Food Funct. 2021, 12, 11599–11610. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Gomez, J.L.; Ortiz-Martinez, M.; Aguilar, O.; Garcia-Lara, S.; Castorena-Torres, F. Antioxidant Activity of Zein Hydrolysates from Zea Species and Their Cytotoxic Effects in a Hepatic Cell Culture. Molecules 2018, 23, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chen, H.; Wang, X.; Li, S.; Chen, Z.; Wang, J.; Liu, W. Isolation and Identification of a Novel Peptide From zein With Antioxidant and Antihypertensive Activities. Food Funct. 2015, 6, 3799–3806. [Google Scholar] [CrossRef]
- Tang, X.Y.; He, Z.Y.; Dai, Y.F.; Xiong, Y.L.; Xie, M.Y.; Chen, J. Peptide Fractionation and Free Radical Scavenging Activity of zein Hydrolysate. J. Agric. Food Chem. 2010, 58, 587–593. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, Y.; Wang, X.; Lin, R.; Zhang, Y.; Ma, H.; Guo, Y.; Xu, L.; Zhao, B. The novel dipeptide Tyr-Ala (TA) significantly enhances the lifespan and healthspan of Caenorhabditis elegans. Food Funct. 2016, 7, 1975–1984. [Google Scholar] [CrossRef]
- Lee, E.C.; Ha, E.; Singh, S.; Legesse, L.; Ahmad, S.; Karnaukhova, E.; Donaldson, R.P.; Jeremic, A.M. Copper (II)-Human Amylin Complex Protects Pancreatic Cells from Amylin Toxicity. Phys. Chem. Chem. Phys. 2013, 15, 12558–12571. [Google Scholar] [CrossRef] [Green Version]
- Lin, N.; Chen, H.; Zhang, H.; Wan, X.; Su, Q. Mitochondrial Reactive Oxygen Species (ROS) Inhibition Ameliorates Palmitate-Induced INS-1 Beta Cell Death. Endocrine 2012, 42, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Sabra, G.; Dubiel, E.A.; Kuehn, C.; Khalfaoui, T.; Beaulieu, J.F.; Vermette, P. INS-1 Cell Glucose-Stimulated Insulin Secretion Is Reduced by the Downregulation of the 67 kDa Laminin Receptor. J. Tissue Eng. Regen. Med. 2015, 29, 1376–1385. [Google Scholar] [CrossRef] [PubMed]
- Skovsø, S. Modeling type 2 Diabetes in Rats Using High Fat Diet and Streptozotocin. J. Diabetes Investig. 2014, 5, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, B.; Ziegler, C.G.; Schally, A.V.; Richter, C.; Steffen, A.; Jabs, N.; Funk, R.H.; Brendel, M.D.; Block, N.L.; Ehrhart-Bornstein, M.; et al. Agonist of Growth Hormone-Releasing Hormone as a Potential Effector for Survival and Proliferation of Pancreatic Islets. Proc. Natl Acad. Sci. USA 2010, 107, 12623–12628. [Google Scholar] [CrossRef] [Green Version]
- Xin, F.; Jiang, L.; Liu, X.; Geng, C.; Wang, W.; Zhong, L.; Yang, G.; Chen, M. Bisphenol A Induces Oxidative Stress-Associated DNA Damage in INS-1 Cells. Mutat. Res.-Genet. Toxicol. Environ. Mutagenes. 2014, 769, 29–33. [Google Scholar] [CrossRef]
- Karsten, V.; Sigrist, S.; Moriscot, C.; Benhamou, P.Y.; Lemarchand, P.; Belcourt, A.; Poindron, P.; Pinget, M.; Kessler, L. Reduction of Macrophage Activation After Antioxidant Enzymes Gene Transfer to Rat Insulinoma INS-1 Cells. Immunobiology 2002, 205, 193–203. [Google Scholar] [CrossRef]
- Ježek, P.; Dlasková, A.; Plecitá-Hlavatá, L. Redox Homeostasis in Pancreatic β Cells. Oxidative Med. Cell. Longev. 2012, 2012, 932838. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.A.; Lee, J.H.; Han, J.S. 2,7″-Phloroglucinol-6,6′-bieckol protects INS-1 cells against high glucose-induced apoptosis. Biomed. Pharm. 2018, 103, 1473–1481. [Google Scholar] [CrossRef]
- Lyu, G.M.; Wang, Y.J.; Huang, X.; Zhang, H.Y.; Sun, L.D.; Liu, Y.J.; Yan, C.H. Hydrophilic CeO2 nanocubes protect pancreatic beta-cell line INS-1 from H2O2-induced oxidative stress. Nanoscale 2016, 8, 7923–7932. [Google Scholar] [CrossRef]
- Jurgens, C.A.; Toukatly, M.N.; Fligner, C.L.; Udayasankar, J.; Subramanian, S.L.; Zraika, S.; Aston-Mourney, K.; Carr, D.B.; Westermark, P.; Westermark, G.T.; et al. β-Cell Loss and β-Cell Apoptosis in Human type 2 Diabetes Are Related to Islet Amyloid Deposition. Am. J. Pathol. 2011, 178, 2632–2640. [Google Scholar] [CrossRef] [Green Version]
- Haataja, L.; Gurlo, T.; Huang, C.J.; Butler, P.C. Islet Amyloid in type 2 Diabetes, and the Toxic Oligomer Hypothesis. Endocr. Rev. 2008, 29, 303–316. [Google Scholar] [CrossRef] [Green Version]
- Cao, P.; Marek, P.; Noor, H.; Patsalo, V.; Tu, L.H.; Wang, H.; Abedini, A.; Raleigh, D.P. Islet Amyloid: From Fundamental Biophysics to Mechanisms of Cytotoxicity. FEBS Lett. 2013, 587, 1106–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Tang, Y.; Shi, S.; Gao, S.; Wang, Y.; Xiao, D.; Chen, T.; He, Q.; Zhang, J.; Lin, Y. Tetrahedral Framework Nucleic Acids Ameliorate Insulin Resistance in Type 2 Diabetes Mellitus via the PI3K/Akt Pathway. ACS Appl. Mater. Interfaces 2021, 13, 40354–40364. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, S.; Zhang, W.; Zhao, P.; He, B.; Wu, N.; Han, P. Epigallocatechin-3-O-gallate (EGCG) attenuates FFAs-induced peripheral insulin resistance through AMPK pathway and insulin signaling pathway in vivo. Diabetes Res. Clin. Pract. 2011, 93, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.Y.; Shi, C.X.; Gao, R.; Sun, H.J.; Xiong, X.Q.; Ding, L.; Chen, Q.; Li, Y.H.; Wang, J.J.; Kang, Y.M.; et al. Irisin inhibits hepatic gluconeogenesis and increases glycogen synthesis via the PI3K/Akt pathway in type 2 diabetic mice and hepatocytes. Clin. Sci. 2015, 129, 839–850. [Google Scholar] [CrossRef]
- Hayden, M.R.; Tyagi, S.C. Uric Acid: A New Look at an Old Risk Marker for Cardiovascular Disease, Metabolic Syndrome, and Type 2 Diabetes Mellitus: The Urate Redox Shuttle. Nutr. Metab. 2004, 1, 10–24. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.T.; Yang, M.Y.; Lee, Y.J.; Yang, T.W.; Wang, C.C.; Wang, C.J. Gallic Acid Improves Diabetic Steatosis by Downregulating MicroRNA-34a-5p through Targeting NFE2L2 Expression in High-Fat Diet-Fed db/db Mice. Antioxidants 2021, 11, 92. [Google Scholar] [CrossRef]
- Echouffo-Tcheugui, J.B.; Dieffenbach, S.D.; Kengne, A.P. Added Value of Novel Circulating and Genetic Biomarkers in type 2 Diabetes Prediction: A Systematic Review. Diabetes Res. Clin. Pract. 2013, 101, 255–269. [Google Scholar] [CrossRef]
- Vlachos, D.; Malisova, S.; Lindberg, F.A.; Karaniki, G. Glycemic Index (GI) or Glycemic Load (GL) and Dietary Interventions for Optimizing Postprandial Hyperglycemia in Patients with T2 Diabetes: A Review. Nutrients 2020, 12, 1561. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, W.; Chen, Y.; Zhang, M.; Wang, L.; Zhou, H.; Wu, P.; Teng, X.; Dong, Y.; Zhou, J.W.; et al. Combined Use of Fasting Plasma Glucose and Glycated Hemoglobin A1c in the Screening of Diabetes and Impaired Glucose Tolerance. Acta Diabetol. 2010, 47, 231–236. [Google Scholar] [CrossRef]
- Xiao, Z.; Yu, X.; Zhang, S.; Liang, A. The Expression Levels and Significance of GSH, MDA, SOD, and 8-OHdG in Osteochondral Defects of Rabbit Knee Joints. Biomed. Res. Int. 2022, 2022, 6916179–6916184. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.; Mikail, M.A.; Ahmed, I.A.; Hazali, N.; Abdul Rasad, M.S.B.; Abdul Ghani, R.; Hashim, R.; Arief, S.J.; Md Isa, M.L.; Draman, S. Comparison of the effects of three different Baccaurea angulata whole fruit juice doses on plasma, aorta and liver MDA levels, antioxidant enzymes and total antioxidant capacity. Eur. J. Nutr. 2018, 57, 1817–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Yu, P.; Tong, J.; Sun, W.; Jiang, H.; Wang, Y.; Xue, K.; Xie, F.; Qian, H.; Liu, N.; et al. Hydrogen treatment reduces tendon adhesion and inflammatory response. J. Cell Biochem. 2019, 120, 1610–1619. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Organs Coefficient @ | |||
|---|---|---|---|---|
| Liver | Spleen | Pancreas | Kidney | |
| Control | 3.95 ± 0.45 | 0.39 ± 0.06 | 0.56 ± 0.05 | 1.20 ± 0.26 |
| T2DM Model | 4.98 ± 0.58 * | 0.50 ± 0.26 | 0.58 ± 0.09 | 1.55 ± 0.30 |
| YA 5 mg/kg | 4.75 ± 0.98 | 0.39 ± 0.15 | 0.53 ± 0.19 | 1.48 ± 0.21 |
| YA 10 mg/kg | 5.02 ± 0.51 | 0.41 ± 0.13 | 0.58 ± 0.16 | 1.65 ± 0.17 |
| YA 20 mg/kg | 4.75 ± 0.44 # | 0.39 ± 0.17 | 0.53 ± 0.06 | 1.27 ± 0.61 |
| DMBG 100 mg/kg | 4.36 ± 0.51 # | 0.26 ± 0.07 | 0.61 ± 0.14 | 1.54 ± 0.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, J.; Zhu, Y.; Wu, Y.; Li, N.; Cao, Y.; Guo, Y.; Xu, L. Antioxidant Effect of Tyr-Ala Extracted from Zein on INS-1 Cells and Type 2 Diabetes High-Fat-Diet-Induced Mice. Antioxidants 2022, 11, 1111. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11061111
Zhai J, Zhu Y, Wu Y, Li N, Cao Y, Guo Y, Xu L. Antioxidant Effect of Tyr-Ala Extracted from Zein on INS-1 Cells and Type 2 Diabetes High-Fat-Diet-Induced Mice. Antioxidants. 2022; 11(6):1111. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11061111
Chicago/Turabian StyleZhai, Jinghui, Yuhua Zhu, Yi Wu, Na Li, Yue Cao, Yi Guo, and Li Xu. 2022. "Antioxidant Effect of Tyr-Ala Extracted from Zein on INS-1 Cells and Type 2 Diabetes High-Fat-Diet-Induced Mice" Antioxidants 11, no. 6: 1111. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11061111