
Evaluation of Different Advanced Approaches to Simulation of Dynamic In Vitro Digestion of Polyphenols from Different Food Matrices—A Systematic Review
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
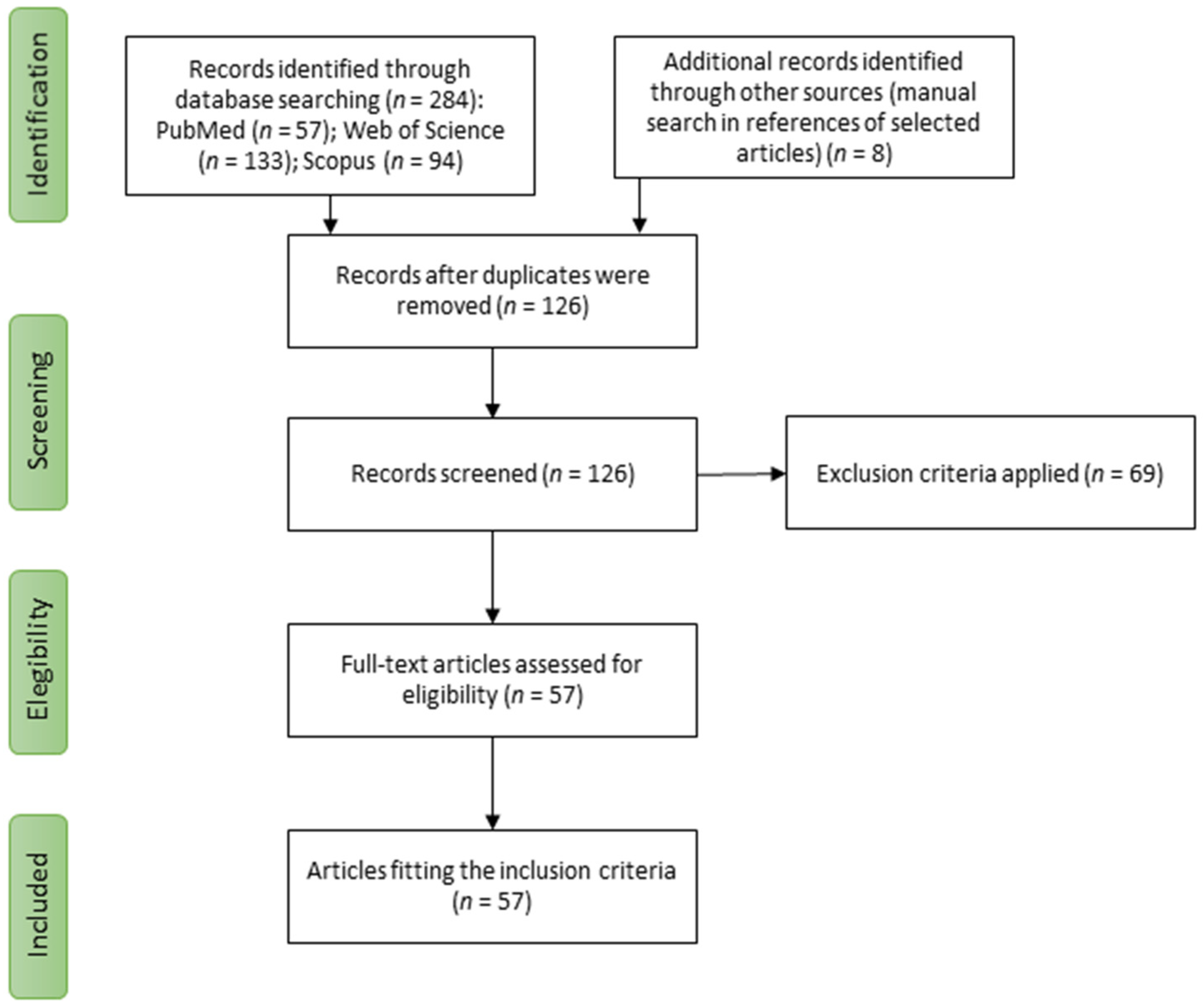
3.1. Study Selection
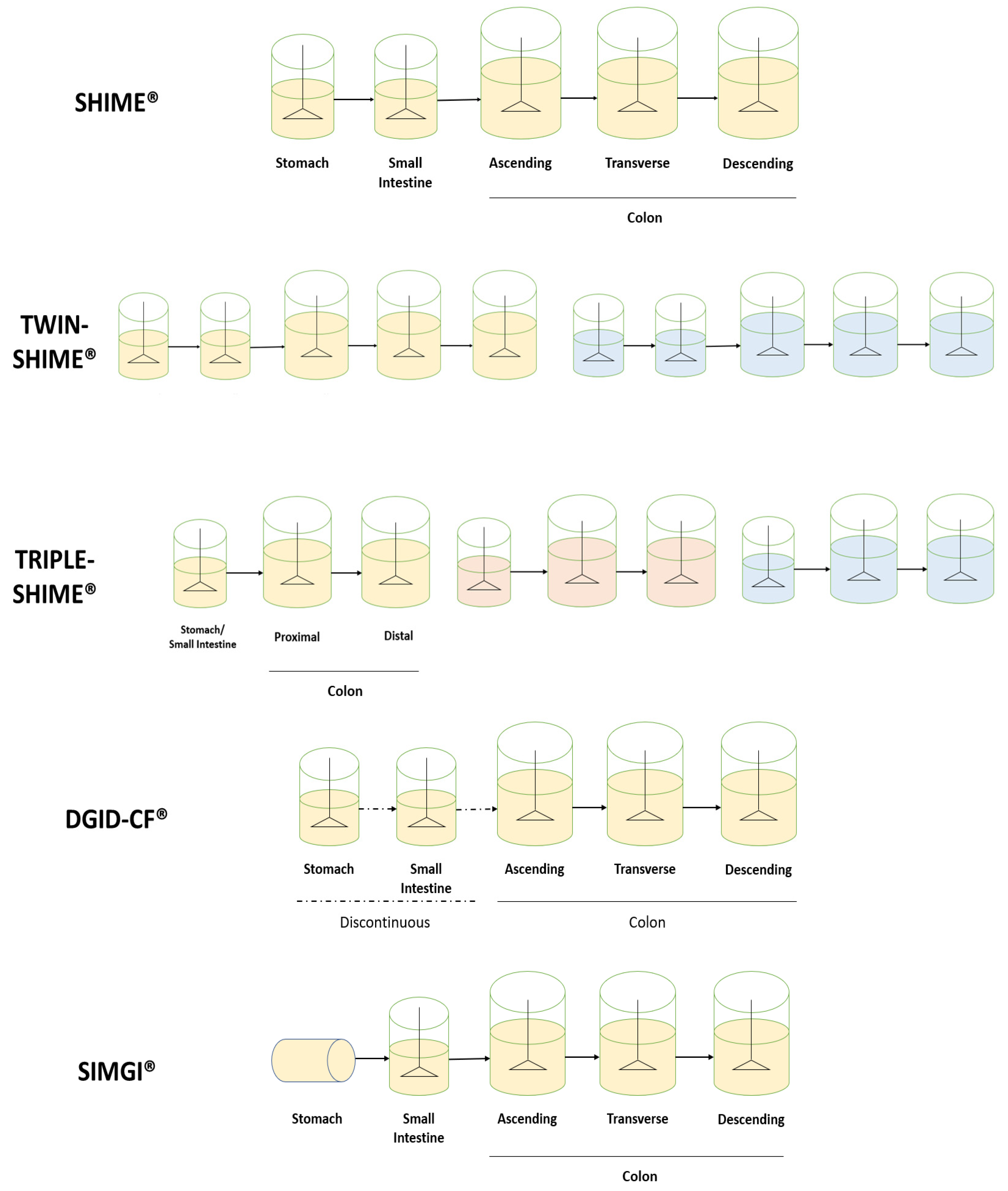
3.2. Dynamic In Vitro Digestion Models
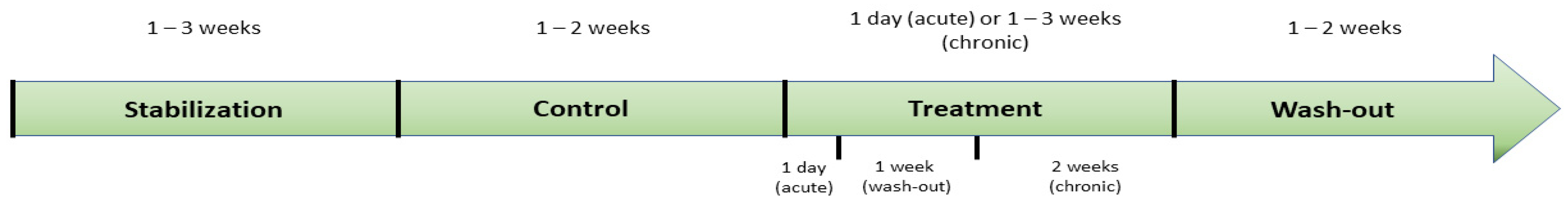
3.3. Assay Conditions: Experimental Design
3.3.1. Equipment Configuration
3.3.2. Experimental Steps
3.3.3. Digestive Conditions
3.3.4. Dose and Administration
3.4. Sample Treatment
3.5. Microbiological Analyses
3.6. Phenolic Compound Identification and Bioaccessibility
3.7. Antioxidant Activity of Samples and Digestates
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramalingum, N.; Mahomoodally, M.F. The Therapeutic Potential of Medicinal Foods. Adv. Pharmacol. Sci. 2014, 2014, 354264. [Google Scholar] [CrossRef] [Green Version]
- Arablou, T.; Aryaeian, N. The effect of ginger (Zingiber Officinale) as an ancient medicinal plant on improving blood lipids. J. Herb. Med. 2018, 12, 11–15. [Google Scholar] [CrossRef]
- Mickymaray, S. Efficacy and Mechanism of Traditional Medicinal Plants and Bioactive Compounds against Clinically Important Pathogens. Antibiotics 2019, 8, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christaki, E.; Bonos, E.; Giannenas, I.; Florou-Paneri, P. Aromatic Plants as a Source of Bioactive Compounds. Agriculture 2012, 2, 228–243. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF/MS Characterization of Phenolic Compounds from Medicinal Plants (Hops and Juniper Berries) and Their Antioxidant Activity. Foods 2019, 9, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirar, A.I.; Alsaadi, D.H.M.; Wada, M.; Mohamed, M.A.; Watanabe, T.; Devkota, H.P. Effects of extraction solvents on total phenolic and flavonoid contents and biological activities of extracts from Sudanese medicinal plants. S. Afr. J. Bot. 2019, 120, 261–267. [Google Scholar] [CrossRef]
- Olech, M.; Nowak, R.; Ivanova, D.; Tashev, A.; Boyadzhieva, S.; Kalotova, G.; Angelov, G.; Gawlik-Dziki, U. LC-ESI-MS/MS-MRM Profiling of Polyphenols and Antioxidant Activity Evaluation of Junipers of Different Origin. Appl. Sci. 2020, 10, 8921. [Google Scholar] [CrossRef]
- Patil, B.S.; Jayaprakasha, G.K.; Chidambara Murthy, K.N.; Vikram, A. Bioactive compounds: Historical perspectives, opportunities, and challenges. J. Agric. Food Chem. 2009, 57, 8142–8160. [Google Scholar] [CrossRef]
- Câmara, J.S.; Albuquerque, B.R.; Aguiar, J.; Corrêa, R.C.G.; Gonçalves, J.L.; Granato, D.; Pereira, J.A.M.; Barros, L.; Ferreira, I.C.F.R. Food Bioactive Compounds and Emerging Techniques for Their Extraction: Polyphenols as a Case Study. Foods 2020, 10, 37. [Google Scholar] [CrossRef]
- Ali, H.M.; Abo-Shady, A.; Sharaf Eldeen, H.A.; Soror, H.A.; Shousha, W.G.; Abdel-Barry, O.A.; Saleh, A.M. Structural features, kinetics and SAR study of radical scavenging and antioxidant activities of phenolic and anilinic compounds. Chem. Cent. J. 2013, 7, 53. [Google Scholar] [CrossRef]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A concise overview on the chemistry, occurrence, and human health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouarab Chibane, L.; Degraeve, P.; Ferhout, H.; Bouajila, J.; Oulahal, N. Plant antimicrobial polyphenols as potential natural food preservatives. J. Sci. Food Agric. 2019, 99, 1457–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musarra-Pizzo, M.; Ginestra, G.; Smeriglio, A.; Pennisi, R.; Sciortino, M.T.; Mandalari, G. The Antimicrobial and Antiviral Activity of Polyphenols from Almond (Prunus dulcis L.) Skin. Nutrients 2019, 11, 2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef] [PubMed]
- Denev, P.; Číž, M.; Kratchanova, M.; Blazheva, D. Black chokeberry (Aronia melanocarpa) polyphenols reveal different antioxidant, antimicrobial and neutrophil-modulating activities. Food Chem. 2019, 284, 108–117. [Google Scholar] [CrossRef]
- Ford, L.; Stratakos, A.C.; Theodoridou, K.; Dick, J.T.A.; Sheldrake, G.N.; Linton, M.; Corcionivoschi, N.; Walsh, P.J. Polyphenols from Brown Seaweeds as a Potential Antimicrobial Agent in Animal Feeds. ACS Omega 2020, 5, 9093. [Google Scholar] [CrossRef] [Green Version]
- Montenegro-Landívar, M.F.; Tapia-Quirós, P.; Vecino, X.; Reig, M.; Valderrama, C.; Granados, M.; Cortina, J.L.; Saurina, J. Polyphenols and their potential role to fight viral diseases: An overview. Sci. Total Environ. 2021, 801, 149719. [Google Scholar] [CrossRef]
- Chojnacka, K.; Witek-Krowiak, A.; Skrzypczak, D.; Mikula, K.; Młynarz, P. Phytochemicals containing biologically active polyphenols as an effective agent against COVID-19-inducing coronavirus. J. Funct. Foods 2020, 73, 104146. [Google Scholar] [CrossRef]
- Blank, D.E.; Hübner, S.d.O.; Alves, G.H.; Cardoso, C.A.L.; Freitag, R.A.; Cleff, M.B.; Blank, D.E.; Hübner, S.d.O.; Alves, G.H.; Cardoso, C.A.L.; et al. Chemical Composition and Antiviral Effect of Extracts of Origanum vulgare. J. Adv. Biol. Biotechnol. 2019, 10, 188–196. [Google Scholar] [CrossRef] [Green Version]
- Iloghalu, U.; Holmes, B.; Khatiwada, J.; Williams, L.L.; Iloghalu, U.; Holmes, B.; Khatiwada, J.; Williams, L.L. Selected Plant Extracts Show Antiviral Effects against Murine Norovirus Surrogate. Adv. Microbiol. 2019, 9, 372–384. [Google Scholar] [CrossRef]
- Lalani, S.; Poh, C.L. Flavonoids as Antiviral Agents for Enterovirus A71 (EV-A71). Viruses 2020, 12, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-López, P.; Lozano-Sanchez, J.; Borrás-Linares, I.; Emanuelli, T.; Menéndez, J.A.; Segura-Carretero, A. Structure–Biological Activity Relationships of Extra-Virgin Olive Oil Phenolic Compounds: Health Properties and Bioavailability. Antioxidants 2020, 9, 685. [Google Scholar] [CrossRef]
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Oniszczuk, T.; Combrzyński, M.; Nowakowska, D.; Matwijczuk, A. Influence of In Vitro Digestion on Composition, Bioaccessibility and Antioxidant Activity of Food Polyphenols-A Non-Systematic Review. Nutrients 2020, 12, 1401. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Datta, S.; Bhatia, K.S.; Bhumika, J.P.; Prasad, R. Role of plant derived bioactive compounds against cancer. S. Afr. J. Bot. 2022, 149, 1017–1028. [Google Scholar] [CrossRef]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisi, O.I.; Puoci, F.; Restuccia, D.; Farina, G.; Iemma, F.; Picci, N. Polyphenols and Their Formulations: Different Strategies to Overcome the Drawbacks Associated with Their Poor Stability and Bioavailability. In Polyphenols in Human Health and Disease, 1st ed.; Watson, R., Preedy, V., Zibadi, S., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 1, pp. 29–45. [Google Scholar] [CrossRef]
- McClements, D.J. Advances in nanoparticle and microparticle delivery systems for increasing the dispersibility, stability, and bioactivity of phytochemicals. Biotechnol. Adv. 2020, 38, 107287. [Google Scholar] [CrossRef]
- Catalkaya, G.; Venema, K.; Lucini, L.; Rocchetti, G.; Delmas, D.; Daglia, M.; De Filippis, A.; Xiao, H.; Quiles, J.L.; Xiao, J.; et al. Interaction of dietary polyphenols and gut microbiota: Microbial metabolism of polyphenols, influence on the gut microbiota, and implications on host health. Food Front. 2020, 1, 109–133. [Google Scholar] [CrossRef]
- Ahmad-Qasem, M.H.; Cánovas, J.; Barrajón-Catalán, E.; Carreres, J.E.; Micol, V.; García-Pérez, J.V. Influence of olive leaf processing on the bioaccessibility of bioactive polyphenols. J. Agric. Food Chem. 2014, 62, 6190–6198. [Google Scholar] [CrossRef]
- Li, C.; Yu, W.; Wu, P.; Chen, X.D. Current in vitro digestion systems for understanding food digestion in human upper gastrointestinal tract. Trends Food Sci. Technol. 2020, 96, 114–126. [Google Scholar] [CrossRef]
- Bohn, T.; Carriere, F.; Day, L.; Deglaire, A.; Egger, L.; Freitas, D.; Golding, M.; Le Feunteun, S.; Macierzanka, A.; Menard, O.; et al. Correlation between in vitro and in vivo data on food digestion. What can we predict with static in vitro digestion models? Crit. Rev. Food Sci. Nutr. 2018, 58, 2239–2261. [Google Scholar] [CrossRef] [Green Version]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Zhang, Z.; Muriel Mundo, J.; McClements, D.J. Factors impacting lipid digestion and nutraceutical bioaccessibility assessed by standardized gastrointestinal model (INFOGEST): Emulsifier type. Food Res. Int. 2020, 137, 109739. [Google Scholar] [CrossRef]
- Reboredo-Rodríguez, P.; Olmo-García, L.; Figueiredo-González, M.; González-Barreiro, C.; Carrasco-Pancorbo, A.; Cancho-Grande, B. Application of the INFOGEST Standardized Method to Assess the Digestive Stability and Bioaccessibility of Phenolic Compounds from Galician Extra-Virgin Olive Oil. J. Agric. Food Chem. 2021, 69, 11592–11605. [Google Scholar] [CrossRef]
- Makran, M.; Faubel, N.; López-García, G.; Cilla, A.; Barberá, R.; Alegría, A.; Garcia-Llatas, G. Sterol bioaccessibility in a plant sterol-enriched beverage using the INFOGEST digestion method: Influence of gastric lipase, bile salts and cholesterol esterase. Food Chem. 2022, 382, 132305. [Google Scholar] [CrossRef] [PubMed]
- Alegría, A.; Garcia-Llatas, G.; Cilla, A. Static Digestion Models: General Introduction. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer: Cham, Switzerland; New York, NY, USA, 2015; pp. 3–12. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [Green Version]
- Thuenemann, E.C.; Giuseppina, G.M.; Rich, G.T.; Faulks, R.M. Dynamic Gastric Model (DGM). In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer: Cham, Switzerland; New York, NY, USA, 2015; pp. 47–59. [Google Scholar] [CrossRef] [Green Version]
- Barros, L.; Retamal, C.; Torres, H.; Zúñiga, R.N.; Troncoso, E. Development of an in vitro mechanical gastric system (IMGS) with realistic peristalsis to assess lipid digestibility. Int. Food Res. J. 2016, 90, 216–225. [Google Scholar] [CrossRef]
- Kozu, H.; Nakata, Y.; Nakajima, M.; Neves, M.A.; Uemura, K.; Sato, S.; Kobayashi, I.; Ichikawa, S. Development of a Human Gastric Digestion Simulator Equipped with Peristalsis Function for the Direct Observation and Analysis of the Food Digestion Process. Food Sci. Technol. Res. 2014, 20, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Cordonnier, C.; Thévenot, J.; Etienne-Mesmin, L.; Denis, S.; Alric, M.; Livrelli, V.; Blanquet-Diot, S. Dynamic In Vitro Models of the Human Gastrointestinal Tract as Relevant Tools to Assess the Survival of Probiotic Strains and Their Interactions with Gut Microbiota. Microorganisms 2015, 3, 725–745. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Singh, R.P. A Human Gastric Simulator (HGS) to Study Food Digestion in Human Stomach. J. Food Sci. 2010, 75, E627–E635. [Google Scholar] [CrossRef]
- López de Lacey, A.M.; Giménez, B.; Pérez-Santín, E.; Faulks, R.; Mandalari, G.; López-Caballero, M.E.; Montero, P. Bioaccessibility of green tea polyphenols incorporated into an edible agar film during simulated human digestion. Food Res. Int. 2012, 48, 462–469. [Google Scholar] [CrossRef]
- Mandalari, G.; Bisignano, C.; Filocamo, A.; Chessa, S.; Sarò, M.; Torre, G.; Faulks, R.M.; Dugo, P. Bioaccessibility of pistachio polyphenols, xanthophylls, and tocopherols during simulated human digestion. Nutrition 2013, 29, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Cueva, C.; Gil-Sánchez, I.; Tamargo, A.; Miralles, B.; Crespo, J.; Bartolomé, B.; Moreno-Arribas, M.V. Gastrointestinal digestion of food-use silver nanoparticles in the dynamic SIMulator of the GastroIntestinal tract (simgi ®). Impact on human gut microbiota. Food Chem. Toxicol. 2019, 132, 110657. [Google Scholar] [CrossRef] [PubMed]
- Dupont, D.; Alric, M.; Blanquet-Diot, S.; Bornhorst, G.; Cueva, C.; Deglaire, A.; Denis, S.; Ferrua, M.; Havenaar, R.; Lelieveld, J.; et al. Can dynamic in vitro digestion systems mimic the physiological reality? Crit. Rev. Food Sci. Nutr. 2019, 59, 1546–1562. [Google Scholar] [CrossRef] [Green Version]
- Guerra, A.; Denis, S.; le Goff, O.; Sicardi, V.; François, O.; Yao, A.F.; Garrait, G.; Manzi, A.P.; Beyssac, E.; Alric, M.; et al. Development and validation of a new dynamic computer-controlled model of the human stomach and small intestine. Biotechnol. Bioeng. 2016, 113, 1325–1335. [Google Scholar] [CrossRef]
- Ménard, O.; Cattenoz, T.; Guillemin, H.; Souchon, I.; Deglaire, A.; Dupont, D.; Picque, D. Validation of a new in vitro dynamic system to simulate infant digestion. Food Chem. 2014, 145, 1039–1045. [Google Scholar] [CrossRef]
- Van de Wiele, T.; van den Abbeele, P.; Ossieur, W.; Possemiers, S.; Marzorati, M. The Simulator of the Human Intestinal Microbial Ecosystem (SHIME®). In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer: Cham, Switzerland; New York, NY, USA, 2018; pp. 305–317. [Google Scholar] [CrossRef]
- Verwei, M.; Minekus, M.; Zeijdner, E.; Schilderink, R.; Havenaar, R. Evaluation of two dynamic in vitro models simulating fasted and fed state conditions in the upper gastrointestinal tract (TIM-1 and tiny-TIM) for investigating the bioaccessibility of pharmaceutical compounds from oral dosage forms. Int. J. Pharm. 2016, 498, 178–186. [Google Scholar] [CrossRef]
- Giuliani, C.; Marzorati, M.; Innocenti, M.; Vilchez-Vargas, R.; Vital, M.; Pieper, D.H.; Van De Wiele, T.; Mulinacci, N. Dietary supplement based on stilbenes: A focus on gut microbial metabolism by the in vitro simulator M-SHIME®. Food Funct. 2016, 7, 4564–4575. [Google Scholar] [CrossRef]
- Li, Q.; Van Herreweghen, F.; De Mey, M.; Goeminne, G.; Van de Wiele, T. The Donor-Dependent and Colon-Region-Dependent Metabolism of (+)-Catechin by Colonic Microbiota in the Simulator of the Human Intestinal Microbial Ecosystem. Molecules 2022, 27, 73. [Google Scholar] [CrossRef]
- Molly, K.; Van de Woestyne, M.; De Smet, I.; Verstraete, W. Validation of the Simulator of the Human Intestinal Microbial Ecosystem (SHIME) Reactor Using Microorganism-associated Activities. Microb. Ecol. 1994, 7, 191–200. [Google Scholar] [CrossRef]
- Lila, M.A.; Ribnicky, D.M.; Rojo, L.E.; Rojas-Silva, P.; Oren, A.; Havenaar, R.; Janle, E.M.; Raskin, I.; Yousef, G.G.; Grace, M.H. Complementary approaches to gauge the bioavailability and distribution of ingested berry polyphenolics. J. Agric. Food Chem. 2012, 60, 5763–5771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krul, C.; Luiten-Schuite, A.; Tenfelde, A.; Van Ommen, B.; Verhagen, H.; Havenaar, R. Antimutagenic activity of green tea and black tea extracts studied in a dynamic in vitro gastrointestinal model. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2001, 474, 71–85. [Google Scholar] [CrossRef]
- Huang, J.; Chen, P.X.; Rogers, M.A.; Wettig, S.D. Investigating the Phospholipid Effect on the Bioaccessibility of Rosmarinic Acid-Phospholipid Complex through a Dynamic Gastrointestinal in Vitro Model. Pharmaceutics 2019, 11, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, A.R.; Pinheiro, A.C.; Vicente, A.A.; Souza-Soares, L.A.; Cerqueira, M.A. Liposomes loaded with phenolic extracts of Spirulina LEB-18: Physicochemical characterization and behavior under simulated gastrointestinal conditions. Food Res. Int. 2019, 120, 656–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz-Buenhombre, M.; Villanueva, S.; Moro, C.; Tomás-Cobos, L.; Viadel, B.; Guadarrama, A. Bioavailability and the mechanism of action of a grape extract rich in polyphenols in cholesterol homeostasis. J. Funct. Foods 2016, 21, 178–185. [Google Scholar] [CrossRef]
- Fernández-Jalao, I.; Sánchez-Moreno, C.; De Ancos, B. Influence of food matrix and high-pressure processing on onion flavonols and antioxidant activity during gastrointestinal digestion. J. Food Eng. 2017, 213, 60–68. [Google Scholar] [CrossRef]
- Herranz-López, M.; Olivares-Vicente, M.; Encinar, J.A.; Barrajón-Catalán, E.; Segura-Carretero, A.; Joven, J.; Micol, V. Multi-Targeted Molecular Effects of Hibiscus sabdariffa Polyphenols: An Opportunity for a Global Approach to Obesity. Nutrients 2017, 9, 907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, D.; Le Feunteun, S. Inhibitory effect of black tea, lemon juice, and other beverages on salivary and pancreatic amylases: What impact on bread starch digestion? A dynamic in vitro study. Food Chem. 2019, 297, 124885. [Google Scholar] [CrossRef] [Green Version]
- Maathuis, A.; Venema, K.; Hoffman, A.; Evans, A.; Sanders, L. The Effect of the Undigested Fraction of Maize Products on the Activity and Composition of the Microbiota Determined in a Dynamic in Vitro Model of the Human Proximal Large Intestine. J. Am. Coll. Nutr. 2013, 28, 657–666. [Google Scholar] [CrossRef]
- Molly, K.; Vande Woestyne, M.; Verstraete, W. Development of a 5-step multi-chamber reactor as a simulation of the human intestinal microbial ecosystem. Appl. Microbiol. Biotechnol. 1993, 39, 254–258. [Google Scholar] [CrossRef]
- Gibson, G.R.; Cummings, J.H.; Macfarlane, G.T. Use of a three-stage continuous culture system to study the effect of mucin on dissimilatory sulfate reduction and methanogenesis by mixed populations of human gut bacteria. Appl. Environ. Microbiol. 1988, 54, 2750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekbatan, S.; Sleno, L.; Sabally, K.; Khairallah, J.; Azadi, B.; Rodes, L.; Prakash, S.; Donnelly, D.J.; Kubow, S. Biotransformation of polyphenols in a dynamic multistage gastrointestinal model. Food Chem. 2016, 204, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Ekbatan, S.S.; Iskandar, M.M.; Sleno, L.; Sabally, K.; Khairallah, J.; Prakash, S.; Kubow, S. Absorption and Metabolism of Phenolics from Digests of Polyphenol-Rich Potato Extracts Using the Caco-2/HepG2 Co-Culture System. Foods 2018, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Kemperman, R.A.; Gross, G.; Mondot, S.; Possemiers, S.; Marzorati, M.; Van de Wiele, T.; Doré, J.; Vaughan, E.E. Impact of polyphenols from black tea and red wine/grape juice on a gut model microbiome. Food Res. Int. 2013, 53, 659–669. [Google Scholar] [CrossRef]
- García-Villalba, R.; Vissenaekens, H.; Pitart, J.; Romo-Vaquero, M.; Espín, J.C.; Grootaert, C.; Selma, M.V.; Raes, K.; Smagghe, G.; Possemiers, S.; et al. Gastrointestinal simulation model TWIN-SHIME shows differences between human urolithin-metabotypes in gut microbiota composition, pomegranate polyphenol metabolism, and transport along the intestinal tract. J. Agric. Food Chem. 2017, 65, 5480–5493. [Google Scholar] [CrossRef] [Green Version]
- Koper, J.E.; Loonen, L.M.; Wells, J.M.; Troise, A.D.; Capuano, E.; Fogliano, V. Polyphenols and Tryptophan Metabolites Activate the Aryl Hydrocarbon Receptor in an in vitro Model of Colonic Fermentation. Mol. Nutr. Food Res. 2019, 63, 1800722. [Google Scholar] [CrossRef]
- Van Dorsten, F.A.; Peters, S.; Gross, G.; Gomez-Roldan, V.; Klinkenberg, M.; De Vos, R.C.; Vaughan, E.E.; Van Duynhoven, J.P.; Possemiers, S.; Van De Wiele, T.; et al. Gut microbial metabolism of polyphenols from black tea and red wine/grape juice is source-specific and colon-region dependent. J. Agric. Food Chem. 2012, 60, 11331–11342. [Google Scholar] [CrossRef] [PubMed]
- Barroso, E.; Van De Wiele, T.; Jiménez-Girón, A.; Muñoz-González, I.; Martín-Alvarez, P.J.; Moreno-Arribas, M.V.; Bartolomé, B.; Peláez, C.; Martínez-Cuesta, M.C.; Requena, T. Lactobacillus plantarum IFPL935 impacts colonic metabolism in a simulator of the human gut microbiota during feeding with red wine polyphenols. Appl. Microbiol. Biotechnol. 2014, 98, 6805–6815. [Google Scholar] [CrossRef] [Green Version]
- Attri, S.; Goel, G. Influence of polyphenol rich seabuckthorn berries juice on release of polyphenols and colonic microbiota on exposure to simulated human digestion model. Food Res. Int. 2018, 111, 314–323. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Duysburgh, C.; Jiang, T.A.; Rebaza, M.; Pinheiro, I.; Marzorati, M. A combination of xylooligosaccharides and a polyphenol blend affect microbial composition and activity in the distal colon exerting immunomodulating properties on human cells. J. Funct. Foods 2018, 47, 163–171. [Google Scholar] [CrossRef]
- Van Rymenant, E.; Salden, B.; Voorspoels, S.; Jacobs, G.; Noten, B.; Pitart, J.; Possemiers, S.; Smagghe, G.; Grootaert, C.; Van Camp, J. A Critical Evaluation of In Vitro Hesperidin 2S Bioavailability in a Model Combining Luminal (Microbial) Digestion and Caco-2 Cell Absorption in Comparison to a Randomized Controlled Human Trial. Mol. Nutr. Food Res. 2018, 62, 1700881. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Sun, J.; Liang, Z.; Xu, H.; Du, P.; Li, A.; Meng, Y.; Reshetnik, E.I.; Liu, L.; Li, C. The bioavailability of soy isoflavones in vitro and their effects on gut microbiota in the simulator of the human intestinal microbial ecosystem. Food Res. Int. 2022, 152, 110868. [Google Scholar] [CrossRef]
- Wu, T.; Grootaert, C.; Voorspoels, S.; Jacobs, G.; Pitart, J.; Kamiloglu, S.; Possemiers, S.; Heinonen, M.; Kardum, N.; Glibetic, M.; et al. Aronia (Aronia melanocarpa) phenolics bioavailability in a combined in vitro digestion/Caco-2 cell model is structure and colon region dependent. J. Funct. Foods 2017, 38, 128–139. [Google Scholar] [CrossRef]
- Wu, T.; Grootaert, C.; Pitart, J.; Vidovic, N.K.; Kamiloglu, S.; Possemiers, S.; Glibetic, M.; Smagghe, G.; Raes, K.; Van de Wiele, T.; et al. Aronia (Aronia melanocarpa) Polyphenols Modulate the Microbial Community in a Simulator of the Human Intestinal Microbial Ecosystem (SHIME) and Decrease Secretion of Proinflammatory Markers in a Caco-2/endothelial Cell Coculture Model. Mol. Nutr. Food Res. 2018, 62, 1800607. [Google Scholar] [CrossRef] [PubMed]
- Freire, F.C.; Adorno, M.A.T.; Sakamoto, I.K.; Antoniassi, R.; Chaves, A.C.S.D.; dos Santos, K.M.O.; Sivieri, K. Impact of multi-functional fermented goat milk beverage on gut microbiota in a dynamic colon model. Food Res. Int. 2017, 99, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Guri, A.; Haratifar, S.; Corredig, M. Bioefficacy of Tea Catechins Associated with Milk Caseins Tested Using Different In Vitro Digestion Models. Food Dig. 2014, 5, 8–18. [Google Scholar] [CrossRef]
- Giuliani, C.; Marzorati, M.; Daghio, M.; Franzetti, A.; Innocenti, M.; Van de Wiele, T.; Mulinacci, N. Effects of Olive and Pomegranate By-Products on Human Microbiota: A Study Using the SHIME® In Vitro Simulator. Molecules 2019, 24, 3791. [Google Scholar] [CrossRef] [Green Version]
- Khoo, C.; Duysburgh, C.; Marzorati, M.; Van den Abbeele, P.; Zhang, D. A Freeze-Dried Cranberry Powder Consistently Enhances SCFA Production and Lowers Abundance of Opportunistic Pathogens In Vitro. BioTech 2022, 11, 14. [Google Scholar] [CrossRef]
- Cueva, C.; Jiménez-Girón, A.; Muñoz-González, I.; Esteban-Fernández, A.; Gil-Sánchez, I.; Dueñas, M.; Martín-Álvarez, P.J.; Pozo-Bayón, M.A.; Bartolomé, B.; Moreno-Arribas, M.V. Application of a new Dynamic Gastrointestinal Simulator (SIMGI) to study the impact of red wine in colonic metabolism. Food Res. Int. 2015, 72, 149–159. [Google Scholar] [CrossRef]
- Gil-Sánchez, I.; Cueva, C.; Sanz-Buenhombre, M.; Guadarrama, A.; Moreno-Arribas, M.V.; Bartolomé, B. Dynamic gastrointestinal digestion of grape pomace extracts: Bioaccessible phenolic metabolites and impact on human gut microbiota. J. Food Compost. Anal. 2018, 68, 41–52. [Google Scholar] [CrossRef]
- Fernández-Jalao, I.; Balderas, C.; Calvo, M.V.; Fontecha, J.; Sánchez-Moreno, C.; De Ancos, B. Impact of High-Pressure Processed Onion on Colonic Metabolism Using a Dynamic Gastrointestinal Digestion Simulator. Metabolites 2021, 11, 262. [Google Scholar] [CrossRef]
- Juániz, I.; Ludwig, I.A.; Bresciani, L.; Dall’Asta, M.; Mena, P.; Del Rio, D.; Cid, C.; de Peña, M.P. Catabolism of raw and cooked green pepper (Capsicum annuum) (poly)phenolic compounds after simulated gastrointestinal digestion and faecal fermentation. J. Funct. Foods 2016, 27, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Papillo, V.A.; Arlorio, M.; Locatelli, M.; Fuso, L.; Pellegrini, N.; Fogliano, V. In vitro evaluation of gastro-intestinal digestion and colonic biotransformation of curcuminoids considering different formulations and food matrices. J. Funct. Foods 2019, 59, 156–163. [Google Scholar] [CrossRef]
- Sáyago-Ayerdi, S.G.; Venema, K.; Tabernero, M.; Sarriá, B.; Bravo, L.; Mateos, R. Bioconversion of polyphenols and organic acids by gut microbiota of predigested Hibiscus sabdariffa L. calyces and Agave (A. tequilana Weber) fructans assessed in a dynamic in vitro model (TIM-2) of the human colon. Food Res. Int. 2021, 143, 110301. [Google Scholar] [CrossRef]
- Taladrid, D.; de Llano, D.G.; Zorraquín-Peña, I.; Tamargo, A.; Silva, M.; Molinero, N.; Moreno-Arribas, M.V.; Bartolomé, B. Gastrointestinal Digestion of a Grape Pomace Extract: Impact on Intestinal Barrier Permeability and Interaction with Gut Microbiome. Nutrients 2021, 13, 2467. [Google Scholar] [CrossRef] [PubMed]
- Zorraquín-peña, I.; Taladrid, D.; Tamargo, A.; Silva, M.; Molinero, N.; de Llano, D.G.; Bartolomé, B.; Moreno-arribas, M.V. Effects of Wine and Its Microbial-Derived Metabolites on Intestinal Permeability Using Simulated Gastrointestinal Digestion/Colonic Fermentation and Caco-2 Intestinal Cell Models. Microorganisms 2021, 9, 1378. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Cueva, C.; Alba, C.; Rodriguez, J.M.; de Pascual-Teresa, S.; Jones, J.; Caturla, N.; Victoria Moreno-Arribas, M.; Bartolomé, B. Gut microbiome-modulating properties of a polyphenol-enriched dietary supplement comprised of hibiscus and lemon verbena extracts. Monitoring of phenolic metabolites. J. Funct. Foods 2022, 91, 105016. [Google Scholar] [CrossRef]
- Tamargo, A.; Cueva, C.; Silva, M.; Molinero, N.; Miralles, B.; Bartolomé, B.; Moreno-Arribas, M.V. Gastrointestinal co-digestion of wine polyphenols with glucose/whey proteins affects their bioaccessibility and impact on colonic microbiota. Food Res. Int. 2022, 155, 111010. [Google Scholar] [CrossRef] [PubMed]
- Anson, N.M.; Selinheimo, E.; Havenaar, R.; Aura, A.M.; Mattila, I.; Lehtinen, P.; Bast, A.; Poutanen, K.; Haenen, G.R.M.M. Bioprocessing of wheat bran improves in vitro bioaccessibility and colonic metabolism of phenolic compounds. J. Agric. Food Chem. 2009, 57, 6148–6155. [Google Scholar] [CrossRef]
- Bianchi, F.; Lopes, N.P.; Adorno, M.A.T.; Sakamoto, I.K.; Genovese, M.I.; Saad, S.M.I.; Sivieri, K. Impact of combining acerola by-product with a probiotic strain on a gut microbiome model. Int. J. Food Sci. Nutr. 2018, 70, 182–194. [Google Scholar] [CrossRef] [Green Version]
- Tamargo, A.; Cueva, C.; Taladrid, D.; Khoo, C.; Moreno-Arribas, M.V.; Bartolomé, B.; González de Llano, D. Simulated gastrointestinal digestion of cranberry polyphenols under dynamic conditions. Impact on antiadhesive activity against uropathogenic bacteria. Food Chem. 2022, 368, 130871. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Morales, V.; Garcia-Llatas, G.; Yebra, M.J.; Sentandreu, V.; Alegría, A. In vitro colonic fermentation of a plant sterol-enriched beverage in a dynamic-colonic gastrointestinal digester. LWT 2021, 145, 111273. [Google Scholar] [CrossRef]
- Venema, K. The TNO In Vitro Model of the Colon (TIM-2). In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer: Cham, Switzerland; New York, NY, USA, 2015; pp. 293–304. [Google Scholar] [CrossRef] [Green Version]
- Sáyago-Ayerdi, S.G.; Zamora-Gasga, V.M.; Venema, K. Prebiotic effect of predigested mango peel on gut microbiota assessed in a dynamic in vitro model of the human colon (TIM-2). Food Res. Int. 2019, 118, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Sáyago-Ayerdi, S.G.; Venema, K.; Tabernero, M.; Sarriá, B.; Bravo, L.L.; Mateos, R. Bioconversion by gut microbiota of predigested mango (Mangifera indica L) ‘Ataulfo’ peel polyphenols assessed in a dynamic (TIM-2) in vitro model of the human colon. Food Res. Int. 2021, 139, 109963. [Google Scholar] [CrossRef]
- Gao, K.; Xu, A.; Krul, C.; Venema, K.; Liu, Y.; Niu, Y.; Lu, J.; Bensoussan, L.; Seeram, N.P.; Heber, D.; et al. Of the Major Phenolic Acids Formed during Human Microbial Fermentation of Tea, Citrus, and Soy Flavonoid Supplements, Only 3,4-Dihydroxyphenylacetic Acid Has Antiproliferative Activity. J. Nutr. 2006, 136, 52–57. [Google Scholar] [CrossRef]
- Sost, M.M.; Ahles, S.; Verhoeven, J.; Verbruggen, S.; Stevens, Y.; Venema, K. A Citrus Fruit Extract High in Polyphenols Beneficially Modulates the Gut Microbiota of Healthy Human Volunteers in a Validated In Vitro Model of the Colon. Nutrients 2021, 13, 3915. [Google Scholar] [CrossRef]
- Possemiers, S.; Verthé, K.; Uyttendaele, S.; Verstraete, W. PCR-DGGE-based quantification of stability of the microbial community in a simulator of the human intestinal microbial ecosystem. FEMS Microbiol. 2004, 49, 495–507. [Google Scholar] [CrossRef]
- Duque, A.L.R.F.; Monteiro, M.; Adorno, M.A.T.; Sakamoto, I.K.; Sivieri, K. An exploratory study on the influence of orange juice on gut microbiota using a dynamic colonic model. Food Res. Int. 2016, 84, 160–169. [Google Scholar] [CrossRef]
- De Boever, P.; Deplancke, B.; Verstraete, W. Fermentation by Gut Microbiota Cultured in a Simulator of the Human Intestinal Microbial Ecosystem Is Improved by Supplementing a Soygerm Powder. J. Nutr. 2000, 130, 2599–2606. [Google Scholar] [CrossRef] [Green Version]
- Venema, K.; van Nuenen, M.; Smeets-Peeters, M.; Minekus, M.; Havenaar, R. TNO’s in vitro large intestinal model: An excellent screening tool for functional food and pharmaceutical research. Ernährung 2000, 24, 558–564. [Google Scholar]
- Cárdenas-Castro, A.P.; Venema, K.; Sarriá, B.; Bravo, L.; Sáyago-Ayerdi, S.G.; Mateos, R. Study of the impact of a dynamic in vitro model of the colon (TIM-2) in the phenolic composition of two Mexican sauces. Food Res. Int. 2021, 139, 109917. [Google Scholar] [CrossRef]
- Minekus, M.; Smeets-Peeters, M.; Havenaar, R.; Bernalier, A.; Fonty, G.; Marol-Bonnin, S.; Alric, M.; Marteau, P.; Huis In’t Veld, J.H.J. A computer-controlled system to simulate conditions of the large intestine with peristaltic mixing, water absorption and absorption of fermentation products. Appl. Microbiol. Biotechnol. 1999, 53, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Maathuis, A.J.H.; van den Heuvel, E.G.; Schoterman, M.H.C.; Venema, K. Galacto-Oligosaccharides Have Prebiotic Activity in a Dynamic In Vitro Colon Model Using a 13C-Labeling Technique. J. Nutr. 2012, 142, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Cuevas-Tena, M.; Alegria, A.; Lagarda, M.J.; Venema, K. Impact of plant sterols enrichment dose on gut microbiota from lean and obese subjects using TIM-2 in vitro fermentation model. J. Funct. Foods 2019, 54, 164–174. [Google Scholar] [CrossRef]
- Elashoff, J.D.; Reedy, T.J.; Meyer, J.H. Analysis of Gastric Emptying Data. Gastroenterology 1982, 83, 1306–1312. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Macfarlane, S.; Gibson, G.R. Validation of a three-stage compound continuous culture system for investigating the effect of retention time on the ecology and metabolism of bacteria in the human colon. Microb. Ecol. 1998, 35, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Morales, V.; Garcia-Llatas, G.; Yebra, M.J.; Sentandreu, V.; Lagarda, M.J.; Alegría, A. Impact of a Plant Sterol- And Galactooligosaccharide-Enriched Beverage on Colonic Metabolism and Gut Microbiota Composition Using an in Vitro Dynamic Model. J. Agric. Food Chem. 2020, 68, 1884–1895. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, M.; Ramiro-Garcia, J.; Koenen, M.E.; Venema, K. To pool or not to pool? Impact of the use of individual and pooled fecal samples for in vitro fermentation studies. J. Microbiol. Methods 2014, 107, 1–7. [Google Scholar] [CrossRef]
- Ribnicky, D.M.; Roopchand, D.E.; Oren, A.; Grace, M.; Poulev, A.; Lila, M.A.; Havenaar, R.; Raskin, I. Effects of a high fat meal matrix and protein complexation on the bioaccessibility of blueberry anthocyanins using the TNO gastrointestinal model (TIM-1). Food Chem. 2014, 142, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Ribnicky, D.M.; Roopchand, D.E.; Poulev, A.; Kuhn, P.; Oren, A.; Cefalu, W.T.; Raskin, I. Artemisia dracunculus L. polyphenols complexed to soy protein show enhanced bioavailability and hypoglycemic activity in C57BL/6 mice. Nutrition 2014, 30, S4–S10. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Patán, F.; Cueva, C.; Monagas, M.; Walton, G.E.; Gibson, G.R.; Quintanilla-López, J.E.; Lebrón-Aguilar, R.; Martín-Álvarez, P.J.; Moreno-Arribas, M.V.; Bartolomé, B. In vitro fermentation of a red wine extract by human gut microbiota: Changes in microbial groups and formation of phenolic metabolites. J. Agric. Food Chem. 2012, 60, 2136–2147. [Google Scholar] [CrossRef]
- Sánchez-Patán, F.; Barroso, E.; Van De Wiele, T.; Jiménez-Girón, A.; Martín-Alvarez, P.J.; Moreno-Arribas, M.V.; Martínez-Cuesta, M.C.; Peláez, C.; Requena, T.; Bartolomé, B. Comparative in vitro fermentations of cranberry and grape seed polyphenols with colonic microbiota. Food Chem. 2015, 183, 273–282. [Google Scholar] [CrossRef]
- Barroso, E.; Sánchez-Patán, F.; Martín-Alvarez, P.J.; Bartolomé, B.; Moreno-Arribas, M.V.; Peláez, C.; Requena, T.; Van De Wiele, T.; Martínez-Cuesta, M.C. Lactobacillus plantarum IFPL935 favors the initial metabolism of red wine polyphenols when added to a colonic microbiota. J. Agric. Food Chem. 2013, 61, 10163–10172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cárdenas-Castro, A.P.; Bianchi, F.; Tallarico-Adorno, M.A.; Montalvo-González, E.; Sáyago-Ayerdi, S.G.; Sivieri, K. In vitro colonic fermentation of Mexican “taco” from corn-tortilla and black beans in a Simulator of Human Microbial Ecosystem (SHIME®) system. Food Res. Int. 2019, 118, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Sáyago-Ayerdi, S.G.; Zamora-Gasga, V.M.; Venema, K. Changes in gut microbiota in predigested Hibiscus sabdariffa L. calyces and Agave (Agave tequilana weber) fructans assessed in a dynamic in vitro model (TIM-2) of the human colon. Food Res Int. 2020, 132, 109036. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hémon, B.; Moskal, A.; Overvad, K.; Tjønneland, A.; Kyrø, C.; Fagherazzi, G.; Boutron-Ruault, M.C.; et al. Dietary polyphenol intake in Europe: The European Prospective Investigation into Cancer and Nutrition (EPIC) study. Eur. J. Nutr. 2016, 55, 1359. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M. Dietary Intake of Natural Antioxidants: Vitamins and Polyphenols. Crit. Rev. Food Sci. Nutr. 2013, 53, 706–721. [Google Scholar] [CrossRef]
- Kardum, N.; Konic Ristic, A.; Zec, M.; Kojadinovic, M.; Petrovic-Oggiano, G.; Zekovic, M.; Kroon, P.A.; Glibetić, M. Design, formulation and sensory evaluation of a polyphenol-rich food placebo: An example of aronia juice for food intervention studies. Int. J. Food Sci. Nutr. 2017, 68, 742–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Sivaprakasapillai, B.; Edirisinghe, I.; Randolph, J.; Steinberg, F.; Kappagoda, T. Effect of grape seed extract on blood pressure in subjects with the metabolic syndrome. Metab. Clin. Exp. 2009, 58, 1743–1746. [Google Scholar] [CrossRef]
- Van der Stelt, I.; Hoek-van den Hil, E.F.; Swarts, H.J.M.; Vervoort, J.J.M.; Hoving, L.; Skaltsounis, L.; Lemonakis, N.; Andreadou, I.; van Schothorst, E.M.; Keijer, J. Nutraceutical oleuropein supplementation prevents high fat diet-induced adiposity in mice. J. Funct. Foods 2015, 14, 702–715. [Google Scholar] [CrossRef]
- Olmez, E.; Vural, K.; Gok, S.; Ozturk, Z.; Kayalar, H.; Ayhan, S.; Var, A. Olive Leaf Extract Improves the Atherogenic Lipid Profile in Rats Fed a High Cholesterol Diet. Phytother. Res. 2015, 29, 1652–1657. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Cornelius, C.; Trovato-Salinaro, A.; Cambria, M.; Locascio, M.; Rienzo, L.; Condorelli, D.; Mancuso, C.; De Lorenzo, A.; Calabrese, E. The hormetic role of dietary antioxidants in free radical-related diseases. Curr. Pharm. Des. 2010, 16, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Cladis, D.P.; Weaver, C.M.; Ferruzzi, M.G. (Poly)phenol toxicity in vivo following oral administration: A targeted narrative review of (poly)phenols from green tea, grape, and anthocyanin-rich extracts. Phytother. Res. 2022, 36, 323–335. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Refined exposure assessment for curcumin (E-100). EFSA J. 2014, 12, 3876. [Google Scholar] [CrossRef]
- De Weirdt, R.; Possemiers, S.; Vermeulen, G.; Moerdijk-Poortvliet, T.C.; Boschker, H.T.; Verstraete, W.; Van de Wiele, T. Human faecal microbiota display variable patterns of glycerol metabolism. FEMS Microbiol. Ecol. 2010, 72, 354–369. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, A.E.; Clesceri, L.S.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater, 18th ed.; American Public Health Association, American Water Works Association and Water Environment, Federation in Washington: Washington, DC, USA, 1992. [Google Scholar]
- Trifirò, A.; Saccani, G.; Gherardi, S.; Vicini, E.; Spotti, E.; Previdi, M.P.; Ndagijimana, M.; Cavalli, S.; Reschiotto, C. Use of ion chromatography for monitoring microbial spoilage in the fruit juice industry. J. Chromatogr. A 1997, 770, 243–252. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, Y.; Dai, X.; Wei, W. Monitoring of Lactobacillus fermentation process by using ion chromatography with a series piezoelectric quartz crystal detector. J. Microbiol. Methods 2001, 44, 105–111. [Google Scholar] [CrossRef]
- Wojtczak, M.; Antczak, A.; Przybyt, M. Use of ionic chromatography in determining the contamination of apple juice by lactic acid. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess 2010, 27, 817–824. [Google Scholar] [CrossRef]
- Gil-Sánchez, I.; Cueva, C.; Tamargo, A.; Quintela, J.C.; de la Fuente, E.; Walker, A.W.; Moreno-Arribas, M.V.; Bartolomé, B. Application of the dynamic gastrointestinal simulator (simgi®) to assess the impact of probiotic supplementation in the metabolism of grape polyphenols. Food Res. Int. 2020, 129, 108790. [Google Scholar] [CrossRef]
- Sun, S.; Huang, S.; Shi, Y.; Shao, Y.; Qiu, J.; Sedjoah, R.C.A.A.; Yan, Z.; Ding, L.; Zou, D.; Xin, Z. Extraction, isolation, characterization and antimicrobial activities of non-extractable polyphenols from pomegranate peel. Food Chem. 2021, 351, 129232. [Google Scholar] [CrossRef] [PubMed]
- Reynoso-Camacho, R.; Sotelo-González, A.M.; Patiño-Ortiz, P.; Rocha-Guzmán, N.E.; Pérez-Ramírez, I.F. Berry by-products obtained from a decoction process are a rich source of low- and high-molecular weight extractable and non-extractable polyphenols. Food Bioprod. Process. 2021, 127, 371–387. [Google Scholar] [CrossRef]
- Pellegrini, N.; Yang, R.M.; Rice-Evans, C. Screening of Dietary Carotenoids and Carot-enoid-Rich Fruit Extracts for Antioxidant Activities Applying 2,2’-azinobis (3-ethylenebenzothiazoline-6-sulfonic acid) Radical Cation Decolorization Assay. Methods Enzymol. 1999, 299, 379–384. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Feed Components | Concentration (g/L) | References |
|---|---|---|
| Arabinogalactan | 1.0 | [71,72,73,75,76,81,102,103] |
| 1.2 | [39] | |
| Pectin | 2.0 | [71,72,73,75,76,81,102,103] |
| Xylan | 1.0 | [71,72,73,75,76,81,102,103] |
| 0.5 | [70] | |
| Starch | 4.0 | [70,75,76,80,81] |
| 3.0 | [71,72,73,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102] | |
| Glucose | 0.4 | [70,71,72,73,75,76,81,102,103] |
| Yeast Extract | 3.0 | [70,71,72,73,75,76,81,102,103] |
| Peptone | 1.0 | [70,73,76,77,81,102,103] |
| 3.0 | [69,74,75,80] | |
| Mucin | 4.0 | [71,72,73,76,81,102,103] |
| 1.0 | [69,74,75,80] | |
| 0.3 | [70] | |
| L-Cysteine Hydrochloride | 0.5 | [70,71,72,73,75,76,81,102,103] |
| Components of the Feed | g/L | |||
|---|---|---|---|---|
| Pectine | 12 a | 9.4 b | 9 c | 4.7 d |
| Xylan | 12 a | 9.4 b | 9 c | 4.7 d |
| Arabinogalactan | 12 a | 9.4 b | 9 c | 4.7 d |
| Amylopectin | 12 a | 9.4 b | 9 c | 4.7 d |
| Starch | 100 a | 78.4 b | 74.6 c | 39.2 d |
| Tween 80 | 270 a | 34.0 b | 31.5 c | 17 d |
| Bactopepton | 375 a | 47.0 b | 43.7 c | 23.5 d |
| Casein | 375 a | 47.0 b | 43.7 c | 23.5 d |
| Ox-bile | 6.25 a | 0.8 b | 0.7 c | 0.4 d |
| MgSO4 | 50 a | 0.5 b | 0.7 c | - |
| L-Cysteine | 20 a | 0.4 b | 0.3 c | - |
| Vitamin mixture | - | 1 mL b | 1.5 mL c | - |
| K2HPO4·3H2O | 4.7 a | 2.5 b | 4.7 c | - |
| NaCl | 8.4 a | 4.5 b | 8.4 c | - |
| CaCl2·2H2O | 0.8 a | 0.45 b | 0.8 c | - |
| FeSO4·7H2O | 0.009 a | 0.005 b | 0.009 c | - |
| Haemin | 0.02 a | - | 0.02 c | - |
| Feed Components | Concentration (g/L) | References | Colon Nutrient Medium (CNM) | Concentration (g/L) | References |
|---|---|---|---|---|---|
| Arabinogalactan | 1 | [83,84,92] | Yeast Extract | 2 | [91] |
| Pectin (from apple) | 2 | Peptone | 2 | ||
| Xylan | 1 | L-cysteine | 0.5 | ||
| Potato Starch | 3 | NaCl | 0.1 | ||
| Glucose | 0.4 | K2HPO4 | 0.04 | ||
| Yeast Extract | 3 | KH2PO4 | 0.04 | ||
| Peptone | 1 | MgSO4·7H2O | 0.01 | ||
| Mucin | 4 | CaCl2·6H2O | 0.01 | ||
| L-cysteine | 0.5 | NaHCO3 | 2 | ||
| Tween 80 | 2 mL/L | ||||
| Hemin | 0.05 | ||||
| Vitamin K | 10 µL/L | ||||
| Bile salts | 0.5 |
| Reference | Dose | Selection | Fecal donors | Protocol source | |
|---|---|---|---|---|---|
| Single SHIME | [73] | 10% of juice/day | - | Pool (four donors) | - |
| [74] | 1597 mg/day | A commercial extract, which indicated composition | one donor | [102] | |
| [76] | 70 mg isoflavones/day | - | Pool (three donors) | - | |
| [77] | 100 mL/day juice with 6.5 g/L polyphenols | Daily recommended dose of Aronia Juice (Kardum, 2017) | one donor | [102] | |
| [78] | 100 mL/day juice with 6.5 g/L polyphenols | Daily recommended dose of Aronia Juice (Kardum, 2017) | one donor | [102] | |
| [79] | 200 mL formulation/day | - | Pool (three donors) | [102] | |
| [80] | 2.18 µM of polyphenols | - | - | - | |
| [94] | 2.5% (w/v) of by-product | As the minimum level established by the Brazilian Health Regulatory Agency for food to be considered as a source of fiber. | Pool (three donors) | - | |
| [119] | 50 g cooked beans and 50 g corn-tortilla | - | Pool (three donors) | [102] | |
| [118] | 500 mg extract/L | - | one donor | [102] | |
| [68] | 1 g extract | Content present in red wine and grape juice extract and tea extract from Lipton’s black tea | one donor | [102] | |
| TWIN- SHIME | [69] | 1.8 g/day | - | one donor per metabolotype | [102] |
| [70] | 1.5 g oregano, 9.7 mg luteolin/100 g | - | Pool (three donors) | [102] | |
| [71] | 1 g extract | Polyphenol doses selected to simulate physiological conditions since total intake for humans has been estimated at 1 g/day | one donor | [102] | |
| [72] | 200 mg extract/day; 200 mg extract/day + 1010 CFU Lactobacillus | It corresponds to daily polyphenol intake of 250 mL of red wine | one donor | [104] | |
| [75] | 500 mg of Cordiart (450 mg hesperidin) | - | one donor | - | |
| [87] | 5 mg of curcuminoids, 500 mg of microencapsulated, 250 mg of turmeric and 25 mg of Meriva®. | - | one donor | [70] | |
| [52] | 1 y 2 g extract/L | Taking into consideration previous human studies with stilbenes | one donor | [102] | |
| Triple- SHIME | [81] | 2–4 g/L of extract | - | one donor | [102] |
| [53] | 200 mg of (+)-Catechin | The dose represents a regular intake (+)-catechin in humans | Pool (12 donors) | [104] | |
| [82] | 7.4 g/L extract | - | Pool (five donors) | - | |
| [117] | 500 mg/L | - | - | [74] | |
| Other | [83] | 225 mL, corresponding to 405 mg of polyephnols | To simulate the intake of a glass of wine | 1 donor per experiment (two donors) | [104] |
| SIMGI | [84] | 700 mg (acute and chronic) | - | one donor per experiment (two donors) | [104] |
| [91] | 90 mg and 270 mg of extract | According to Sánchez-Patán, 2012; Cueva et al., 2013 | one donor per experiment (two donors) | - | |
| [89] | 1 g extract (47.96 mg GAE/g phenolics) | - | one donor | [92] | |
| [92] | 1 g of extract | - | one donor | [84] | |
| [95] | 240 mL wine and 80 mL of prepared food models. | Considering a daily dose of 240 mL of wine (Cueva, 2015, Muñoz Gonzalez, 2013) | one donor | [84] | |
| [90] | 80 mL wine (200 mg of gallic acid equivalents) | - | one donor | [92] | |
| [55] | 0.5 of extract with 100 g of meal matrix | - | - | - | |
| TIM-1 | [114] | 134.5 mg, 403.5 mg and 2 g of extract | - | - | - |
| [115] | 0.5 g of extract | - | - | - | |
| [55] | 20 mg rosmarinic acid | - | - | - | |
| [106] | 7.5 g/day, 2.5 mL/h two Mexican sauces | - | Pool (seven donors) | [78] | |
| TIM-2 | [100] | 600 mg extract | - | Pool (10 donors) | - |
| [73] | 250 y 350 mg extract/day | - | Pool (seven donors) | [113] | |
| [98] | 7.5 g/day, 2.5 mL/h pre-digested mango peel | - | Pool (three donors) | [105] | |
| [120] | 7.5 g/day, 2.5 mL/h pre-digested mango peel | - | Pool (three donors) | [105] | |
| [88] | - | - | Pool (three donors) | [105] | |
| [99] | 7.5 g/day, 2.5 mL/h pre-digested mango peel | - | Pool (three donors) | [105] |
| Target Group | Selective Culture Media | Incubation Time | Incubation Conditions | Reference |
|---|---|---|---|---|
| Total aerobes | Brain heart infusion (BHI) Agar | 24 | Aerobic | [68,73] |
| Standard Methods agar | 48 | Aerobic | [103] | |
| Trypticase Soy Agar (TSA) | 48 | Aerobic | [83,84,91,92,95] | |
| Total anaerobes | Brain heart infusion (BHI) Agar | 72 | Anaerobic | [68,72] |
| Standard Methods agar | 48 | Anaerobic | [103] | |
| Wilkins-Chalgren agar | 48 | Anaerobic | [83,84,91,92,95] | |
| Lactic acid bacteria | De Man, Rogosa and Sharpe (MRS) Agar | 48 | Aerobic | [72,73] |
| 48 | Anaerobic | [83,84,91,92,95,103] | ||
| Lactobacilli | LAMVAB Agar | 48 | Aerobic | [72] |
| 72 | Microaerophilic incubation | [68] | ||
| 48 | Anaerobic | [91,92,95] | ||
| Bifidobacterium spp. | Bifidobacteirum agar modified by Beerens (Difco., BD, USA) | 48 | Anaerobic | [91] |
| BIM-25 agar | 72 | Anaerobic | [103] | |
| Total Coliforms | MacConkey agar | 24 | Aerobic | [68,72,73] |
| Staphylococci | Mannitol Salt agar broth | 48 | Aerobic | [68,84,95] |
| Clostridium spp. | Reinforced Clostridial agar | 48 | Anaerobic | [103] |
| Tryptose Sulfite Cycloserine (TSC) | 48 | Anaerobic | [91,92,95] | |
| Enterobacteria | MacConkey agar | 48 | Anaerobic | [83,84,91,92,95] |
| Enterococcus spp. | KF Streptococcus agar | 48 | Aerobic | [103] |
| Enterococcus agar | 48 | Anaerobic | [84,91,92,95] | |
| Enterococcus agar | 48 | Aerobic | [68] | |
| Bartley and slanz agar | 48 | Aerobic | [73] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duque-Soto, C.; Quintriqueo-Cid, A.; Rueda-Robles, A.; Robert, P.; Borrás-Linares, I.; Lozano-Sánchez, J. Evaluation of Different Advanced Approaches to Simulation of Dynamic In Vitro Digestion of Polyphenols from Different Food Matrices—A Systematic Review. Antioxidants 2023, 12, 101. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12010101
Duque-Soto C, Quintriqueo-Cid A, Rueda-Robles A, Robert P, Borrás-Linares I, Lozano-Sánchez J. Evaluation of Different Advanced Approaches to Simulation of Dynamic In Vitro Digestion of Polyphenols from Different Food Matrices—A Systematic Review. Antioxidants. 2023; 12(1):101. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12010101
Chicago/Turabian StyleDuque-Soto, Carmen, Alejandra Quintriqueo-Cid, Ascensión Rueda-Robles, Paz Robert, Isabel Borrás-Linares, and Jesús Lozano-Sánchez. 2023. "Evaluation of Different Advanced Approaches to Simulation of Dynamic In Vitro Digestion of Polyphenols from Different Food Matrices—A Systematic Review" Antioxidants 12, no. 1: 101. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12010101