

A Novel Dual-Function Nitric Oxide Donor Therapy for Preeclampsia—A Proof-of-Principle Study in a Murine Model
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Vascular Function Studies
2.3. Animal Model and AKT-1005 Treatment
2.4. Urine Collections
2.5. Light and Electron Microscopy
2.6. Blood Pressure Measurement
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Statistical Analysis
3. Results
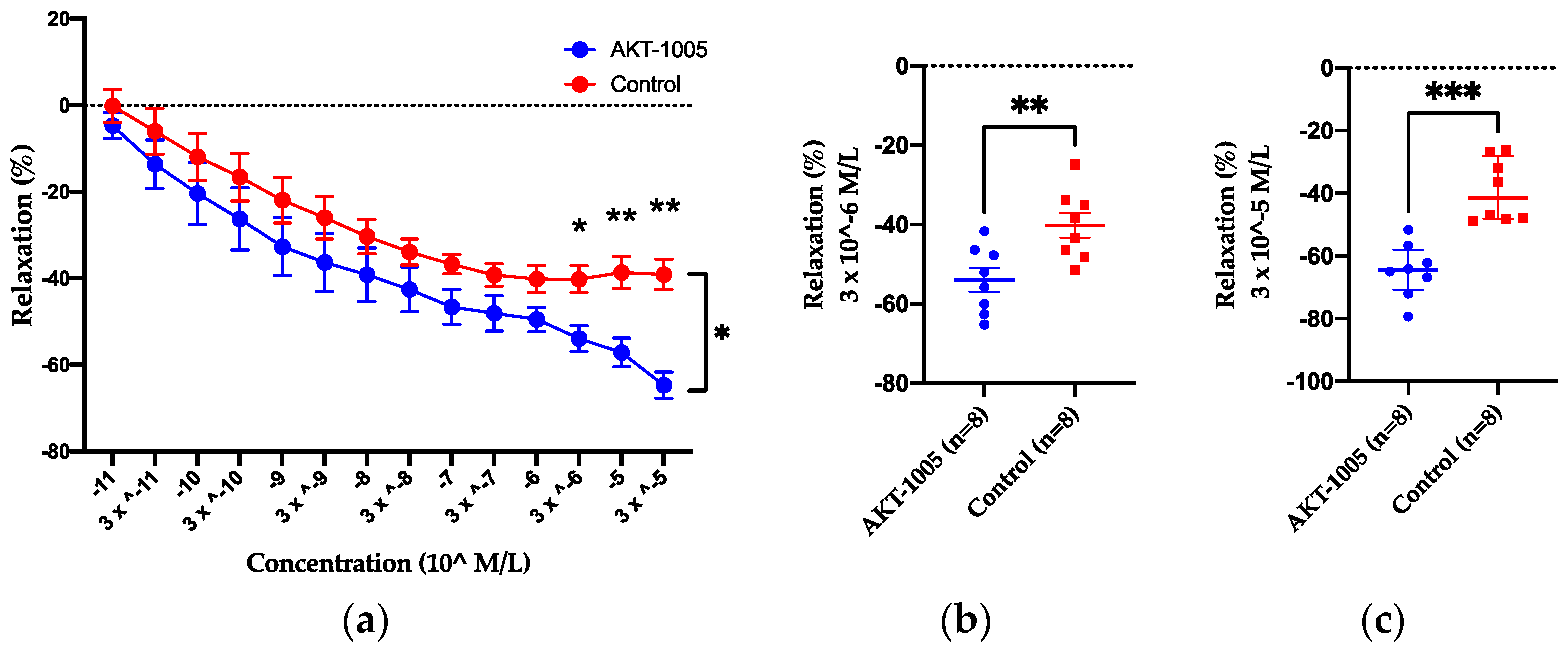
3.1. AKT-1005 Has Vasodilating Effect on Resistance Vessels Ex Vivo and Does Not Cause Hypotension in Control Mice
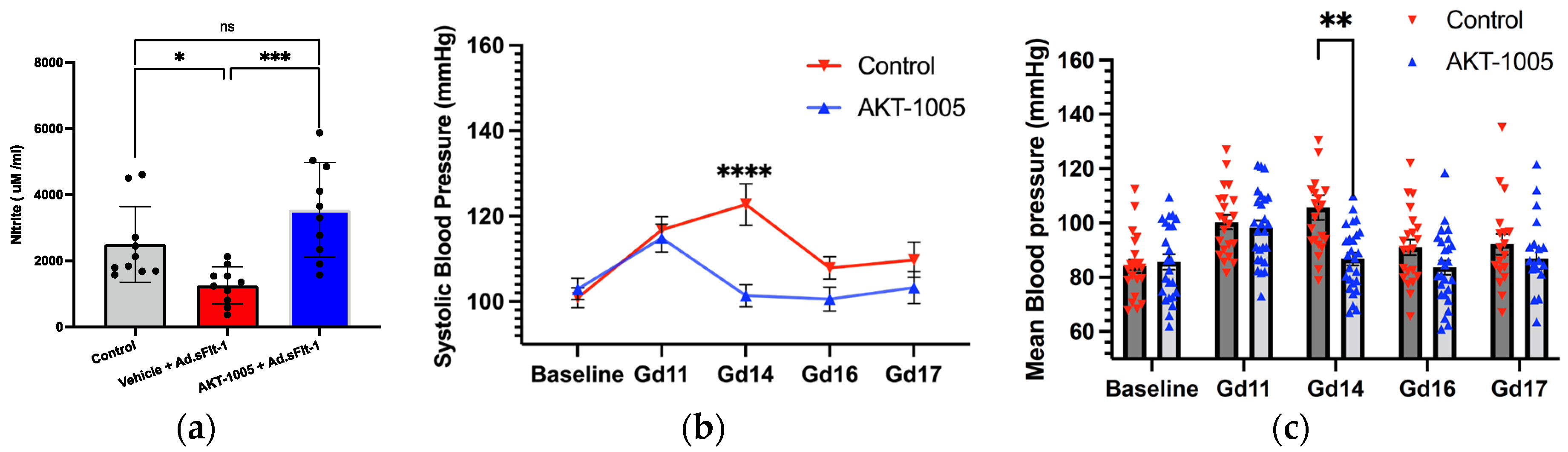
3.2. AKT-1005 Increases NO Bioavailability and Ameliorates Hypertension in Ad.sFlt-1-Transfected Mice
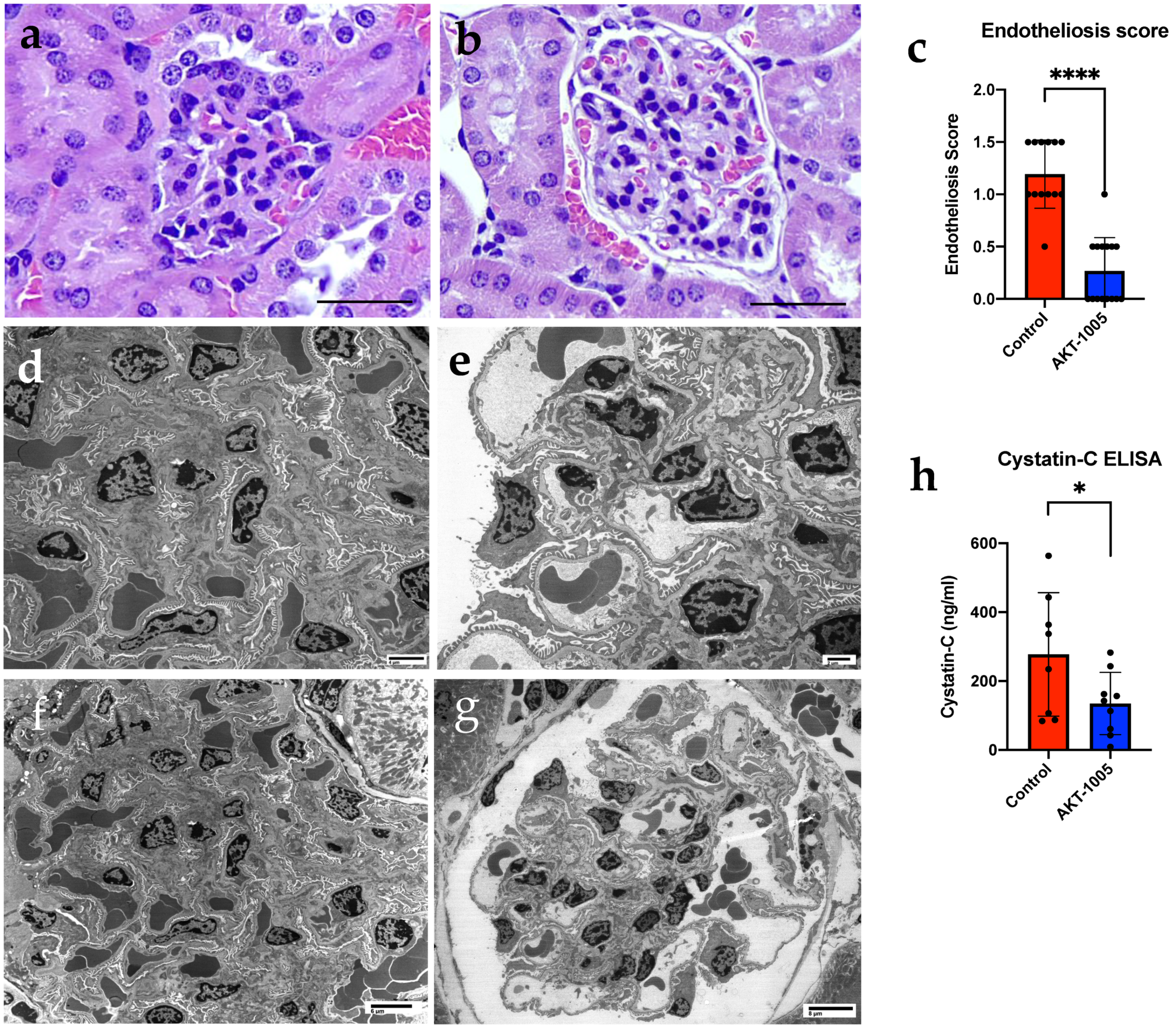
3.3. AKT-1005 Prevents Renal Glomerular Endotheliosis and Improves Kidney Function in Ad.sFlt-1-Transfected Mice
3.4. AKT-1005 Relieves Placental Stress in Ad.sFlt-1-Transfected Mice
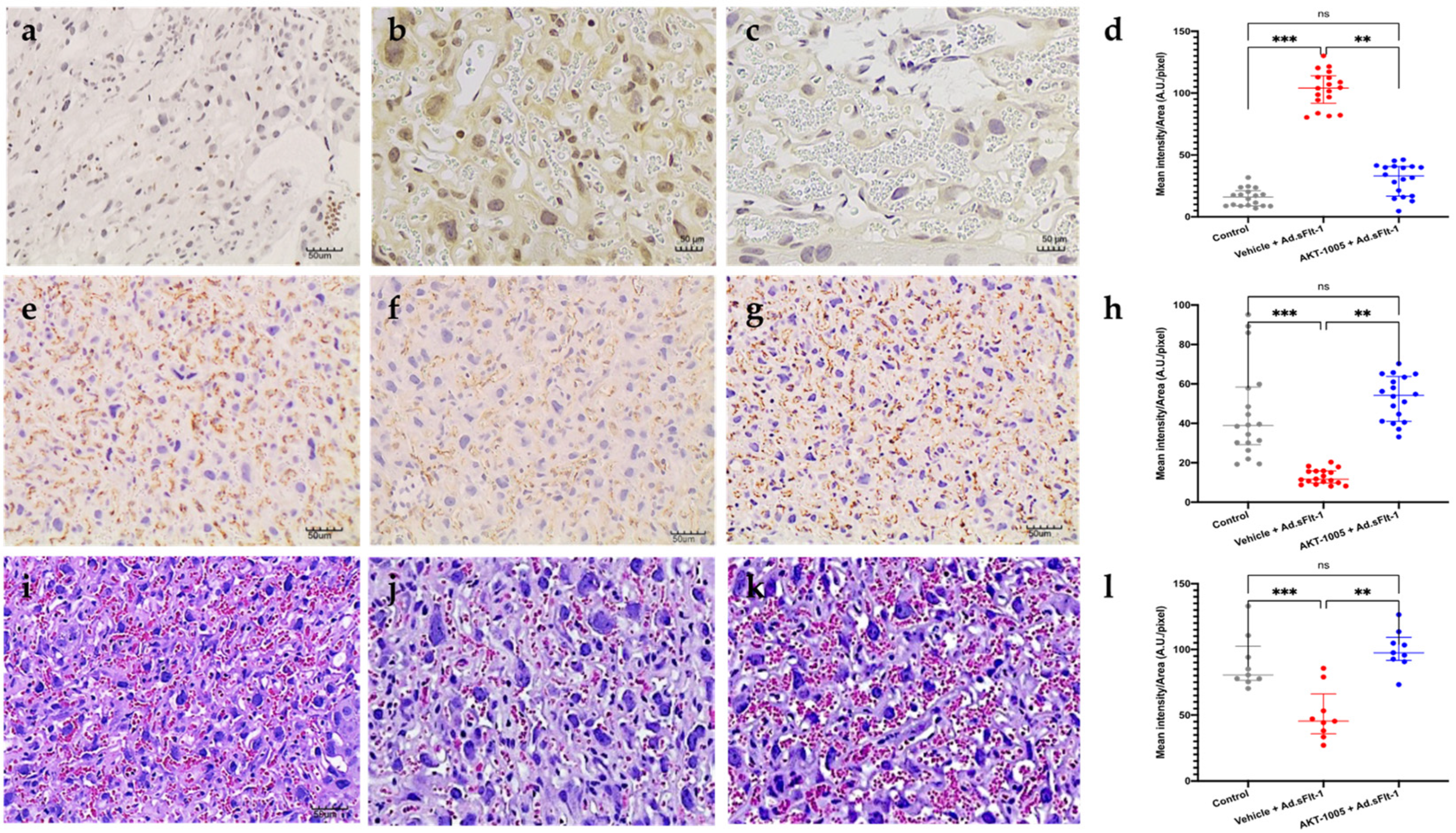
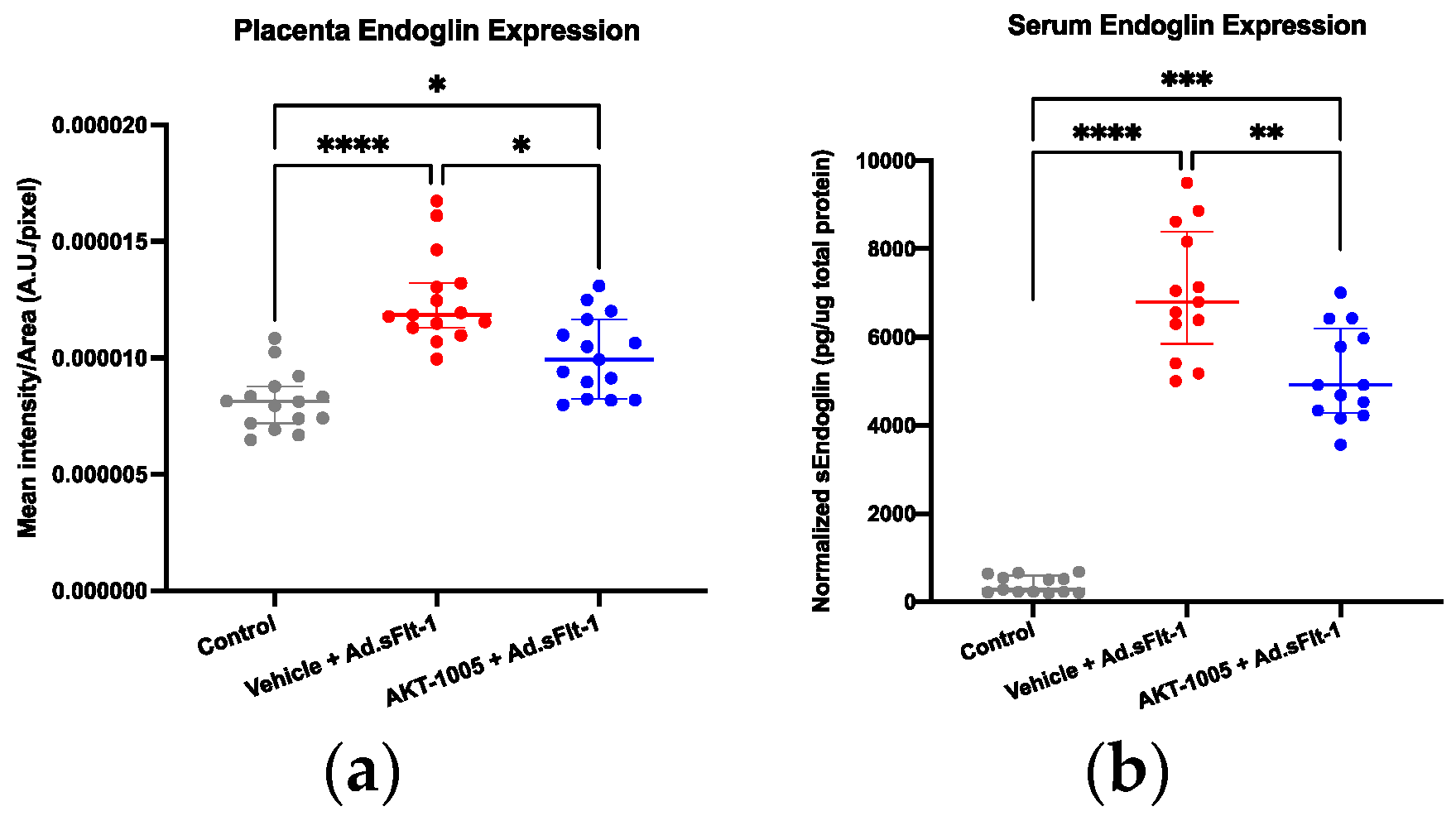
3.5. AKT-1005 Reduces Anti-Angiogenic Response in Ad.sFlt-1-Transfected Mice
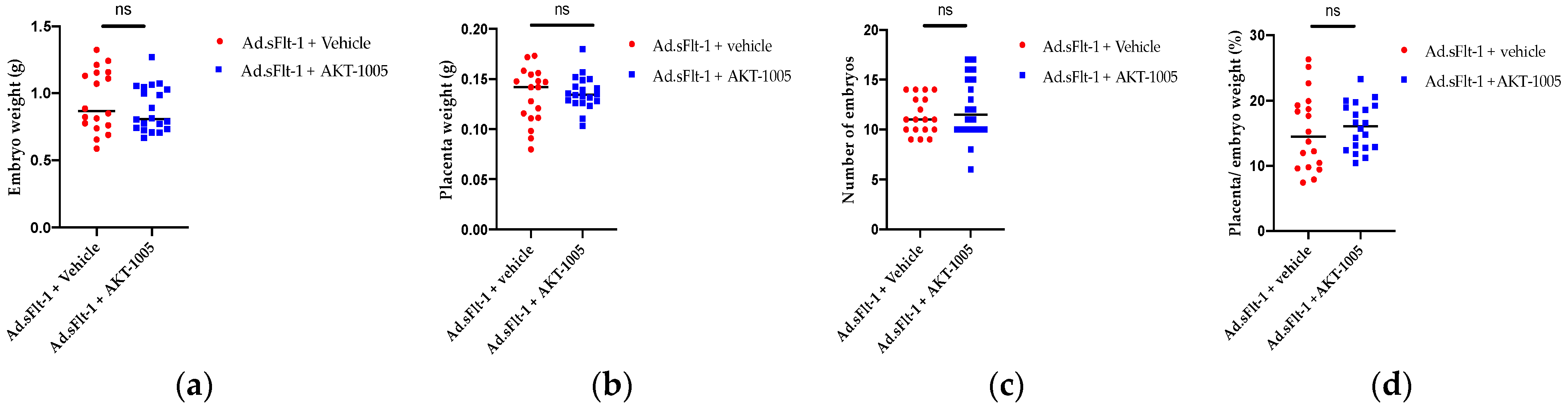
3.6. AKT-1005 Does Not Alter Pregnancy Outcomes in Ad.sFlt-1-Transfected Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American College of Obstetricians and Gynecologists. ACOG Practice Bulletin No. 202: Gestational Hypertension and Preeclampsia. Obstet. Gynecol. 2019, 133, e1–e25. [Google Scholar]
- MacKay, A.P.; Berg, C.J.; Atrash, H.K. Pregnancy-related mortality from preeclampsia and eclampsia. Obstet. Gynecol. 2001, 97, 533–538. [Google Scholar]
- Steegers, E.A.; von Dadelszen, P.; Duvekot, J.J.; Pijnenborg, R. Pre-eclampsia. Lancet 2010, 376, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.S.; Wojdyla, D.; Say, L.; Gülmezoglu, A.M.; Van Look, P.F. WHO analysis of causes of maternal death: A systematic review. Lancet 2006, 367, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Stevens, W.; Shih, T.; Incerti, D.; Ton, T.G.N.; Lee, H.C.; Peneva, D.; Macones, G.A.; Sibai, B.M.; Jena, A.B. Short-term costs of preeclampsia to the United States health care system. Am. J. Obstet. Gynecol. 2017, 217, 237–248.e216. [Google Scholar] [CrossRef] [PubMed]
- Maynard, S.E.; Min, J.Y.; Merchan, J.; Lim, K.H.; Li, J.; Mondal, S.; Libermann, T.A.; Morgan, J.P.; Sellke, F.W.; Stillman, I.E.; et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J. Clin. Investig. 2003, 111, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.J.; Maynard, S.E.; Qian, C.; Lim, K.H.; England, L.J.; Yu, K.F.; Schisterman, E.F.; Thadhani, R.; Sachs, B.P.; Epstein, F.H.; et al. Circulating angiogenic factors and the risk of preeclampsia. N. Engl. J. Med. 2004, 350, 672–683. [Google Scholar] [CrossRef]
- Levine, R.J.; Lam, C.; Qian, C.; Yu, K.F.; Maynard, S.E.; Sachs, B.P.; Sibai, B.M.; Epstein, F.H.; Romero, R.; Thadhani, R.; et al. Soluble endoglin and other circulating antiangiogenic factors in preeclampsia. N. Engl. J. Med. 2006, 355, 992–1005. [Google Scholar] [CrossRef]
- Rana, S.; Lemoine, E.; Granger, J.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef]
- Venkatesha, S.; Toporsian, M.; Lam, C.; Hanai, J.-I.; Mammoto, T.; Kim, Y.M.; Bdolah, Y.; Lim, K.-H.; Yuan, H.-T.; Libermann, T.A.; et al. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat. Med. 2006, 12, 642–649. [Google Scholar] [CrossRef]
- Covarrubias, A.E.; Lecarpentier, E.; Lo, A.; Salahuddin, S.; Gray, K.J.; Karumanchi, S.A.; Zsengellér, Z.K. AP39, a Modulator of Mitochondrial Bioenergetics, Reduces Antiangiogenic Response and Oxidative Stress in Hypoxia-Exposed Trophoblasts: Relevance for Preeclampsia Pathogenesis. Am. J. Pathol. 2019, 189, 104–114. [Google Scholar] [CrossRef]
- Wang, Y.; Walsh, S.W. Placental mitochondria as a source of oxidative stress in pre-eclampsia. Placenta 1998, 19, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Vaka, V.R.; McMaster, K.M.; Cunningham, M.W., Jr.; Ibrahim, T.; Hazlewood, R.; Usry, N.; Cornelius, D.C.; Amaral, L.M.; LaMarca, B. Role of Mitochondrial Dysfunction and Reactive Oxygen Species in Mediating Hypertension in the Reduced Uterine Perfusion Pressure Rat Model of Preeclampsia. Hypertension 2018, 72, 703–711. [Google Scholar] [CrossRef]
- Myatt, L.; Kossenjans, W.; Sahay, R.; Eis, A.; Brockman, D. Oxidative stress causes vascular dysfunction in the placenta. J. Matern. Fetal Med. 2000, 9, 79–82. [Google Scholar] [PubMed]
- Myatt, L.; Cui, X. Oxidative stress in the placenta. Histochem. Cell Biol. 2004, 122, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Myatt, L. Review: Reactive oxygen and nitrogen species and functional adaptation of the placenta. Placenta 2010, 31, S66–S69. [Google Scholar] [CrossRef]
- Marín, R.; Chiarello, D.I.; Abad, C.; Rojas, D.; Toledo, F.; Sobrevia, L. Oxidative stress and mitochondrial dysfunction in early-onset and late-onset preeclampsia. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165961. [Google Scholar] [CrossRef]
- Tanne, J.H. Pre-eclampsia: FDA approves blood test to identify pregnant women at risk. BMJ 2023, 382, 1594. [Google Scholar] [CrossRef]
- López-Jaramillo, P.; Arenas, W.D.; García, R.G.; Rincon, M.Y.; López, M. The role of the L-arginine-nitric oxide pathway in preeclampsia. Ther. Adv. Cardiovasc. Dis. 2008, 2, 261–275. [Google Scholar] [CrossRef]
- Holden, D.P.; Fickling, S.A.; Whitley, G.S.; Nussey, S.S. Plasma concentrations of asymmetric dimethylarginine, a natural inhibitor of nitric oxide synthase, in normal pregnancy and preeclampsia. Am. J. Obstet. Gynecol. 1998, 178, 551–556. [Google Scholar] [CrossRef]
- Orange, S.J.; Painter, D.; Horvath, J.; Yu, B.; Trent, R.; Hennessy, A. Placental endothelial nitric oxide synthase localization and expression in normal human pregnancy and pre-eclampsia. Clin. Exp. Pharmacol. Physiol. 2003, 30, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Conrad, K.P. Expression of endothelial nitric oxide synthase by extravillous trophoblast cells in the human placenta. Placenta 2000, 21, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Nanaev, A.; Chwalisz, K.; Frank, H.G.; Kohnen, G.; Hegele-Hartung, C.; Kaufmann, P. Physiological dilation of uteroplacental arteries in the guinea pig depends on nitric oxide synthase activity of extravillous trophoblast. Cell Tissue Res. 1995, 282, 407–421. [Google Scholar] [CrossRef]
- Morris, N.H.; Sooranna, S.R.; Learmont, J.G.; Poston, L.; Ramsey, B.; Pearson, J.D.; Steer, P.J. Nitric oxide synthase activities in placental tissue from normotensive, pre-eclamptic and growth retarded pregnancies. Br. J. Obstet. Gynaecol. 1995, 102, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Belkacemi, L.; Bainbridge, S.A.; Dickinson, M.A.; Smith, G.N.; Graham, C.H. Glyceryl trinitrate inhibits hypoxia/reoxygenation-induced apoptosis in the syncytiotrophoblast of the human placenta: Therapeutic implications for preeclampsia. Am. J. Pathol. 2007, 170, 909–920. [Google Scholar] [CrossRef]
- Barsoum, I.B.; Renaud, S.J.; Graham, C.H. Glyceryl trinitrate inhibits hypoxia-induced release of soluble fms-like tyrosine kinase-1 and endoglin from placental tissues. Am. J. Pathol. 2011, 178, 2888–2896. [Google Scholar] [CrossRef]
- Alexander, B.T.; Llinas, M.T.; Kruckeberg, W.C.; Granger, J.P. L-arginine attenuates hypertension in pregnant rats with reduced uterine perfusion pressure. Hypertension 2004, 43, 832–836. [Google Scholar] [CrossRef]
- Murphy, S.R.; LaMarca, B.; Cockrell, K.; Arany, M.; Granger, J.P. L-arginine supplementation abolishes the blood pressure and endothelin response to chronic increases in plasma sFlt-1 in pregnant rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R259–R263. [Google Scholar] [CrossRef]
- Pintye, D.; Sziva, R.E.; Mastyugin, M.; Young, B.C.; Jacas, S.; Török, M.; Salahuddin, S.; Jagtap, P.; Southan, G.J.; Zsengellér, Z.K. A Novel Dual-Function Redox Modulator Relieves Oxidative Stress and Anti-Angiogenic Response in Placental Villus Explant Exposed to Hypoxia-Relevance for Preeclampsia Therapy. Biology 2023, 12, 1229. [Google Scholar] [CrossRef]
- Biwer, L.A.; Carvajal, B.V.; Lu, Q.; Man, J.J.; Jaffe, I.Z. Mineralocorticoid and Estrogen Receptors in Endothelial Cells Coordinately Regulate Microvascular Function in Obese Female Mice. Hypertension 2021, 77, 2117–2126. [Google Scholar] [CrossRef]
- Good, M.E.; Eucker, S.A.; Li, J.; Bacon, H.M.; Lang, S.M.; Butcher, J.T.; Johnson, T.J.; Gaykema, R.P.; Patel, M.K.; Zuo, Z.; et al. Endothelial cell Pannexin1 modulates severity of ischemic stroke by regulating cerebral inflammation and myogenic tone. JCI Insight 2018, 3, e96272. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.D.; Zsengeller, Z.K.; Khankin, E.V.; Lo, A.S.; Rajakumar, A.; DuPont, J.J.; McCurley, A.; Moss, M.E.; Zhang, D.; Clark, C.D.; et al. Soluble fms-like tyrosine kinase 1 promotes angiotensin II sensitivity in preeclampsia. J. Clin. Investig. 2016, 126, 2561–2574. [Google Scholar] [CrossRef] [PubMed]
- Holwerda, K.M.; Burke, S.D.; Faas, M.M.; Zsengeller, Z.; Stillman, I.E.; Kang, P.M.; van Goor, H.; McCurley, A.; Jaffe, I.Z.; Karumanchi, S.A.; et al. Hydrogen sulfide attenuates sFlt1-induced hypertension and renal damage by upregulating vascular endothelial growth factor. J. Am. Soc. Nephrol. 2014, 25, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Zsengellér, Z.K.; Rosen, S. The Use of Cytochrome C Oxidase Enzyme Activity and Immunohistochemistry in Defining Mitochondrial Injury in Kidney Disease. J. Histochem. Cytochem. 2016, 64, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Dupont, V.; Berg, A.H.; Yamashita, M.; Huang, C.; Covarrubias, A.E.; Ali, S.; Stotland, A.; Van Eyk, J.E.; Jim, B.; Thadhani, R.; et al. Impaired renal reserve contributes to preeclampsia via the kynurenine and soluble fms-like tyrosine kinase 1 pathway. J. Clin. Investig. 2022, 132, e158346. [Google Scholar] [CrossRef]
- ACOG Committee on Obstetric Practice. ACOG practice bulletin. Diagnosis and management of preeclampsia and eclampsia. Number 33, January 2002. Obstet. Gynecol. 2002, 99, 159–167. [Google Scholar]
- American College of Obstetricians and Gynecologists. ACOG Practice Bulletin No. 203: Chronic Hypertension in Pregnancy. Obstet. Gynecol. 2019, 133, e26–e50. [Google Scholar] [CrossRef]
- Vermillion, S.T.; Scardo, J.A.; Newman, R.B.; Chauhan, S.P. A randomized, double-blind trial of oral nifedipine and intravenous labetalol in hypertensive emergencies of pregnancy. Am. J. Obstet. Gynecol. 1999, 181, 858–861. [Google Scholar] [CrossRef]
- Pintye, D.; Sziva, R.E.; Mastyugin, M.; Török, M.; Jacas, S.; Lo, A.; Salahuddin, S.; Zsengellér, Z.K. Nitroxide-HMP-Protects Human Trophoblast HTR-8/SVneo Cells from H2O2-Induced Oxidative Stress by Reducing the HIF1A Signaling Pathway. Antioxidants 2023, 12, 1578. [Google Scholar] [CrossRef]
- Biwer, L.A.; Lu, Q.; Ibarrola, J.; Stepanian, A.; Man, J.J.; Carvajal, B.V.; Camarda, N.D.; Zsengeller, Z.; Skurnik, G.; Seely, E.W.; et al. Smooth Muscle Mineralocorticoid Receptor Promotes Hypertension after Preeclampsia. Circ. Res. 2023, 132, 674–689. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pintye, D.; Sziva, R.E.; Biwer, L.A.; Karreci, E.S.; Jacas, S.; Mastyugin, M.; Török, M.; Young, B.C.; Jagtap, P.; Southan, G.J.; et al. A Novel Dual-Function Nitric Oxide Donor Therapy for Preeclampsia—A Proof-of-Principle Study in a Murine Model. Antioxidants 2023, 12, 2036. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12122036
Pintye D, Sziva RE, Biwer LA, Karreci ES, Jacas S, Mastyugin M, Török M, Young BC, Jagtap P, Southan GJ, et al. A Novel Dual-Function Nitric Oxide Donor Therapy for Preeclampsia—A Proof-of-Principle Study in a Murine Model. Antioxidants. 2023; 12(12):2036. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12122036
Chicago/Turabian StylePintye, Diana, Réka E. Sziva, Lauren A. Biwer, Esilida Sula Karreci, Sonako Jacas, Maxim Mastyugin, Marianna Török, Brett C. Young, Prakash Jagtap, Garry J. Southan, and et al. 2023. "A Novel Dual-Function Nitric Oxide Donor Therapy for Preeclampsia—A Proof-of-Principle Study in a Murine Model" Antioxidants 12, no. 12: 2036. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12122036