
The Protective Anticancer Effect of Natural Lycopene Supercritical CO2 Watermelon Extracts in Adenocarcinoma Lung Cancer Cells

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Oleoresin Extraction and Encapsulation
2.2. Cell Culture of A549 CCL-185™
2.3. Cell Cycle
2.4. Determination of Cell Death and Apoptosis
2.5. Analysis of Intracellular Reactive Oxygen Species (ROS)
2.6. Evaluation of Mitochondrial Stress
2.7. Clonogenic Assay
2.8. Western Blot
2.9. Real Time PCR
2.10. Interleukin-8 (IL-8) Detection Assessment
2.11. Statistical Analysis
3. Results
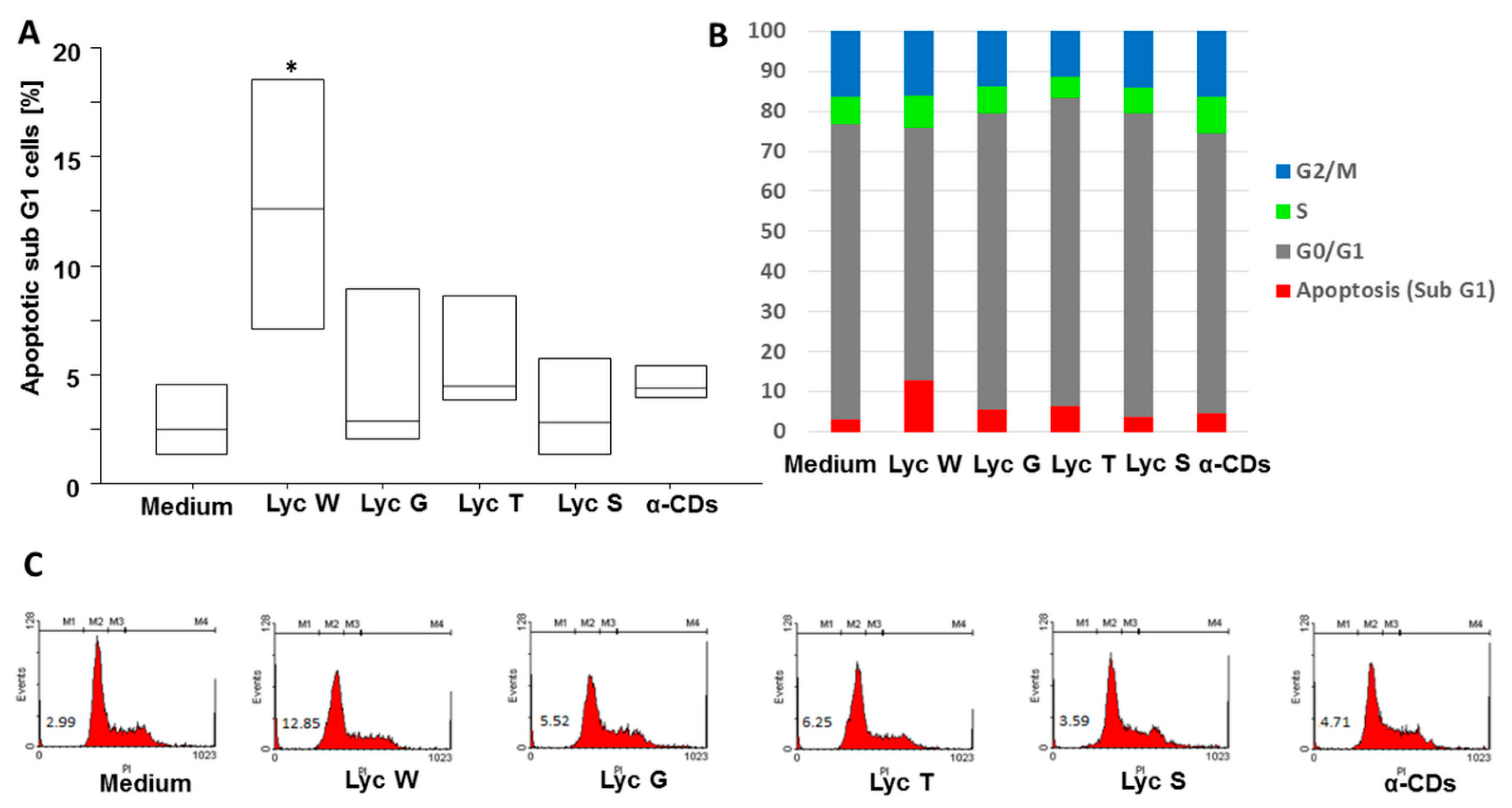
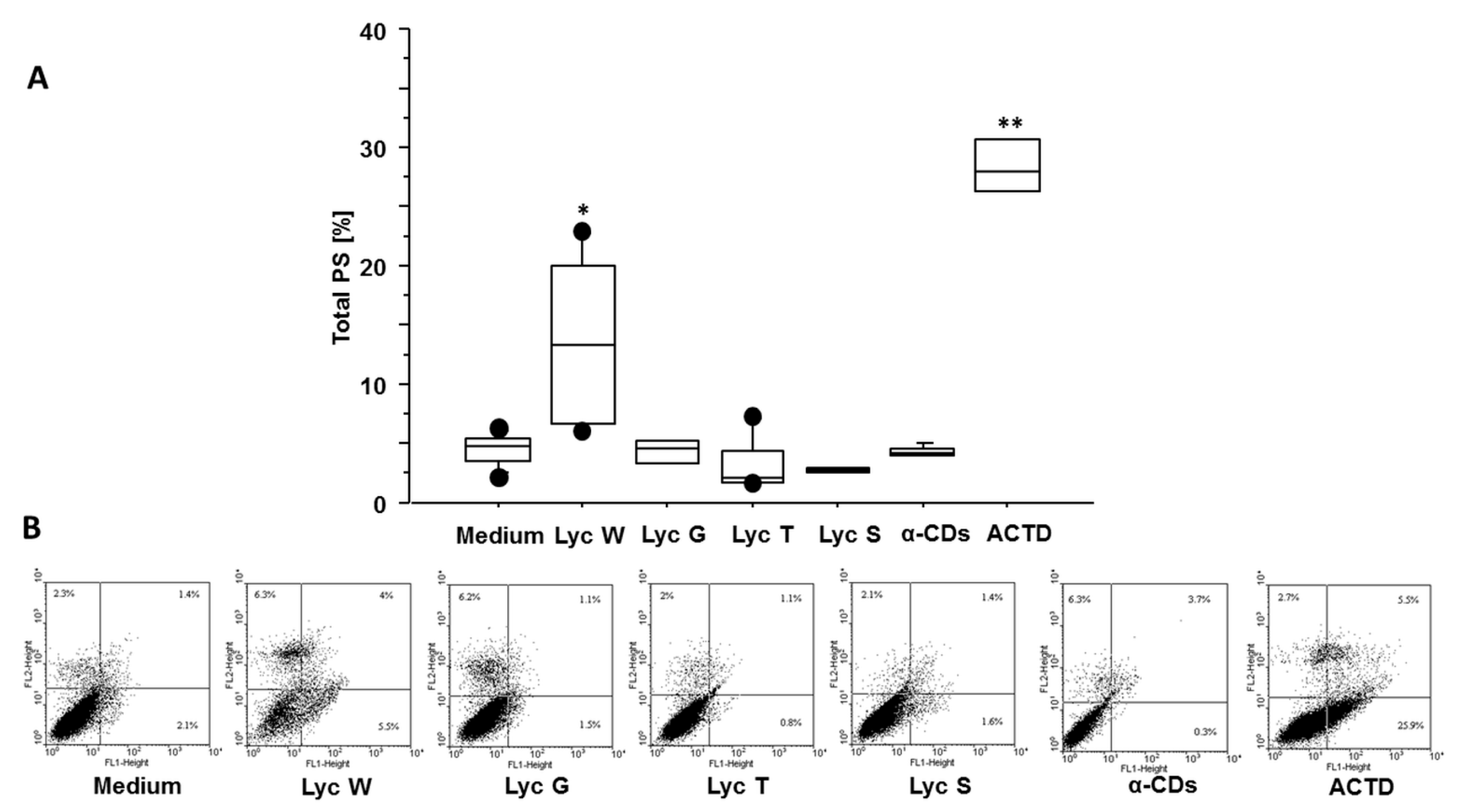
3.1. Lyc W Induces Cell Apoptosis
3.2. Effect of All Three of the α-CD Encapsulated Oleoresins on Colony Formation Ability in A549 Cell Line
3.3. Lyc W Effect on Mitochondrial Stress in A549 Cell Line
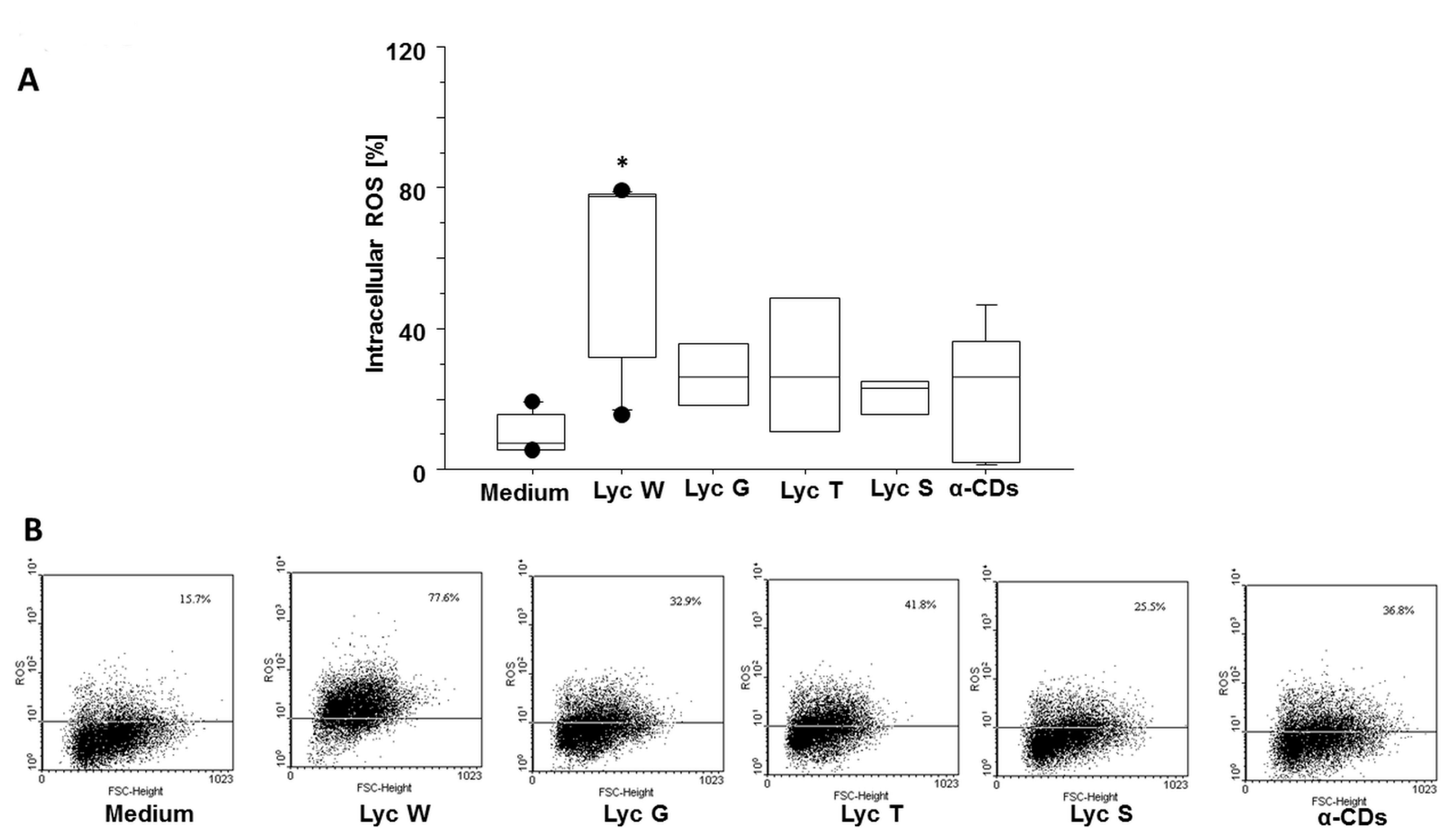
3.4. Lyc W Effect on Spontaneous ROS Generation
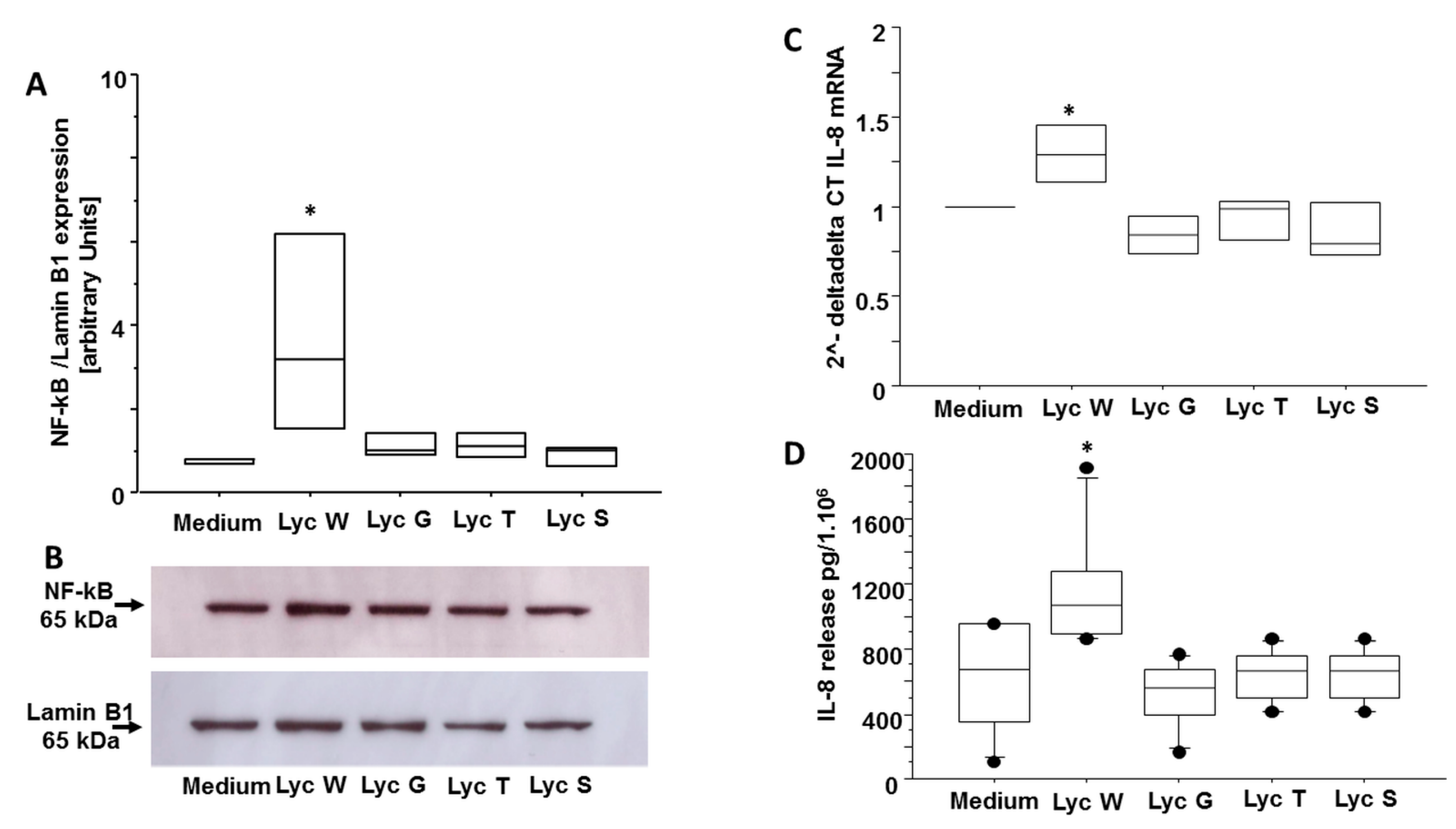
3.5. Lyc W Acts on A549 Survival via Enhanced of the Transcription Factor NF-kB and of Pro-Inflammatory Cytokine IL-8
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meléndez-Martínez, A.J.; Böhm, V.; Borge, G.I.A.; Cano, M.P.; Fikselová, M.; Gruskiene, R.; Lavelli, V.; Loizzo, M.R.; Mandić, A.I.; Brahm, P.M.; et al. Carotenoids: Considerations for Their Use in Functional Foods, Nutraceuticals, Nutricosmetics, Supplements, Botanicals, and Novel Foods in the Context of Sustainability, Circular Economy, and Climate Change. Annu. Rev. Food Sci. Technol. 2021, 12, 433–460. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawata, A.; Murakami, Y.; Suzuki, S.; Fujisawa, S. Anti-Inflammatory Activity of β-Carotene, Lycopene and Tri-n-Butylborane, a Scavenger of Reactive Oxygen Species. Vivo Athens Greece 2018, 32, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Cvetkovic, D.; Fiedor, L.; Fiedor, J.; Wiśniewska-Becker, A.; Markovic, D. Molecular Base for Carotenoids Antioxidant Activity in Model and Biological Systems: The Health-Related Effects. In Carotenoids: Food Sources, Production and Health Benefits; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2013; pp. 93–126. ISBN 978-1-62808-622-5. [Google Scholar]
- Parker, R.S. Carotenoids in Human Blood and Tissues. J. Nutr. 1989, 119, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Mackinney, G. Carotenoids. Annu. Rev. Biochem. 1952, 21, 473–492. [Google Scholar] [CrossRef]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L.; Valanis, B.; Williams, J.H.; et al. Effects of a Combination of Beta Carotene and Vitamin A on Lung Cancer and Cardiovascular Disease. N. Engl. J. Med. 1996, 334, 1150–1155. [Google Scholar] [CrossRef] [Green Version]
- Alpha-Tocopherol Beta Carotene Cancer Prevention Study Group. The Effect of Vitamin E and Beta Carotene on the Incidence of Lung Cancer and Other Cancers in Male Smokers. N. Engl. J. Med. 1994, 330, 1029–1035. [Google Scholar] [CrossRef]
- Russo, G.L.; Moccia, S.; Russo, M.; Spagnuolo, C. Redox Regulation by Carotenoids: Evidence and Conflicts for Their Application in Cancer. Biochem. Pharmacol. 2021, 194, 114838. [Google Scholar] [CrossRef]
- Spychalowicz, A.; Wilk, G.; Śliwa, T.; Ludew, D.; Guzik, T.J. Novel Therapeutic Approaches in Limiting Oxidative Stress and Inflammation. Curr. Pharm. Biotechnol. 2012, 13, 2456–2466. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxid. Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef]
- Cockfield, J.A.; Schafer, Z.T. Antioxidant Defenses: A Context-Specific Vulnerability of Cancer Cells. Cancers 2019, 11, 1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Mechanistic Insight into Oxidative Stress-Triggered Signaling Pathways and Type 2 Diabetes. Molecules 2022, 27, 950. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Kamisah, Y.; Mohamed, N.; Muhammad, N.; Masbah, N.; Fahami, N.A.M.; Mohamed, I.N.; Shuid, A.N.; Saad, Q.M.; Abdullah, A.; et al. Potential Role of Tocotrienols on Non-Communicable Diseases: A Review of Current Evidence. Nutrients 2020, 12, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pace, E.; Di Vincenzo, S.; Di Salvo, E.; Genovese, S.; Dino, P.; Sangiorgi, C.; Ferraro, M.; Gangemi, S. MiR-21 Upregulation Increases IL-8 Expression and Tumorigenesis Program in Airway Epithelial Cells Exposed to Cigarette Smoke. J. Cell. Physiol. 2019, 234, 22183–22194. [Google Scholar] [CrossRef]
- Vu, T.; Jin, L.; Datta, P.K. Effect of Cigarette Smoking on Epithelial to Mesenchymal Transition (EMT) in Lung Cancer. J. Clin. Med. 2016, 5, 44. [Google Scholar] [CrossRef]
- Zheng, M. Classification and Pathology of Lung Cancer. Surg. Oncol. Clin. N. Am. 2016, 25, 447–468. [Google Scholar] [CrossRef]
- Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 5 June 2022).
- Bessone, M.I.D.; Berardi, D.E.; Cirigliano, S.M.; Delbart, D.I.; Peters, M.G.; Todaro, L.B.; Urtreger, A.J. Protein Kinase C Alpha (PKCα) Overexpression Leads to a Better Response to Retinoid Acid Therapy through Retinoic Acid Receptor Beta (RARβ) Activation in Mammary Cancer Cells. J. Cancer Res. Clin. Oncol. 2020, 146, 3241–3253. [Google Scholar] [CrossRef]
- Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pccompound/?term=lycopene (accessed on 5 June 2022).
- Monselise, S.P.; Halevy, A.H. Detection of Lycopene in Pink Orange Fruit. Science 1961, 133, 1478. [Google Scholar] [CrossRef]
- Ilahy, R.; Tlili, I.; Siddiqui, M.W.; Hdider, C.; Lenucci, M.S. Inside and Beyond Color: Comparative Overview of Functional Quality of Tomato and Watermelon Fruits. Front. Plant Sci. 2019, 10, 769. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://www.alliedmarketresearch.com/lycopene-market-A06684 (accessed on 5 June 2022).
- Zhuang, E.; Uchio, E.; Lilly, M.; Zi, X.; Fruehauf, J.P. A Phase II Study of Docetaxel plus Lycopene in Metastatic Castrate Resistant Prostate Cancer. Biomed. Pharmacother. 2021, 143, 112226. [Google Scholar] [CrossRef]
- De Andrades, E.O.; da Costa, J.M.A.R.; de Lima Neto, F.E.M.; de Araujo, A.R.; de Oliveira Silva Ribeiro, F.; Vasconcelos, A.G.; de Jesus Oliveira, A.C.; Sobrinho, J.L.S.; de Almeida, M.P.; Carvalho, A.P.; et al. Acetylated Cashew Gum and Fucan for Incorporation of Lycopene Rich Extract from Red Guava (Psidium guajava L.) in Nanostructured Systems: Antioxidant and Antitumor Capacity. Int. J. Biol. Macromol. 2021, 191, 1026–1037. [Google Scholar] [CrossRef] [PubMed]
- Beynon, R.A.; Richmond, R.C.; Santos Ferreira, D.L.; Ness, A.R.; May, M.; Smith, G.D.; Vincent, E.E.; Adams, C.; Ala-Korpela, M.; Würtz, P.; et al. Investigating the Effects of Lycopene and Green Tea on the Metabolome of Men at Risk of Prostate Cancer: The ProDiet Randomised Controlled Trial. Int. J. Cancer 2019, 144, 1918–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, W.J.; Sheng, W.S.; Peng, C.; Xiaodong, M.; Yao, T.Z. Investigating into Anti-Cancer Potential of Lycopene: Molecular Targets. Biomed. Pharmacother. 2021, 138, 111546. [Google Scholar] [CrossRef]
- Kim, M.; Kim, S.H.; Lim, J.W.; Kim, H. Lycopene Induces Apoptosis by Inhibiting Nuclear Translocation of β-Catenin in Gastric Cancer Cells. J. Physiol. Pharmacol. 2019, 70, 605–611. [Google Scholar] [CrossRef]
- Wang, R.; Lu, X.; Yu, R. Lycopene Inhibits Epithelial-Mesenchymal Transition and Promotes Apoptosis in Oral Cancer via PI3K/AKT/m-TOR Signal Pathway. Drug Des. Dev. Ther. 2020, 14, 2461–2471. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Lim, J.W.; Kim, H. Lycopene Inhibits Reactive Oxygen Species-Mediated NF-ΚB Signaling and Induces Apoptosis in Pancreatic Cancer Cells. Nutrients 2019, 11, 762. [Google Scholar] [CrossRef] [Green Version]
- Bruno, A.; Durante, M.; Marrese, P.P.; Migoni, D.; Laus, M.N.; Pace, E.; Pastore, D.; Mita, G.; Piro, G.; Lenucci, M.S. Shades of Red: Comparative Study on Supercritical CO2 Extraction of Lycopene-Rich Oleoresins from Gac, Tomato and Watermelon Fruits and Effect of the α-Cyclodextrin Clathrated Extracts on Cultured Lung Adenocarcinoma Cells’ Viability. J. Food Compos. Anal. 2018, 65, 23–32. [Google Scholar] [CrossRef]
- Fan, J.-H.; Wang, J.-B.; Yang, H.; Dawsey, S.M.; Taylor, P.R.; Qiao, Y.-L.; Abnet, C.C. Combined Lifestyle Factors and Risk of All-Cause and Cause-Specific Mortality among Participants in the Linxian Nutrition Intervention Trial: A Cohort, Observational Study. Front. Cardiovasc. Med. 2022, 9, 772617. [Google Scholar] [CrossRef]
- Kunzmann, A.T.; Coleman, H.G.; Huang, W.-Y.; Cantwell, M.M.; Kitahara, C.M.; Berndt, S.I. Fruit and Vegetable Intakes and Risk of Colorectal Cancer and Incident and Recurrent Adenomas in the PLCO Cancer Screening Trial. Int. J. Cancer 2016, 138, 1851–1861. [Google Scholar] [CrossRef]
- Abar, L.; Vieira, A.R.; Aune, D.; Stevens, C.; Vingeliene, S.; Navarro Rosenblatt, D.A.; Chan, D.; Greenwood, D.C.; Norat, T. Blood Concentrations of Carotenoids and Retinol and Lung Cancer Risk: An Update of the WCRF-AICR Systematic Review of Published Prospective Studies. Cancer Med. 2016, 5, 2069–2083. [Google Scholar] [CrossRef] [Green Version]
- Lenucci, M.S.; Caccioppola, A.; Durante, M.; Serrone, L.; Leonardo, R.; Piro, G.; Dalessandro, G. Optimisation of Biological and Physical Parameters for Lycopene Supercritical CO2 Extraction from Ordinary and High-Pigment Tomato Cultivars. J. Sci. Food Agric. 2010, 90, 1709–1718. [Google Scholar] [CrossRef] [PubMed]
- Wankar, J.; Kotla, N.G.; Gera, S.; Rasala, S.; Pandit, A.; Rochev, Y.A. Recent Advances in Host–Guest Self-Assembled Cyclodextrin Carriers: Implications for Responsive Drug Delivery and Biomedical Engineering. Adv. Funct. Mater. 2020, 30, 1909049. [Google Scholar] [CrossRef]
- Durante, M.; Lenucci, M.S.; Gazza, L.; Taddei, F.; Nocente, F.; De Benedetto, G.E.; De Caroli, M.; Piro, G.; Mita, G. Bioactive Composition and Sensory Evaluation of Innovative Spaghetti Supplemented with Free or α-Cyclodextrin Chlatrated Pumpkin Oil Extracted by Supercritical CO2. Food Chem. 2019, 294, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Lenucci, M.S.; Marrese, P.P.; Rizzi, V.; De Caroli, M.; Piro, G.; Fini, P.; Russo, G.L.; Mita, G. A-Cyclodextrin Encapsulation of Supercritical CO2 Extracted Oleoresins from Different Plant Matrices: A Stability Study. Food Chem. 2016, 199, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Milano, F.; Caroli, M.D.; Giotta, L.; Piro, G.; Mita, G.; Frigione, M.; Lenucci, M.S. Tomato Oil Encapsulation by α-, β-, and γ-Cyclodextrins: A Comparative Study on the Formation of Supramolecular Structures, Antioxidant Activity, and Carotenoid Stability. Foods 2020, 9, 1553. [Google Scholar] [CrossRef] [PubMed]
- Sano, C.D.; D’Anna, C.; Scurria, A.; Lino, C.; Pagliaro, M.; Ciriminna, R.; Pace, E. Mesoporous Silica Particles Functionalized with Newly Extracted Fish Oil (Omeg@Silica) Inhibit Lung Cancer Cell Growth. Nanomedicine 2021, 16, 2061–2074. [Google Scholar] [CrossRef]
- Bruno, A.; Siena, L.; Gerbino, S.; Ferraro, M.; Chanez, P.; Giammanco, M.; Gjomarkaj, M.; Pace, E. Apigenin Affects Leptin/Leptin Receptor Pathway and Induces Cell Apoptosis in Lung Adenocarcinoma Cell Line. Eur. J. Cancer 2011, 47, 2042–2051. [Google Scholar] [CrossRef]
- Ferraro, M.; Gjomarkaj, M.; Siena, L.; Di Vincenzo, S.; Pace, E. Formoterol and Fluticasone Propionate Combination Improves Histone Deacetylation and Anti-Inflammatory Activities in Bronchial Epithelial Cells Exposed to Cigarette Smoke. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1718–1727. [Google Scholar] [CrossRef]
- Costantini, F.; Di Sano, C.; Barbieri, G. The Hydroxytyrosol Induces the Death for Apoptosis of Human Melanoma Cells. Int. J. Mol. Sci. 2020, 21, 8074. [Google Scholar] [CrossRef]
- Brzozowska, B.; Gałecki, M.; Tartas, A.; Ginter, J.; Kaźmierczak, U.; Lundholm, L. Freeware Tool for Analysing Numbers and Sizes of Cell Colonies. Radiat. Environ. Biophys. 2019, 58, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Haslett, J.N.; Sanoudou, D.; Kho, A.T.; Bennett, R.R.; Greenberg, S.A.; Kohane, I.S.; Beggs, A.H.; Kunkel, L.M. Gene Expression Comparison of Biopsies from Duchenne Muscular Dystrophy (DMD) and Normal Skeletal Muscle. Proc. Natl. Acad. Sci. USA 2002, 99, 15000–15005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-H.; Lim, S.-N.; Chen, C.-Y.; Chi, H.-C.; Yeh, C.-T.; Lin, W.-R. Functional Role of Mitochondrial DNA in Cancer Progression. Int. J. Mol. Sci. 2022, 23, 1659. [Google Scholar] [CrossRef] [PubMed]
- Arathi, B.P.; Sowmya, P.R.-R.; Kuriakose, G.C.; Vijay, K.; Baskaran, V.; Jayabaskaran, C.; Lakshminarayana, R. Enhanced Cytotoxic and Apoptosis Inducing Activity of Lycopene Oxidation Products in Different Cancer Cell Lines. Food Chem. Toxicol. 2016, 97, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Iacobini, C.; Vitale, M.; Haxhi, J.; Pesce, C.; Pugliese, G.; Menini, S. Food-Related Carbonyl Stress in Cardiometabolic and Cancer Risk Linked to Unhealthy Modern Diet. Nutrients 2022, 14, 1061. [Google Scholar] [CrossRef]
- Mathuram, T.L.; Ravikumar, V.; Reece, L.M.; Karthik, S.; Sasikumar, C.S.; Cherian, K.M. Tideglusib Induces Apoptosis in Human Neuroblastoma IMR32 Cells, Provoking Sub-G0/G1 Accumulation and ROS Generation. Environ. Toxicol. Pharmacol. 2016, 46, 194–205. [Google Scholar] [CrossRef]
- Mehdad, A.; Brumana, G.; Souza, A.A.; Barbosa, J.; Ventura, M.M.; de Freitas, S.M. A Bowman-Birk Inhibitor Induces Apoptosis in Human Breast Adenocarcinoma through Mitochondrial Impairment and Oxidative Damage Following Proteasome 20S Inhibition. Cell Death Discov. 2016, 2, 15067. [Google Scholar] [CrossRef] [Green Version]
- Nazemiyeh, E.; Eskandani, M.; Sheikhloie, H.; Nazemiyeh, H. Formulation and Physicochemical Characterization of Lycopene-Loaded Solid Lipid Nanoparticles. Adv. Pharm. Bull. 2016, 6, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Sharma, G.; Ghoshal, G.; Kesharwani, P.; Singh, B.; Shivhare, U.S.; Katare, O.P. Lycopene Loaded Whey Protein Isolate Nanoparticles: An Innovative Endeavor for Enhanced Bioavailability of Lycopene and Anti-Cancer Activity. Int. J. Pharm. 2018, 546, 97–105. [Google Scholar] [CrossRef]
- Lenucci, M.S.; De Caroli, M.; Marrese, P.P.; Iurlaro, A.; Rescio, L.; Böhm, V.; Dalessandro, G.; Piro, G. Enzyme-Aided Extraction of Lycopene from High-Pigment Tomato Cultivars by Supercritical Carbon Dioxide. Food Chem. 2015, 170, 193–202. [Google Scholar] [CrossRef]
- Honda, M.; Higashiura, T.; Fukaya, T. Safety Assessment of a Natural Tomato Oleoresin Containing High Amounts of Z-Isomers of Lycopene Prepared with Supercritical Carbon Dioxide. J. Sci. Food Agric. 2017, 97, 1027–1033. [Google Scholar] [CrossRef]
- Violi, F.; Nocella, C.; Loffredo, L.; Carnevale, R.; Pignatelli, P. Interventional Study with Vitamin E in Cardiovascular Disease and Meta-Analysis. Free Radic. Biol. Med. 2022, 178, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Klein, E.A.; Thompson, I.M.J.; Tangen, C.M.; Crowley, J.J.; Lucia, M.S.; Goodman, P.J.; Minasian, L.M.; Ford, L.G.; Parnes, H.L.; Gaziano, J.M.; et al. Vitamin E and the Risk of Prostate Cancer: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA 2011, 306, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Cuzick, J. Preventive Therapy for Cancer. Lancet Oncol. 2017, 18, e472–e482. [Google Scholar] [CrossRef]
- Goldkorn, T.; Filosto, S.; Chung, S. Lung Injury and Lung Cancer Caused by Cigarette Smoke-Induced Oxidative Stress: Molecular Mechanisms and Therapeutic Opportunities Involving the Ceramide-Generating Machinery and Epidermal Growth Factor Receptor. Antioxid. Redox Signal. 2014, 21, 2149–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; LLeonart, M.E. Oxidative Stress and Cancer: An Overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Zingone, A.; Yu, Y.; Zhu, B.; Candia, J.; Cao, L.; Ryan, B.M. Relationship between Circulating Inflammation Proteins and Lung Cancer Diagnosis in the National Lung Screening Trial. Cancer Epidemiol. Prev. Biomark. 2019, 28, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Hoesel, B.; Schmid, J.A. The Complexity of NF-ΚB Signaling in Inflammation and Cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Montero, P.; Milara, J.; Pérez-Leal, M.; Estornut, C.; Roger, I.; Pérez-Fidalgo, A.; Sanz, C.; Cortijo, J. Paclitaxel-Induced Epidermal Alterations: An In Vitro Preclinical Assessment in Primary Keratinocytes and in a 3D Epidermis Model. Int. J. Mol. Sci. 2022, 23, 1142. [Google Scholar] [CrossRef]
- Lee, A.J.; Lim, J.W.; Kim, H. Ascorbic Acid Suppresses House Dust Mite-Induced Expression of Interleukin-8 in Human Respiratory Epithelial Cells. J. Cancer Prev. 2021, 26, 64–70. [Google Scholar] [CrossRef]
- Scuto, M.; Ontario, M.L.; Salinaro, A.T.; Caligiuri, I.; Rampulla, F.; Zimbone, V.; Modafferi, S.; Rizzolio, F.; Canzonieri, V.; Calabrese, E.J.; et al. Redox Modulation by Plant Polyphenols Targeting Vitagenes for Chemoprevention and Therapy: Relevance to Novel Anti-Cancer Interventions and Mini-Brain Organoid Technology. Free Radic. Biol. Med. 2022, 179, 59–75. [Google Scholar] [CrossRef]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Wang, H.; et al. Role of the NFκB-Signaling Pathway in Cancer. OncoTargets Ther. 2018, 11, 2063–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinatizadeh, M.R.; Schock, B.; Chalbatani, G.M.; Zarandi, P.K.; Jalali, S.A.; Miri, S.R. The Nuclear Factor Kappa B (NF-KB) Signaling in Cancer Development and Immune Diseases. Genes Dis. 2021, 8, 287–297. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lyc W | Lyc G | Lyc T | |
|---|---|---|---|
| Total carotenoids (μg/mL suspension) | 504.5 ± 5.5 | 89.0 ± 5.4 | 266.0 ± 17.1 |
| All-trans-lycopene | 215.8 ± 1.1 | 49.1 ± 2.1 | 152.7 ± 9.2 |
| lycopene cis isomers | 20.2 ± 0.9 | 7.7 ± 1.2 | 33.6 ± 1.7 |
| α-carotene | 27.0 ± 2.0 | 2.3 ± 0.2 | 16.0 ± 2.7 |
| β-carotene | 241.5 ± 1.5 | 29.9 ± 1.9 | 63.7 ± 3.5 |
| Total tocochromanols (μg/mL suspension) | 22.4 ± 1.8 | 8.6 ± 0.9 | 498.5 ± 16.9 |
| α-tocopherol | nd | 8.6 ± 0.9 | 425.7 ± 12.3 |
| γ-tocopherol | 8.9 ± 0.9 | nd | 72.8 ± 4.6 |
| γ-tocotrienol | 13.5 ± 0.9 | nd | nd |
| Total lycopene/Total tocochromanols | 10.53 | 6.6 | 0.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Sano, C.; Lazzara, V.; Durante, M.; D’Anna, C.; Bonura, A.; Dino, P.; Uasuf, C.G.; Pace, E.; Lenucci, M.S.; Bruno, A. The Protective Anticancer Effect of Natural Lycopene Supercritical CO2 Watermelon Extracts in Adenocarcinoma Lung Cancer Cells. Antioxidants 2022, 11, 1150. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11061150
Di Sano C, Lazzara V, Durante M, D’Anna C, Bonura A, Dino P, Uasuf CG, Pace E, Lenucci MS, Bruno A. The Protective Anticancer Effect of Natural Lycopene Supercritical CO2 Watermelon Extracts in Adenocarcinoma Lung Cancer Cells. Antioxidants. 2022; 11(6):1150. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11061150
Chicago/Turabian StyleDi Sano, Caterina, Valentina Lazzara, Miriana Durante, Claudia D’Anna, Angela Bonura, Paola Dino, Carina Gabriela Uasuf, Elisabetta Pace, Marcello Salvatore Lenucci, and Andreina Bruno. 2022. "The Protective Anticancer Effect of Natural Lycopene Supercritical CO2 Watermelon Extracts in Adenocarcinoma Lung Cancer Cells" Antioxidants 11, no. 6: 1150. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11061150