
Human Placental Extract Delays In Vitro Cellular Senescence through the Activation of NRF2-Mediated Antioxidant Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Cell Proliferation Assay
2.4. Senescence-Associated β-Galactosidase Activity
2.5. Quantitative PCR Analysis
2.6. Intracellular Reactive Oxygen Species Scavenging Activity Assay
2.7. RNA-Sequencing
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. HPE Treatment Delays the Onset of Cellular Senescence Induced by Chronic Oxidative Stress
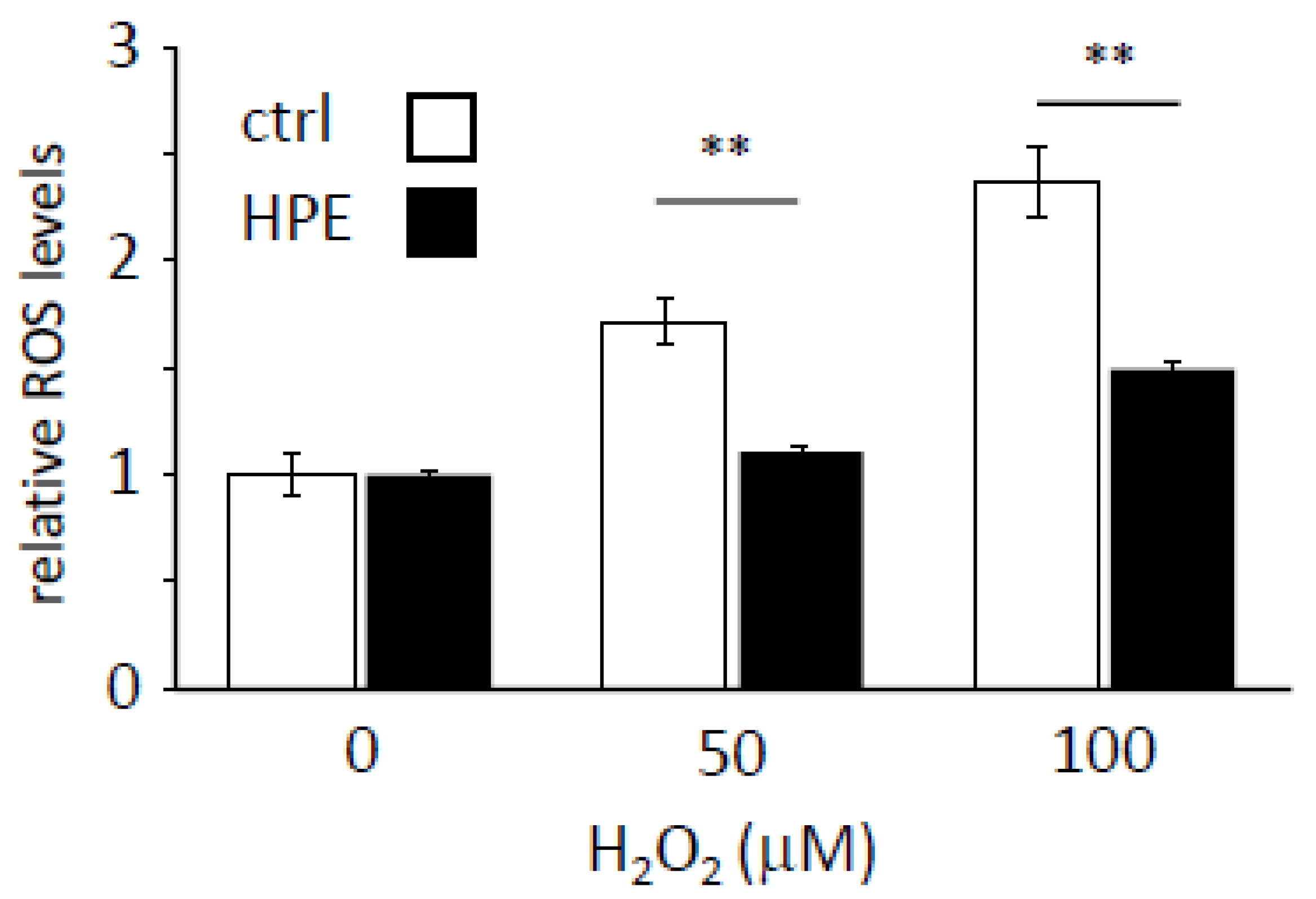
3.2. HPE Enhances the Scavenging of Cellular ROS Levels
3.3. HPE Upregulates a Set of Antioxidant Genes under H2O2-Treated Conditions
3.4. HPE Increases NRF2 Protein Levels under Oxidative Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Characteristics of the Aging Skin. Adv. Wound Care 2013, 2, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Mesa-Arango, A.C.; Flórez-Muñoz, S.V.; Sanclemente, G. Mechanisms of skin aging. Iatreia 2017, 30, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Yagi, M.; Yonei, Y. Glycative stress and anti-aging: 7. Glycative stress and skin aging. Glycative Stress Res. 2018, 5, 50–54. [Google Scholar]
- Bocheva, G.; Slominski, R.M.; Slominski, A.T. Neuroendocrine Aspects of Skin Aging. Int. J. Mol. Sci. 2019, 20, 2798. [Google Scholar] [CrossRef] [Green Version]
- Melzer, D.; Pilling, L.C.; Ferrucci, L. The genetics of human ageing. Nat. Rev. Genet. 2020, 21, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Emanuelli, M.; Sartini, D.; Molinelli, E.; Campagna, R.; Pozzi, V.; Salvolini, E.; Simonetti, O.; Campanati, A.; Offidani, A. The Double-Edged Sword of Oxidative Stress in Skin Damage and Melanoma: From Physiopathology to Therapeutical Approaches. Antioxidants 2022, 11, 612. [Google Scholar] [CrossRef] [PubMed]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.L. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Murakami, S.; Motohashi, H. Roles of Nrf2 in cell proliferation and differentiation. Free Radic. Biol. Med. 2015, 88, 168–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tossetta, G.; Fantone, S.; Montanari, E.; Marzioni, D.; Goteri, G. Role of NRF2 in Ovarian Cancer. Antioxidants 2022, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, Y.; Kagawa, S.; Kobayashi, H.; Saito, M.; Iwasaki, Y.; Kimura, Y.; Kobayashi, K.; Tsuyuguchi, M. Clinical treatment test of Melsmon on menopausal disorder. Medicat. Treat 1981, 9, 1–10. [Google Scholar]
- Tiwary, S.K.; Shukla, D.; Tripathi, A.K.; Agrawal, S.; Singh, M.K.; Shukla, V.K. Effect of placental-extract gel and cream on non-healing wounds. J. Wound Care 2006, 15, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.H.; Lee, E.J.; Lee, S.Y.; Cho, S.J.; Hong, Y.S.; Park, S.B. Effect of human placental extract on menopausal symptoms, fatigue, and risk factors for cardiovascular disease in middle-aged Korean women. Menopause 2008, 15, 296–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, M.; Park, S.B. Effect of human placental extract on health status in elderly Koreans. Evid.-Based Complementary Altern. Med. Ecam 2012, 2012, 732915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.Y.; Lee, K.; Lee, S.M.; Yoo, S.H.; Hwang, S.G.; Choi, J.Y.; Lee, S.W.; Hwang, J.S.; Kim, K.K.; Kang, H.C.; et al. Efficacy and Safety of Human Placental Extract for Alcoholic and Nonalcoholic Steatohepatitis An Open Label, Randomized, Comparative Study. Biol. Pharm. Bull. 2014, 37, 1857–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, C.; Koike, K.; Takano, F.; Sugiur, K.; Suzuki, N. Efficacy of porcine placental extract on wrinkle widths below the eye in climacteric women. Climacteric 2014, 17, 370–376. [Google Scholar] [CrossRef]
- Nagae, M.; Nagata, M.; Teramoto, M.; Yamakawa, M.; Matsuki, T.; Ohnuki, K.; Shimizu, K. Effect of Porcine Placenta Extract Supplement on Skin Condition in Healthy Adult Women: A Randomized, Double-Blind Placebo-Controlled Study. Nutrients 2020, 12, 1671. [Google Scholar] [CrossRef]
- Watanabe, S.; Togashi, S.; Takahashi, N.; Fukui, T. L-tryptophan as an antioxidant in human placenta extract. J. Nutr. Sci. Vitam. 2002, 48, 36–39. [Google Scholar] [CrossRef]
- Togashi, S.; Takahashi, N.; Iwama, M.; Watanabe, S.; Tamagawa, K.; Fukui, T. Antioxidative collagen-derived peptides in human-placenta extract. Placenta 2002, 23, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Bak, D.H.; Na, J.; Choi, M.J.; Lee, B.C.; Oh, C.T.; Kim, J.Y.; Han, H.J.; Kim, M.J.; Kim, T.H.; Kim, B.J. Anti-apoptotic effects of human placental hydrolysate against hepatocyte toxicity in vivo and in vitro. Int. J. Mol. Med. 2018, 42, 2569–2583. [Google Scholar] [CrossRef] [Green Version]
- Bak, D.H.; Na, J.; Im, S.I.; Oh, C.T.; Kim, J.Y.; Park, S.K.; Han, H.J.; Seok, J.; Choi, S.Y.; Ko, E.J.; et al. Antioxidant effect of human placenta hydrolysate against oxidative stress on muscle atrophy. J. Cell. Physiol. 2019, 234, 1643–1658. [Google Scholar] [CrossRef] [PubMed]
- Samiei, F.; Jamshidzadeh, A.; Noorafshan, A.; Ghaderi, A. Human Placental Extract Ameliorates Structural Lung Changes Iinduced by Amiodarone in Rats. Iran. J. Pharm. Res. 2016, 15, 75–82. [Google Scholar] [PubMed]
- Ghoneum, M.; El-Gerbed, M.S.A. Human placental extract ameliorates methotrexate-induced hepatotoxicity in rats via regulating antioxidative and anti-inflammatory responses. Cancer Chemother. Pharm. 2021, 88, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.Y.; Chin, L.C.; Kimura, K.; Nakahata, Y. Human placental extract activates a wide array of gene expressions related to skin functions. Sci. Rep. 2022, 12, 11031. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Phark, S.; Lee, M.; Lim, J.Y.; Sul, D. Anti-oxidative and anti-inflammatory activities of placental extracts in benzo[a]pyrene-exposed rats. Placenta 2010, 31, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Campagna, R.; Pozzi, V.; Sartini, D.; Salvolini, E.; Brisigotti, V.; Molinelli, E.; Campanati, A.; Offidani, A.; Emanuelli, M. Beyond Nicotinamide Metabolism: Potential Role of Nicotinamide N-Methyltransferase as a Biomarker in Skin Cancers. Cancers 2021, 13, 4943. [Google Scholar] [CrossRef] [PubMed]
- Bataille, A.M.; Manautou, J.E. Nrf2: A potential target for new therapeutics in liver disease. Clin. Pharmacol. Ther. 2012, 92, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Majumdar, R.; Tsuji, Y. Nuclear lamins and progerin are dispensable for antioxidant Nrf2 response to arsenic and cadmium. Cell. Signal. 2017, 33, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; O’Connor, T.; Yamamoto, M. Keap1 regulates both cytoplasmic-nuclear shuttling and degradation of Nrf2 in response to electrophiles. Genes Cells 2003, 8, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Kang, M.I.; Okawa, H.; Ohtsuji, M.; Zenke, Y.; Chiba, T.; Igarashi, K.; Yamamoto, M. Oxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2. Mol. Cell. Biol. 2004, 24, 7130–7139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayers, D.; Baron, B.; Hunter, T. miRNA Influences in NRF2 Pathway Interactions within Cancer Models. J. Nucleic Acids 2015, 2015, 143636. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.; Chin, L.-C.; Kimura, K.; Nakahata, Y. Human Placental Extract Delays In Vitro Cellular Senescence through the Activation of NRF2-Mediated Antioxidant Pathway. Antioxidants 2022, 11, 1545. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11081545
Huang L, Chin L-C, Kimura K, Nakahata Y. Human Placental Extract Delays In Vitro Cellular Senescence through the Activation of NRF2-Mediated Antioxidant Pathway. Antioxidants. 2022; 11(8):1545. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11081545
Chicago/Turabian StyleHuang, Liguo, Lit-Chein Chin, Koichi Kimura, and Yasukazu Nakahata. 2022. "Human Placental Extract Delays In Vitro Cellular Senescence through the Activation of NRF2-Mediated Antioxidant Pathway" Antioxidants 11, no. 8: 1545. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11081545