
Comprehensive Expression Analyses of Plastidial Thioredoxins of Arabidopsis thaliana Indicate a Main Role of Thioredoxin m2 in Roots


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Arabidopsis Plant Growth Conditions
2.2. Protein Extraction and Western Blot Analysis
2.3. GFP Fusion Constructs
2.4. Confocal Microscopy Observation of GFP Fluorescence and Propidium Iodide Staining
2.5. Scanning Electron Microscopy
2.6. Stomatal Conductance and Transpiration Determinations
2.7. In Vitro FBPase Activation Assays
2.8. Statistical Analyses
3. Results
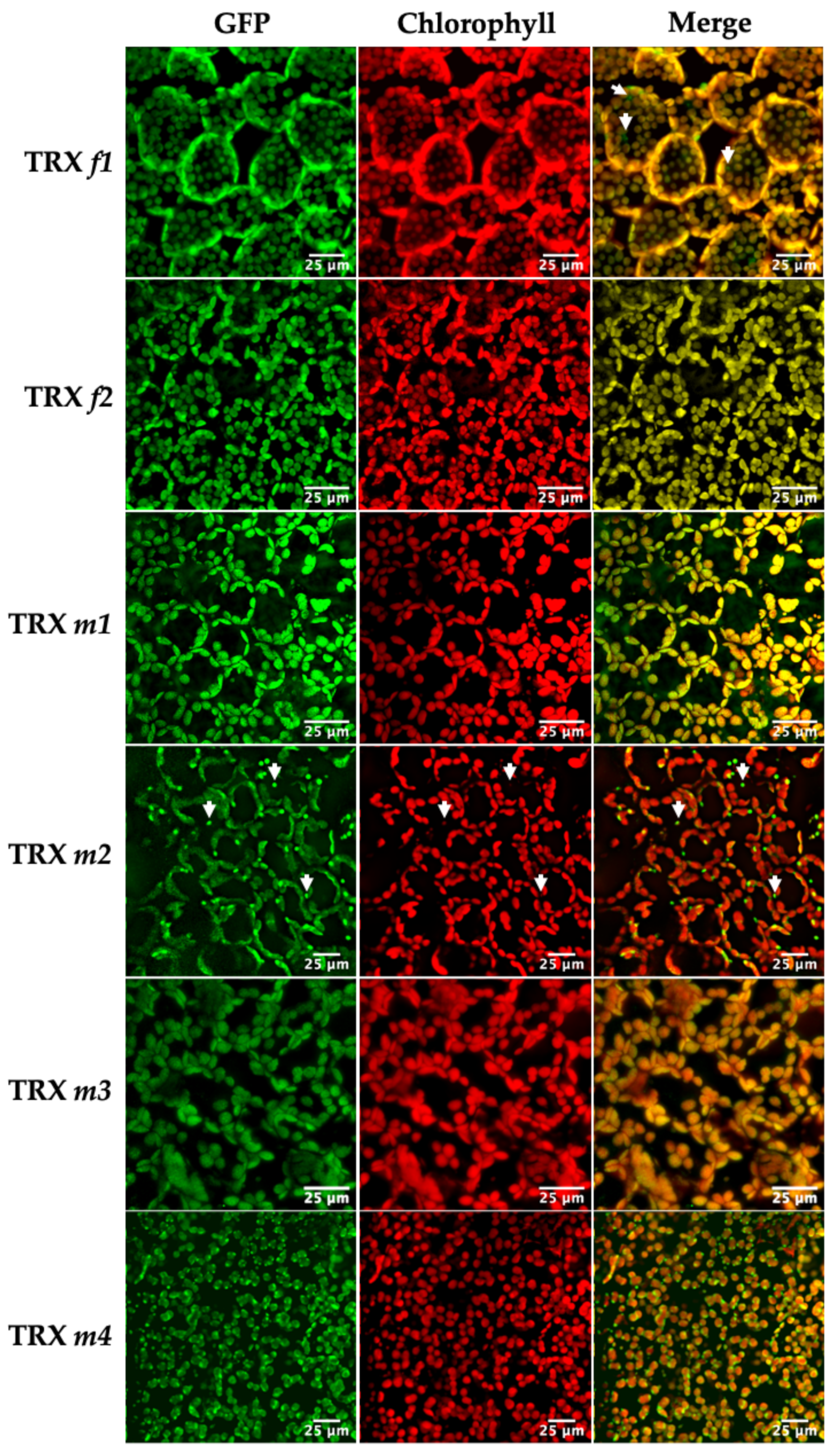
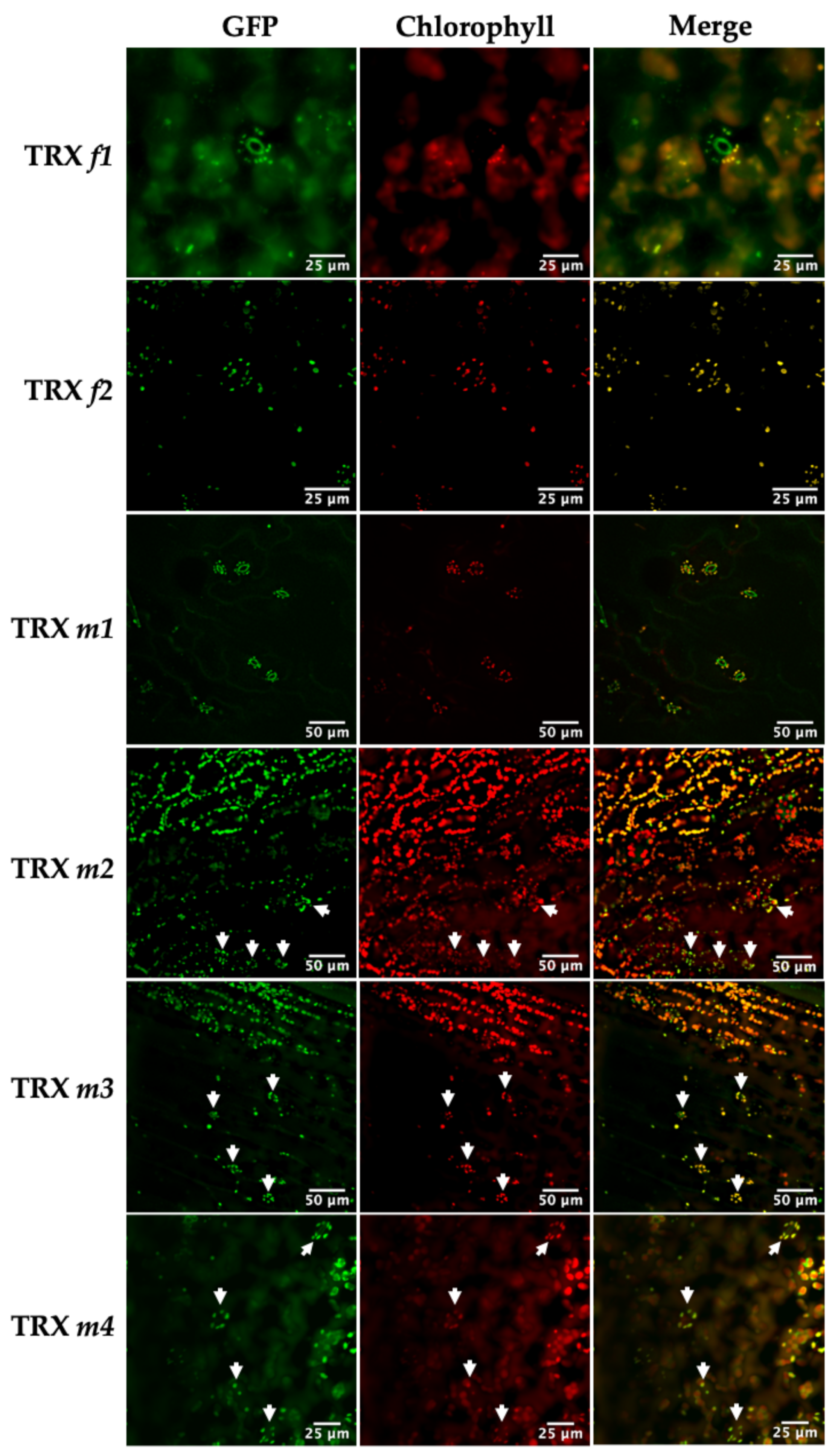
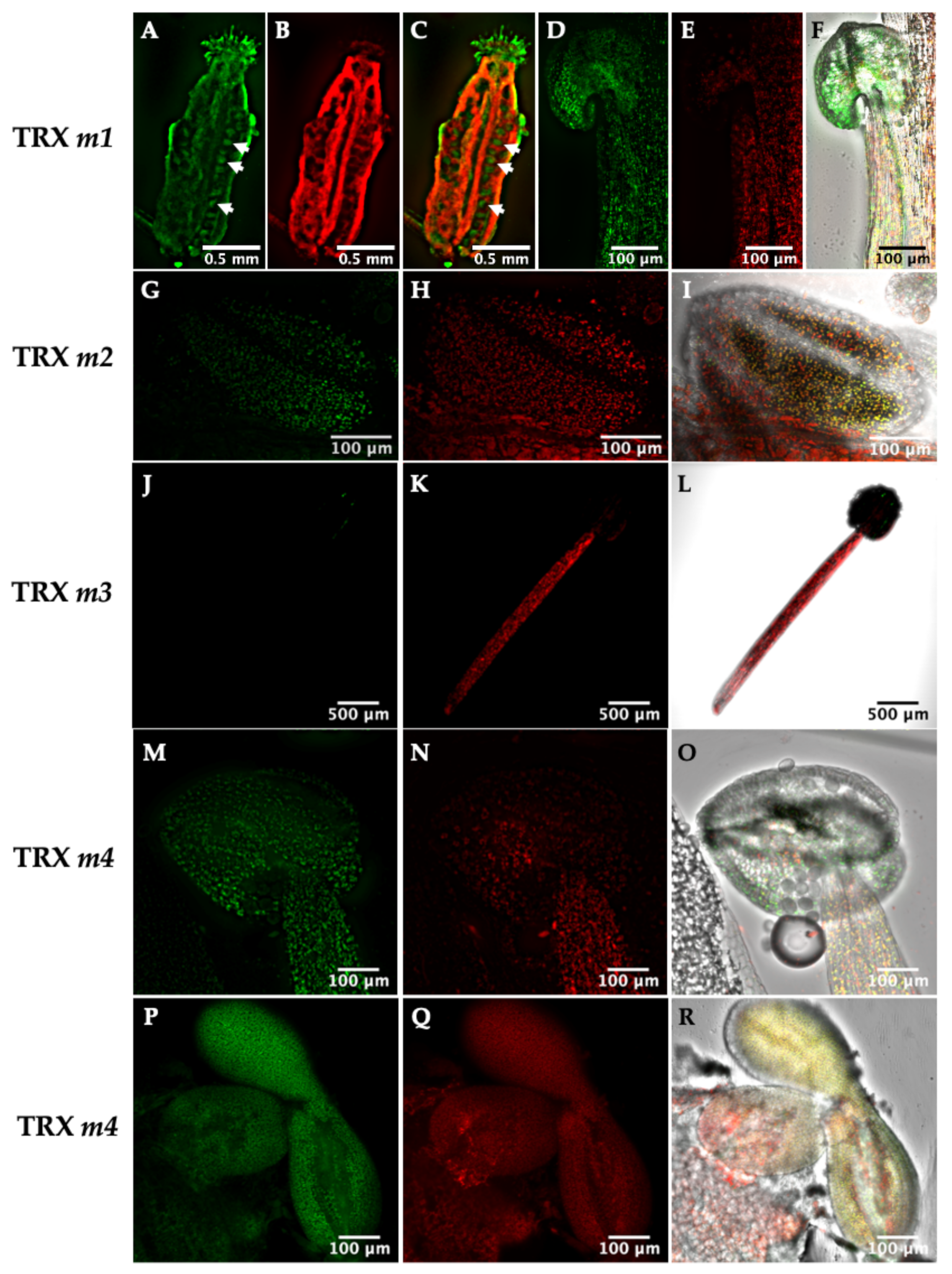
3.1. Location of TRXs f and m in Photosynthetic and Heterotrophic Tissues
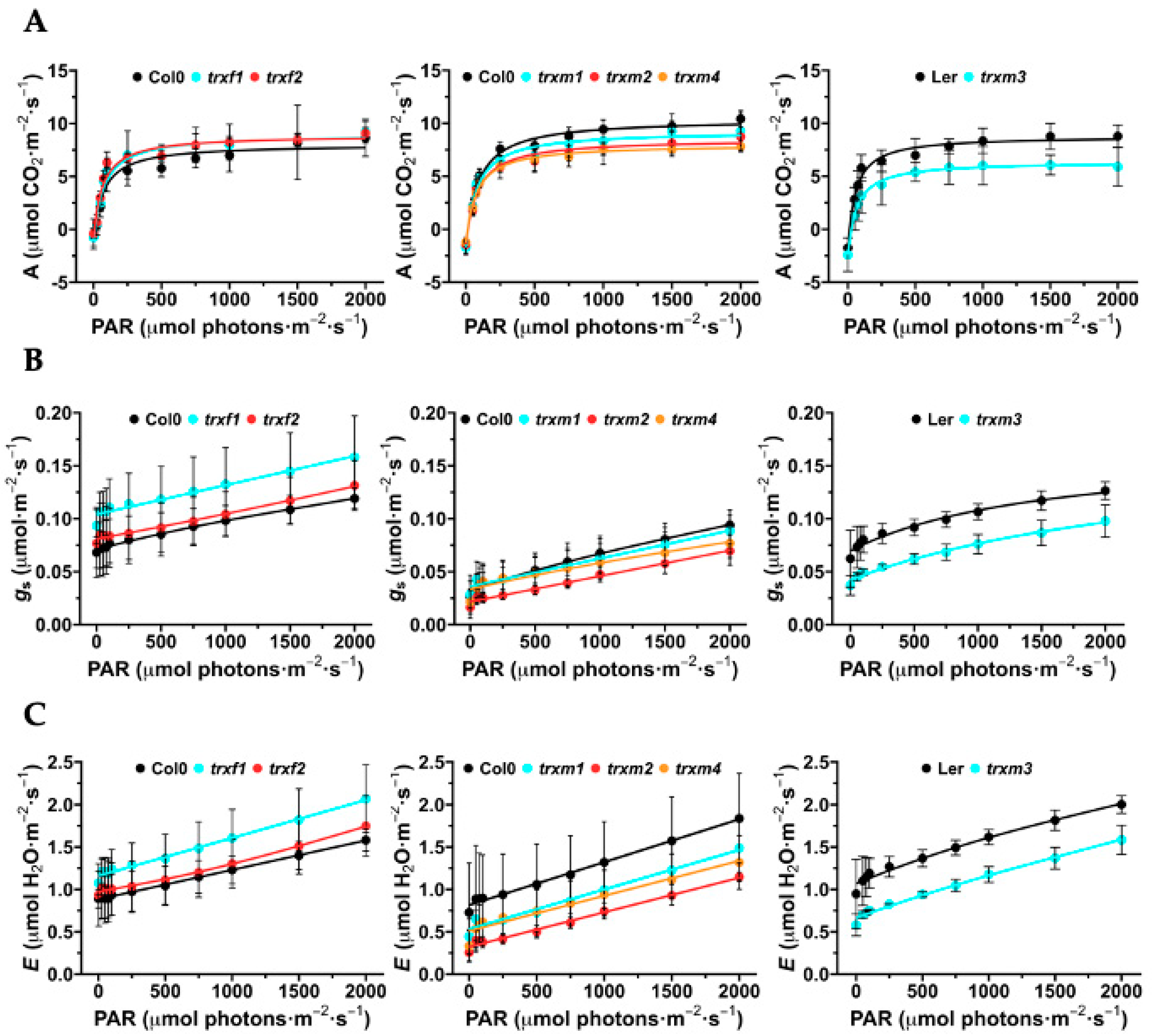
3.2. Photosynthesis, Stomatal Conductance and Transpiration of the TRX Mutants
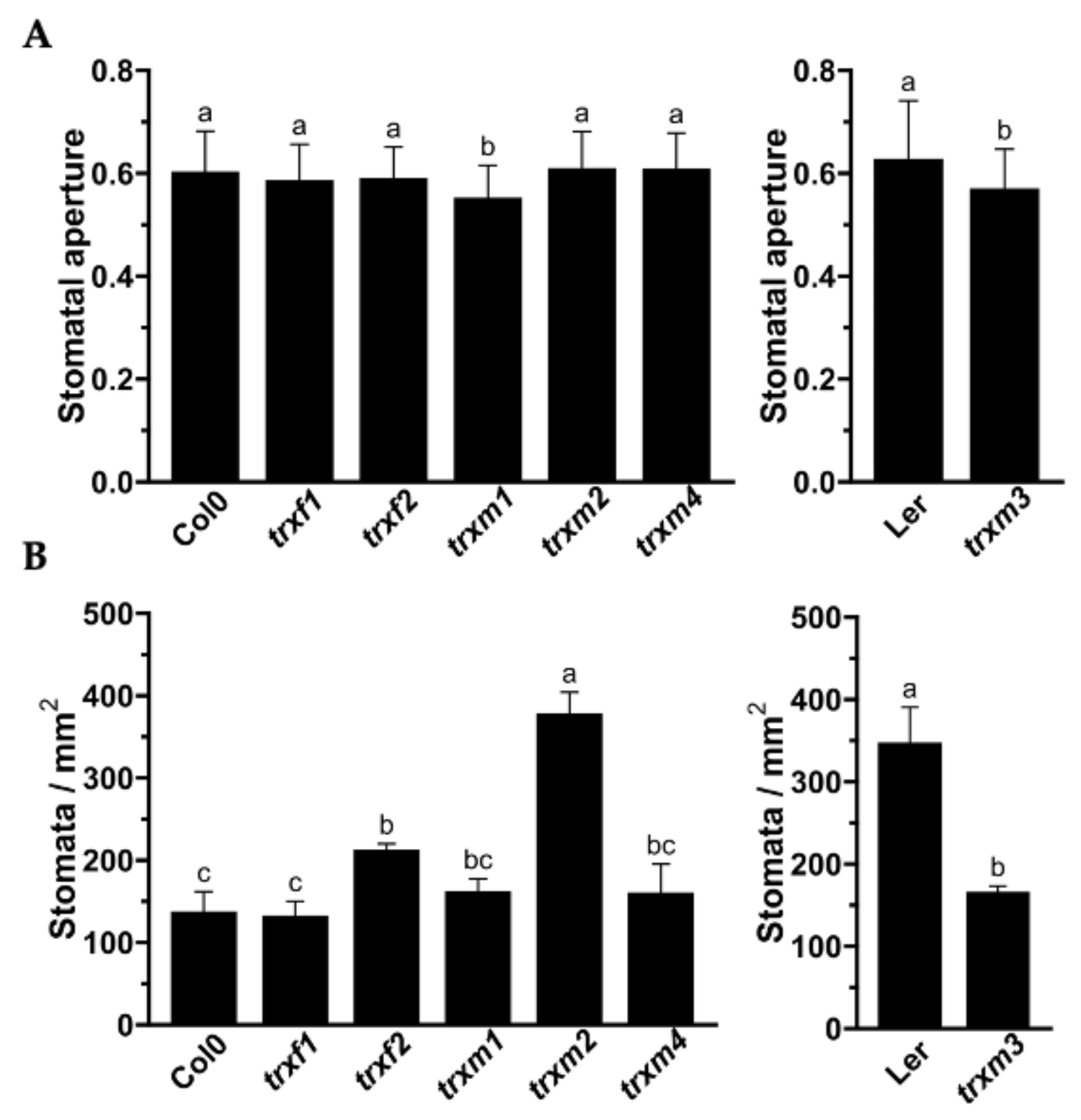
3.3. Stomata Number and Aperture of the TRXs Mutants
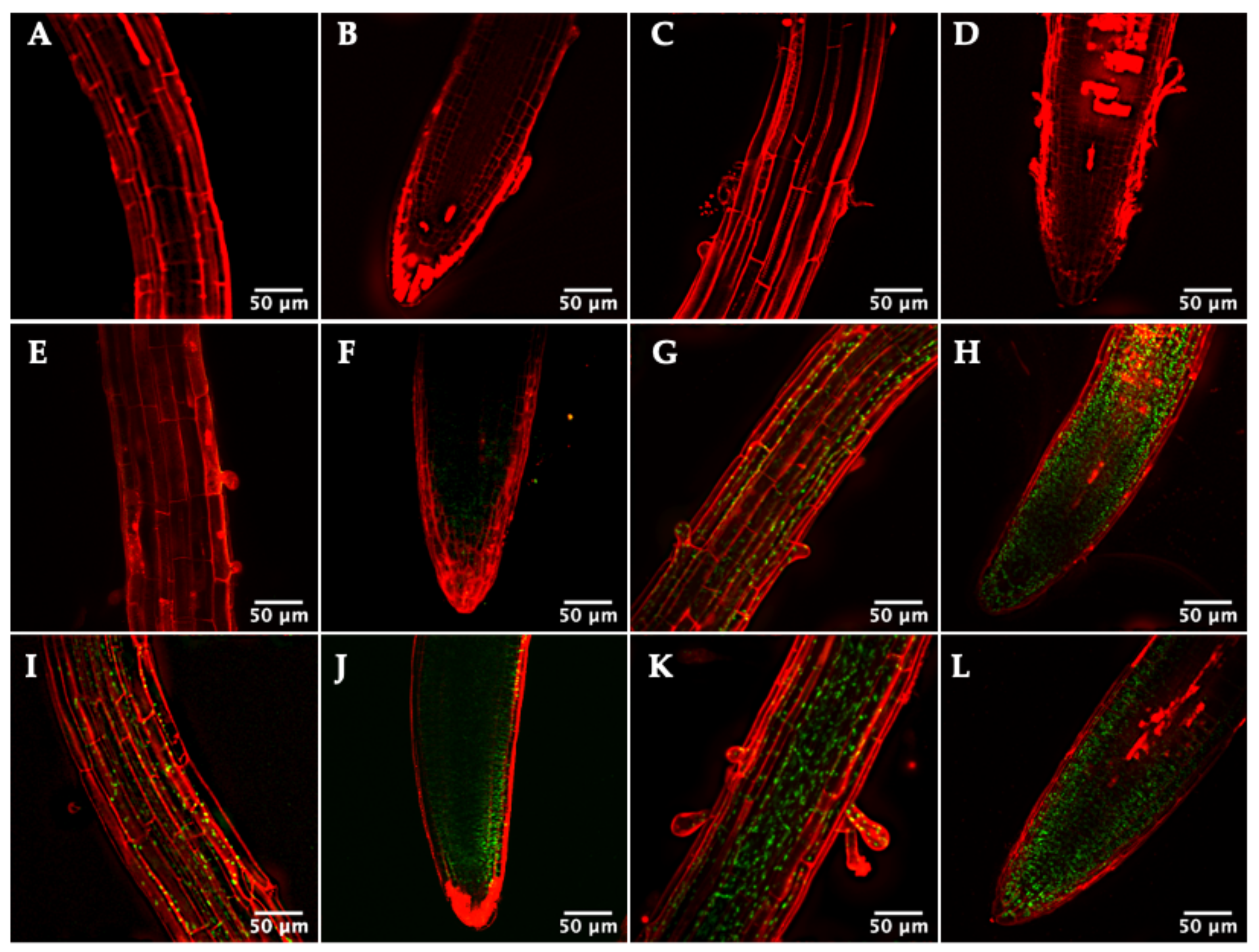
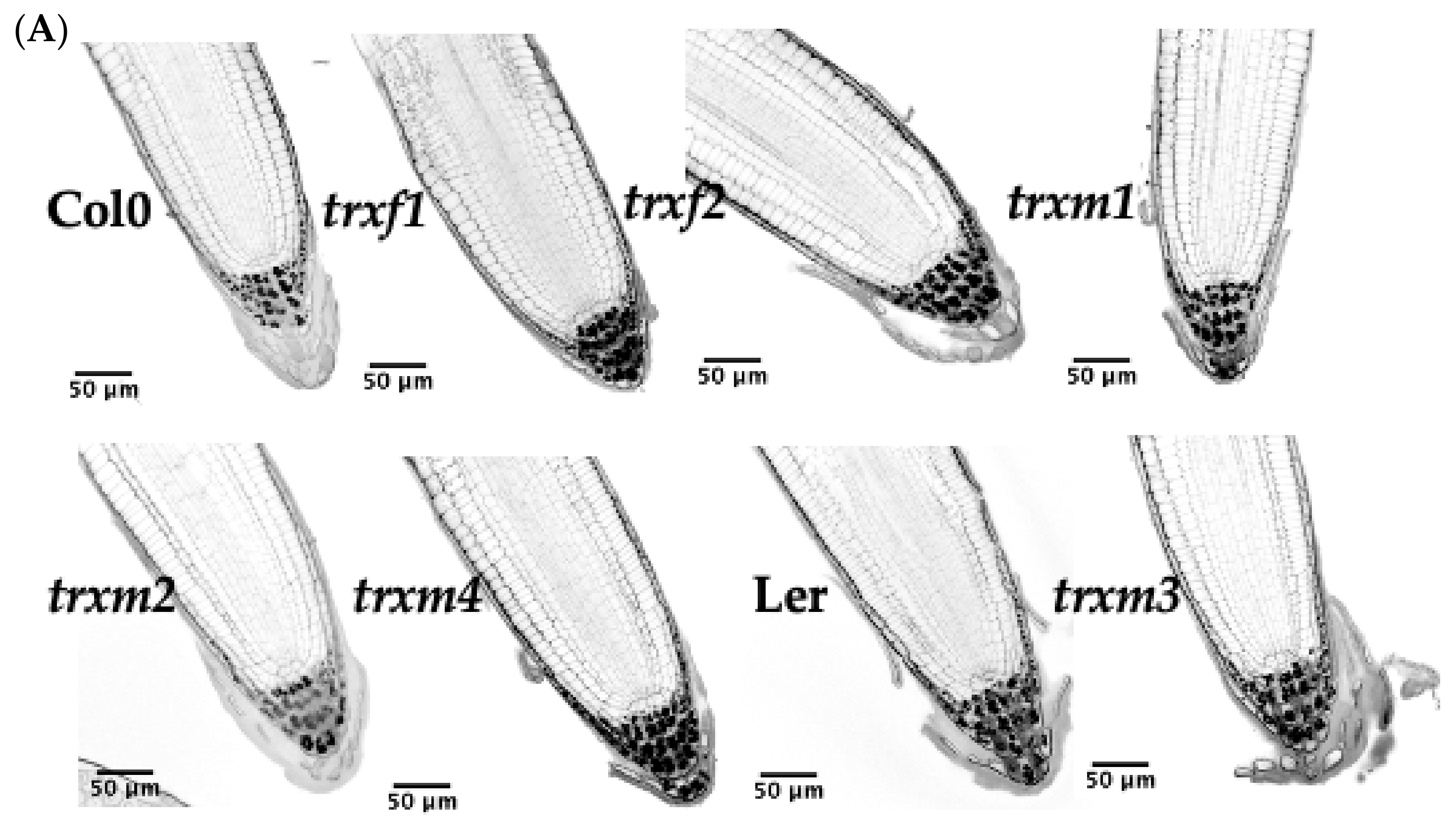
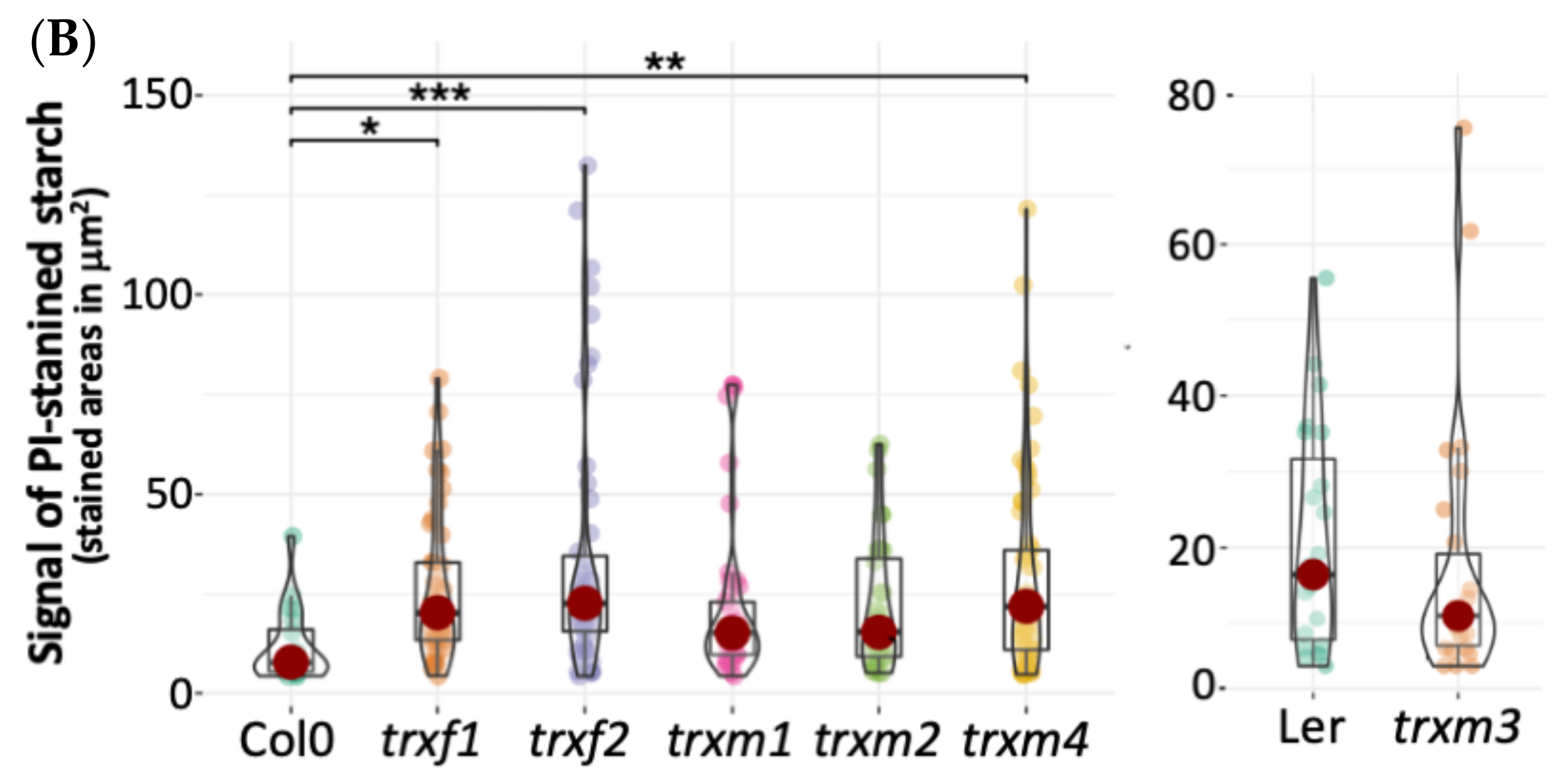
3.4. Primary Root Structure of the TRX Mutants
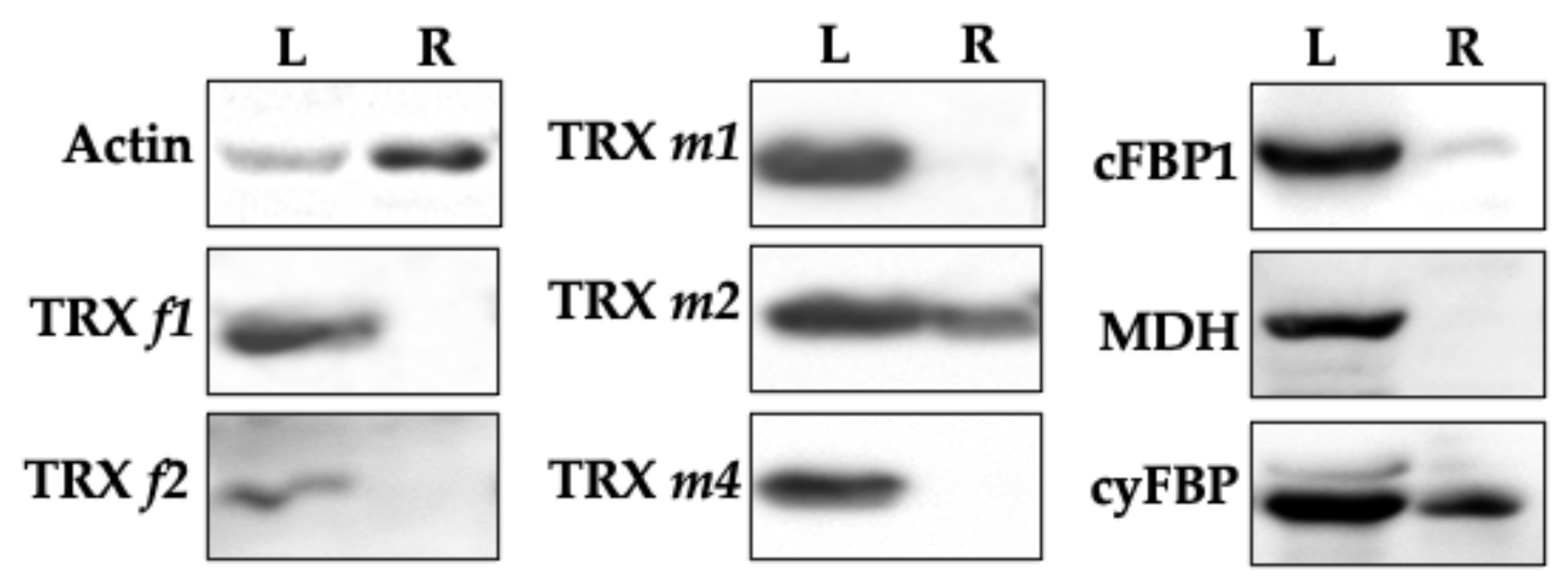
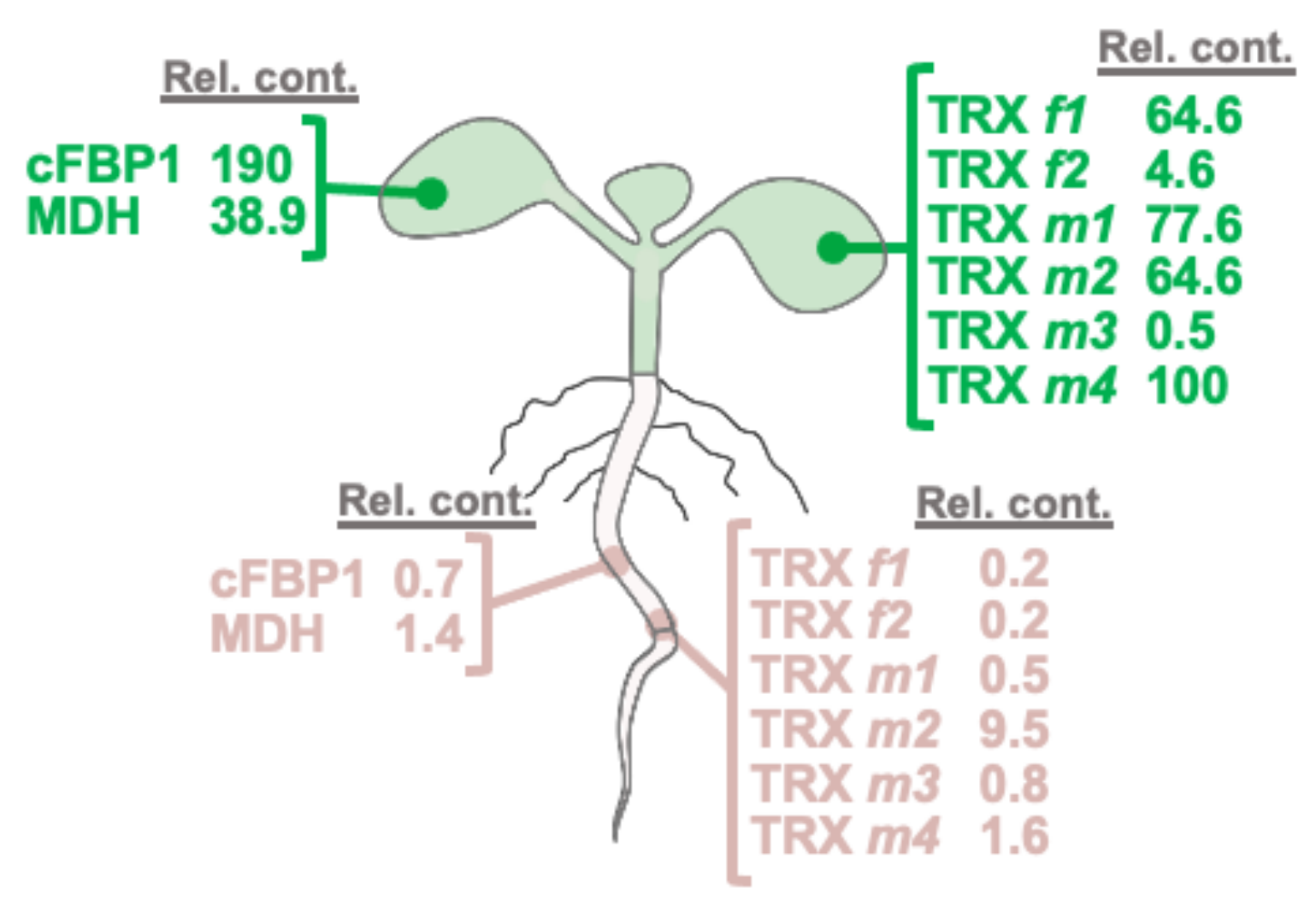
3.5. Immunodetection of Plastid TRXs and Target Proteins in Leaves and Roots
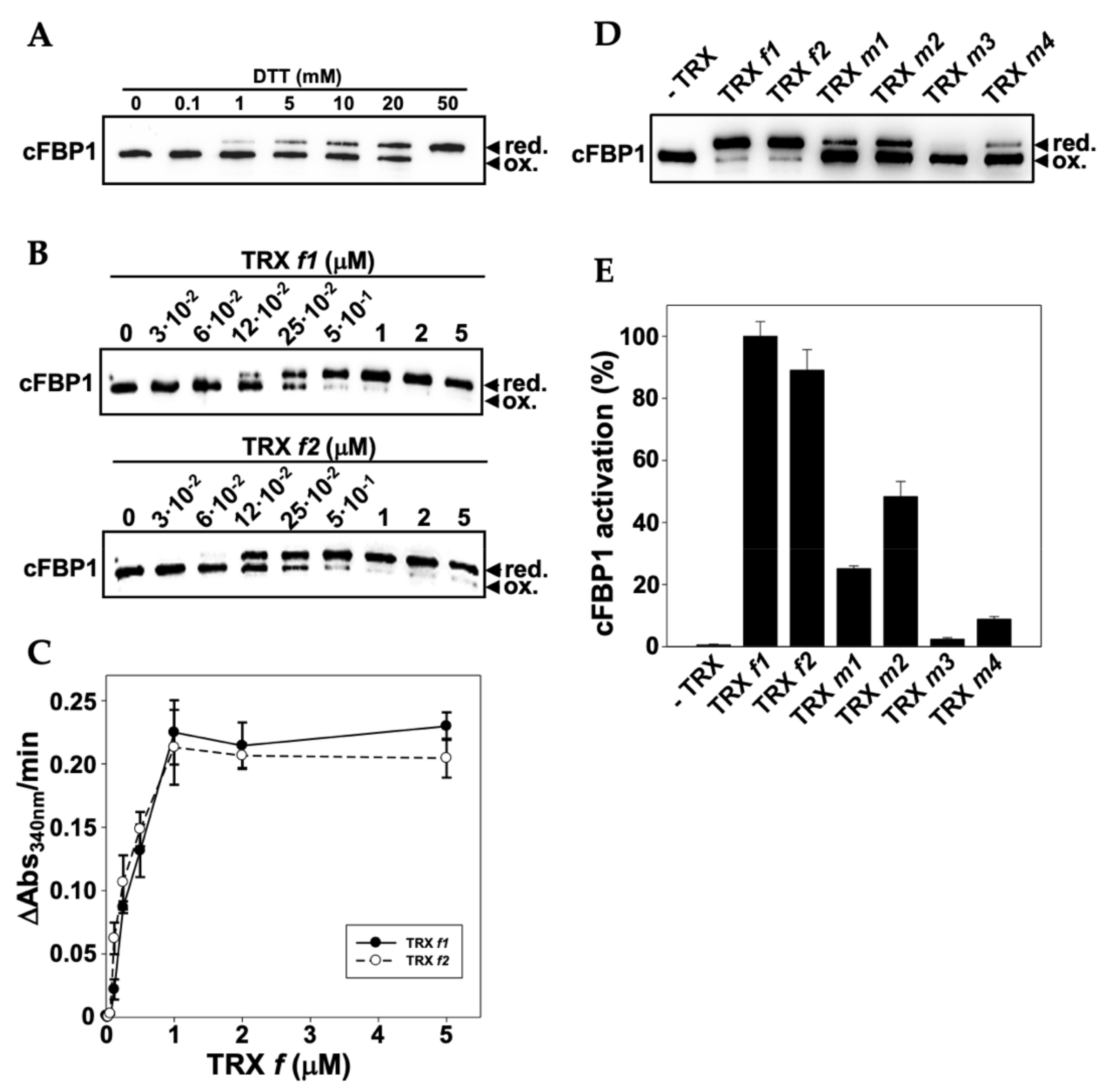
3.6. Functional Specificity of Arabidopsis TRXs f and m in the Redox Activation of FBPase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buchanan, B.B.; Schürmann, P.; Wolosiuk, R.A.; Jacquot, J.P. The ferredoxin/thioredoxin system: From discovery to molecular structures and beyond. Photosynth. Res. 2002, 73, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Montrichard, F.; Alkhalfioui, F.; Yano, H.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin targets in plants: The first 30 years. J. Proteom. 2009, 72, 452–474. [Google Scholar] [CrossRef] [PubMed]
- Gütle, D.D.; Roret, T.; Müller, S.J.; Couturier, J.; Lemaire, S.D.; Hecker, A.; Dhalleine, T.; Buchanan, B.B.; Reski, R.; Einsle, O.; et al. Chloroplast FBPase and SBPase are thioredoxin-linked enzymes with similar architecture but different evolutionary histories. Proc. Natl. Acad. Sci. USA 2016, 113, 6779–6784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokochi, Y.; Yoshida, K.; Hahn, F.; Miyagi, A.; Wakabayashi, K.I.; Kawai-Yamada, M.; Weber, A.; Hisabori, T. Redox regulation of NADP-malate dehydrogenase is vital for land plants under fluctuating light environment. Proc. Natl. Acad. Sci. USA 2021, 118, e2016903118. [Google Scholar] [CrossRef]
- The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [Green Version]
- Serrato, A.J.; Fernández-Trijueque, J.; Barajas-López, J.D.; Chueca, A.; Sahrawy, M. Plastid thioredoxins: A “one-for-all” redox-signaling system in plants. Front. Plant Sci. 2013, 4, 463. [Google Scholar] [CrossRef] [Green Version]
- Sahrawy, M.; Hecht, V.; Lopez-Jaramillo, J.; Chueca, A.; Chartier, Y.; Meyer, Y. Intron position as an evolutionary marker of thioredoxins and thioredoxin domains. J. Mol. Evol. 1996, 42, 422–431. [Google Scholar] [CrossRef]
- Motohashi, K.; Kondoh, A.; Stumpp, M.T.; Hisabori, T. Comprehensive survey of proteins targeted by chloroplast thioredoxin. Proc. Natl. Acad. Sci. USA 2001, 8, 11224–11229. [Google Scholar] [CrossRef] [Green Version]
- Collin, V.; Lamkemeyer, P.; Miginiac-Maslow, M.; Hirasawa, M.; Knaff, D.B.; Dietz, K.-J.; Issakidis-Bourguet, E. Characterization of plastidial thioredoxins from Arabidopsis belonging to the new y-type. Plant Physiol. 2004, 136, 4088–4095. [Google Scholar] [CrossRef] [Green Version]
- Chibani, K.; Wingsle, G.; Jacquot, J.P.; Gelhaye, E.; Rouhier, N. Comparative genomic study of the thioredoxin family in photosynthetic organisms with emphasis on Populus trichocarpa. Mol. Plant 2009, 2, 308–322. [Google Scholar] [CrossRef]
- Fernández-Trijueque, J.; Serrato, A.J.; Sahrawy, M. Proteomic analyses of thioredoxins f and m Arabidopsis thaliana mutants indicate specific functions for these proteins in plants. Antioxidants 2019, 8, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrato, A.J.; Rojas-González, J.A.; Torres-Romero, D.; Vargas, P.; Mérida, Á.; Sahrawy, M. Thioredoxins m are major players in the multifaceted light-adaptive response in Arabidopsis thaliana. Plant J. 2021, 108, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Vieira Dos Santos, C.; Rey, P. Plant thioredoxins are key actors in the oxidative stress response. Trends Plant Sci. 2006, 11, 329–334. [Google Scholar] [CrossRef]
- Meyer, Y.; Belin, C.; Delorme-Hinoux, V.; Reichheld, J.P.; Riondet, C. Thioredoxin and glutaredoxin systems in plants: Molecular mechanisms, crosstalks, and functional significance. Antioxid. Redox Signal. 2012, 17, 1124–1160. [Google Scholar] [CrossRef] [PubMed]
- Geigenberger, P.; Thormählen, I.; Daloso, D.M.; Fernie, A.R. The unprecedented versatility of the plant thioredoxin system. Trends Plant Sci. 2017, 22, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Barajas-López, J.D.; Serrato, A.J.; Olmedilla, A.; Chueca, A.; Sahrawy, M. Localization in roots and flowers of pea chloroplast thioredoxin f and m proteins reveals new roles in non-photosynthetic organs. Plant Physiol. 2007, 145, 946–960. [Google Scholar] [CrossRef] [Green Version]
- Balmer, Y.; Vensel, W.H.; DuPont, F.M.; Buchanan, B.B.; Hurkman, W.J. Proteome of amyloplasts isolated from developing wheat endosperm presents evidence of broad metabolic capability. J. Exp. Bot. 2006, 57, 1591–1602. [Google Scholar] [CrossRef] [Green Version]
- Lawson, T.; Vialet-Chabrand, S. Speedy stomata, photosynthesis and plant water use efficiency. New Phytol. 2016, 221, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Sasidharan, R.; Schippers, J.; Schmidt, R.R. Redox and low-oxygen stress: Signal integration and interplay. Plant Physiol. 2021, 186, 66–78. [Google Scholar] [CrossRef]
- Considine, M.J.; Foyer, C.H. Oxygen and reactive oxygen species-dependent regulation of plant growth and development. Plant Physiol. 2021, 186, 79–92. [Google Scholar] [CrossRef]
- Yoshida, K.; Hisabory, T. Two distinct redox cascades cooperatively regulate chloroplast functions and sustain plant viability. Proc. Natl. Acad. Sci. USA 2016, 113, 3967–3976. [Google Scholar] [CrossRef] [Green Version]
- Serrato, A.J.; Romero-Puertas, M.C.; Lázaro-Payo, A.; Sahrawy, M. Regulation by S-nitrosylation of the Calvin-Benson cycle fructose-1,6-bisphosphatase in Pisum sativum. Redox Biol. 2018, 14, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Kurose, T.; Hino, T.; Tanaka, K.; Kawamukai, M.; Niwa, Y.K.; Matsuoka, K.; Jinbo, T.; Kimura, T. Development of series of gateway binary vectors, pGWBs for realizing efficient construction of fusion genes for plant transformation. J. Biosci. Bioeng. 2007, 104, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Heim, R.; Cubitt, A.B.; Tsien, R.Y. Improved green fluorescence. Nature 1995, 373, 663–664. [Google Scholar] [CrossRef] [PubMed]
- Alassimone, J.; Naseer, S.; Geldner, N. A developmental framework for endodermal differentiation and polarity. Proc. Natl. Acad. Sci. USA 2010, 107, 5214–5219. [Google Scholar] [CrossRef] [Green Version]
- Rojas-González, J.A.; Soto-Súarez, M.; García-Díaz, A.; Romero-Puertas, M.C.; Sandalio, L.M.; Mérida, A.; Thormählen, I.; Geigenberger, P.; Serrato, A.J.; Sahrawy, M. Disruption of both chloroplastic and cytosolic FBPases genes results in dwarf phenotype and important starch and metabolite changes in Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 2673–2689. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Chen, W. The role of autophagy in chloroplast degradation and chlorophagy in immune defenses during Pst DC3000 (AvrRps4) infection. PLoS ONE 2013, 8, e73091. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Sullivan, R.W.; Kight, A.; Henry, R.L.; Huang, J.; Jones, A.M.; Korth, K.L. Deletion of the chloroplast-localized Thylakoid formation1 gene product in Arabidopsis leads to deficient thylakoid formation and variegated leaves. Plant Physiol. 2004, 136, 3594–3604. [Google Scholar] [CrossRef] [Green Version]
- Ojeda, V.; Pérez-Ruiz, J.M.; González, M.; Nájera, V.A.; Sahrawy, M.; Serrato, A.J.; Geigenberger, P.; Cejudo, F.J. NADPH thioredoxin teductase C and thioredoxins act concertedly in seedling development. Plant Physiol. 2017, 174, 1436–1448. [Google Scholar] [CrossRef] [Green Version]
- Pagano, E.A.; Chueca, A.; López-Gorgé, J. Expression of thioredoxins f and m, and of their targets fructose-1,6-bisphosphatase and NADP-malate dehydrogenase, in pea plants grown under normal and light/temperature stress conditions. J. Exp. Bot. 2000, 51, 1299–1307. [Google Scholar]
- Traverso, J.A.; Vignols, F.; Cazalis, R.; Serrato, A.J.; Pulido, P.; Sahrawy, M.; Meyer, Y.; Cejudo, F.J.; Chueca, A. Immunocytochemical localization of Pisum sativum TRXs f and m in non-photosynthetic tissues. J. Exp. Bot. 2008, 59, 1267–1277. [Google Scholar] [CrossRef] [Green Version]
- Née, G.; Châtel-Innocenti, G.; Meimoun, P.; Leymarie, J.; Montrichard, F.; Satour, P.; Bailly, C.; Issakidis-Bourguet, E. A new role for plastid thioredoxins in seed physiology in relation to hormone regulation. Int. J. Mol. Sci. 2021, 22, 10395. [Google Scholar] [CrossRef] [PubMed]
- Serrato, A.J.; Pérez-Ruiz, J.M.; Spínola, M.C.; Cejudo, F.J. A novel NADPH thioredoxin reductase, localized in the chloroplast, which deficiency causes hypersensitivity to abiotic stress in Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 43821–43827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, Y.; Buchanan, B.B.; Vignols, F.; Reichheld, J.P. Thioredoxins and glutaredoxins: Unifying elements in redox biology. Annu. Rev. Genet. 2009, 43, 335–367. [Google Scholar] [CrossRef]
- Korkuc, P.; Schippers, J.H.; Walther, D. Characterization and identification of cis-regulatory elements in Arabidopsis based on single-nucleotide polymorphism information. Plant Physiol. 2014, 164, 181–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senkler, J.; Senkler, M.; Eubel, H.; Hildebrandt, T.; Lengwenus, C.; Schertl, P.; Schwarzländer, M.; Wagner, S.; Wittig, I.; Braun, H.P. The mitochondrial complexome of Arabidopsis thaliana. Plant J. 2017, 89, 1079–1092. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Takano, T.; Liu, S.; Zhang, X. Arabidopsis mitochondrial voltage-dependent anion channel 3 (AtVDAC3) protein interacts with thioredoxin m2. FEBS Lett. 2015, 589, 1207–1213. [Google Scholar] [CrossRef] [Green Version]
- Jing, X.; Yao, J.; Ma, X.; Zhang, Y.; Sun, Y.; Xiang, M.; Hou, P.; Li, N.; Zhao, R.; Li, J.; et al. Kandelia candel thioredoxin f confers osmotic stress tolerance in transgenic tobacco. Int. J. Mol. Sci. 2020, 21, 3335. [Google Scholar] [CrossRef]
- Montillet, J.L.; Rondet, D.; Brugière, S.; Henri, P.; Rumeau, D.; Reichheld, J.P.; Couté, Y.; Leonhardt, N.; Rey, P. Plastidial and cytosolic thiol reductases participate in the control of stomatal functioning. Plant Cell Environ. 2021, 44, 1417–1435. [Google Scholar] [CrossRef]
- Ballicora, M.A.; Frueauf, J.B.; Fu, Y.; Schürmann, P.; Preiss, J. Activation of the potato tuber ADP-glucose pyrophosphorylase by thioredoxin. J. Biol. Chem. 2000, 275, 1315–1320. [Google Scholar] [CrossRef] [Green Version]
- Valerio, C.; Costa, A.; Marri, L.; Issakidis-Bourguet, E.; Pupillo, P.; Trost, P.; Sparla, F. Thioredoxin-regulated β-amylase (BAM1) triggers diurnal starch degradation in guard cells, and in mesophyll cells under osmotic stress. J. Exp. Bot. 2011, 62, 545–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santelia, D.; Lunn, J.E. Transitory Starch Metabolism in Guard Cells: Unique Features for a Unique Function. Plant Physiol. 2017, 174, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courteille, A.; Vesa, S.; Sanz-Barrio, R.; Cazale, A.C.C.; Becuwe-Linka, N.; Farran, I.; Havaux, M.; Rey, P.; Rumeau, D. Thioredoxin m4 controls photosynthetic alternative electron pathways in Arabidopsis. Plant Physiol. 2013, 161, 508–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mestres-Ortega, D.; Meyer, Y. The Arabidopsis thaliana genome encodes at least four thioredoxins m and a new prokaryotic-like thioredoxin. Gene 1999, 240, 307–316. [Google Scholar] [CrossRef]
- Becker, J.D.; Boavida, L.C.; Carneiro, J.; Haury, M.; Feijó, J.A. Transcriptional profiling of Arabidopsis tissues reveals the unique characteristics of the pollen transcriptome. Plant Physiol. 2003, 133, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Belin, C.; Bashandy, T.; Cela, J.; Delorme-Hinoux, V.; Riondet, C.; Reichheld, J.P. A comprehensive study of thiol reduction gene expression under stress conditions in Arabidopsis thaliana. Plant Cell Environ. 2015, 38, 299–314. [Google Scholar] [CrossRef]
- Azri, W.; Brunel, N.; Franchel, J.; Ben Rejeb, I.; Jacquot, J.P.; Julien, J.L.; Herbette, S.; Roeckel-Drevet, P. Putative involvement of Thioredoxin h in early response to gravitropic stimulation of poplar stems. J. Plant Physiol. 2013, 170, 707–711. [Google Scholar] [CrossRef]
- Benitez-Alfonso, Y.; Cilia, M.; San Roman, A.; Thomas, C.; Maule, A.; Hearn, S.; Jackson, D. Control of Arabidopsis meristem development by thioredoxin-dependent regulation of intercellular transport. Proc. Natl. Acad. Sci. USA 2009, 106, 3615–3620. [Google Scholar] [CrossRef] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahrawy, M.; Fernández-Trijueque, J.; Vargas, P.; Serrato, A.J. Comprehensive Expression Analyses of Plastidial Thioredoxins of Arabidopsis thaliana Indicate a Main Role of Thioredoxin m2 in Roots. Antioxidants 2022, 11, 1365. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11071365
Sahrawy M, Fernández-Trijueque J, Vargas P, Serrato AJ. Comprehensive Expression Analyses of Plastidial Thioredoxins of Arabidopsis thaliana Indicate a Main Role of Thioredoxin m2 in Roots. Antioxidants. 2022; 11(7):1365. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11071365
Chicago/Turabian StyleSahrawy, Mariam, Juan Fernández-Trijueque, Paola Vargas, and Antonio J. Serrato. 2022. "Comprehensive Expression Analyses of Plastidial Thioredoxins of Arabidopsis thaliana Indicate a Main Role of Thioredoxin m2 in Roots" Antioxidants 11, no. 7: 1365. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11071365