

Dieckol, Derived from the Edible Brown Algae Ecklonia cava, Attenuates Methylglyoxal-Associated Diabetic Nephropathy by Suppressing AGE–RAGE Interaction
 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Antiglycation Property
2.3. Cell Culture
2.4. Cell Viability
2.5. Intracellular ROS Production
2.6. In Silico Molecular Docking Study
2.7. Intracellular MGO Concentration
2.8. Intracellular AGEs Accumulation
2.9. Apoptosis Analysis
2.10. Western Blot
2.11. Statistical Analysis
3. Results
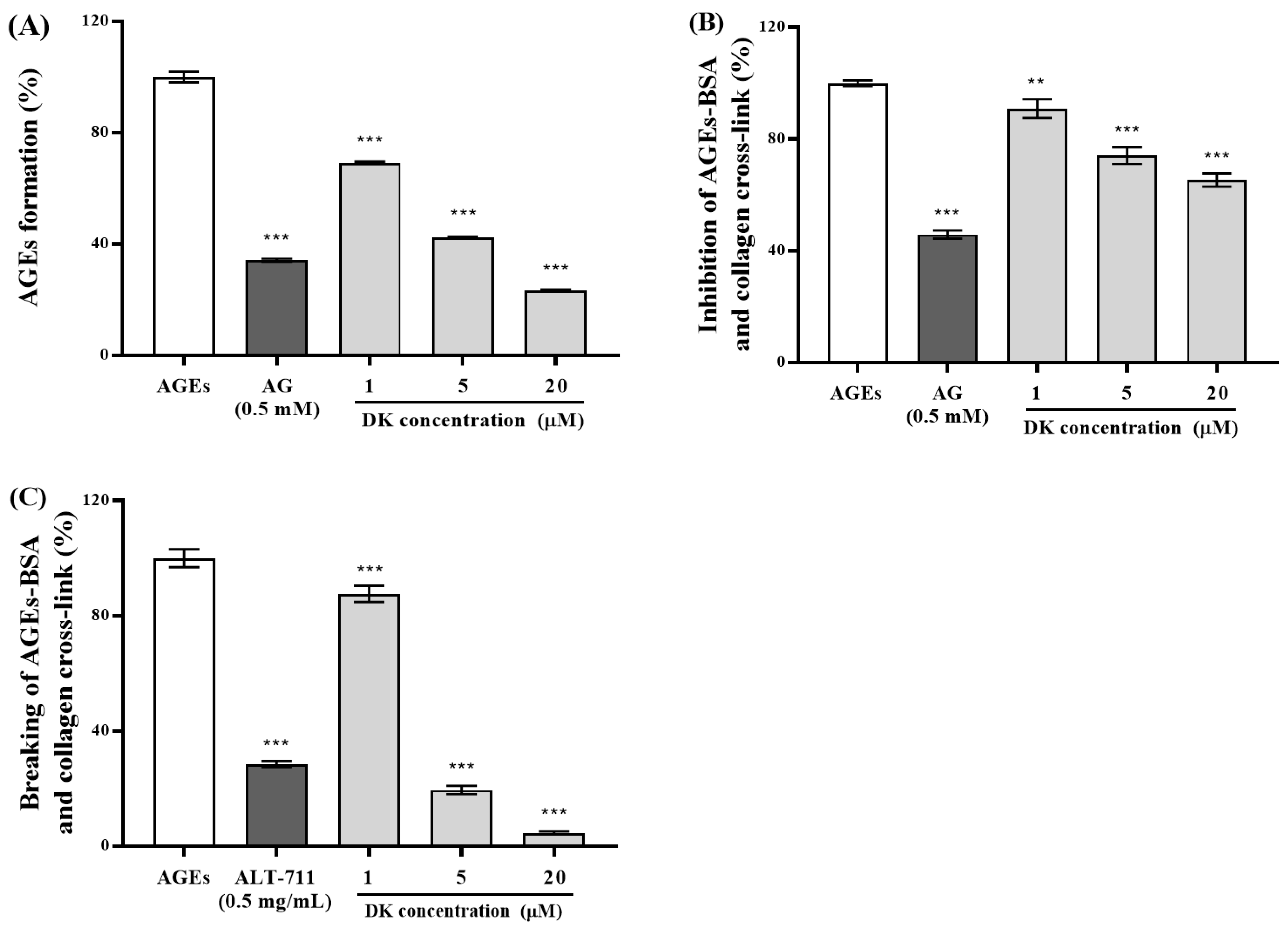
3.1. Antiglycation Property of DK
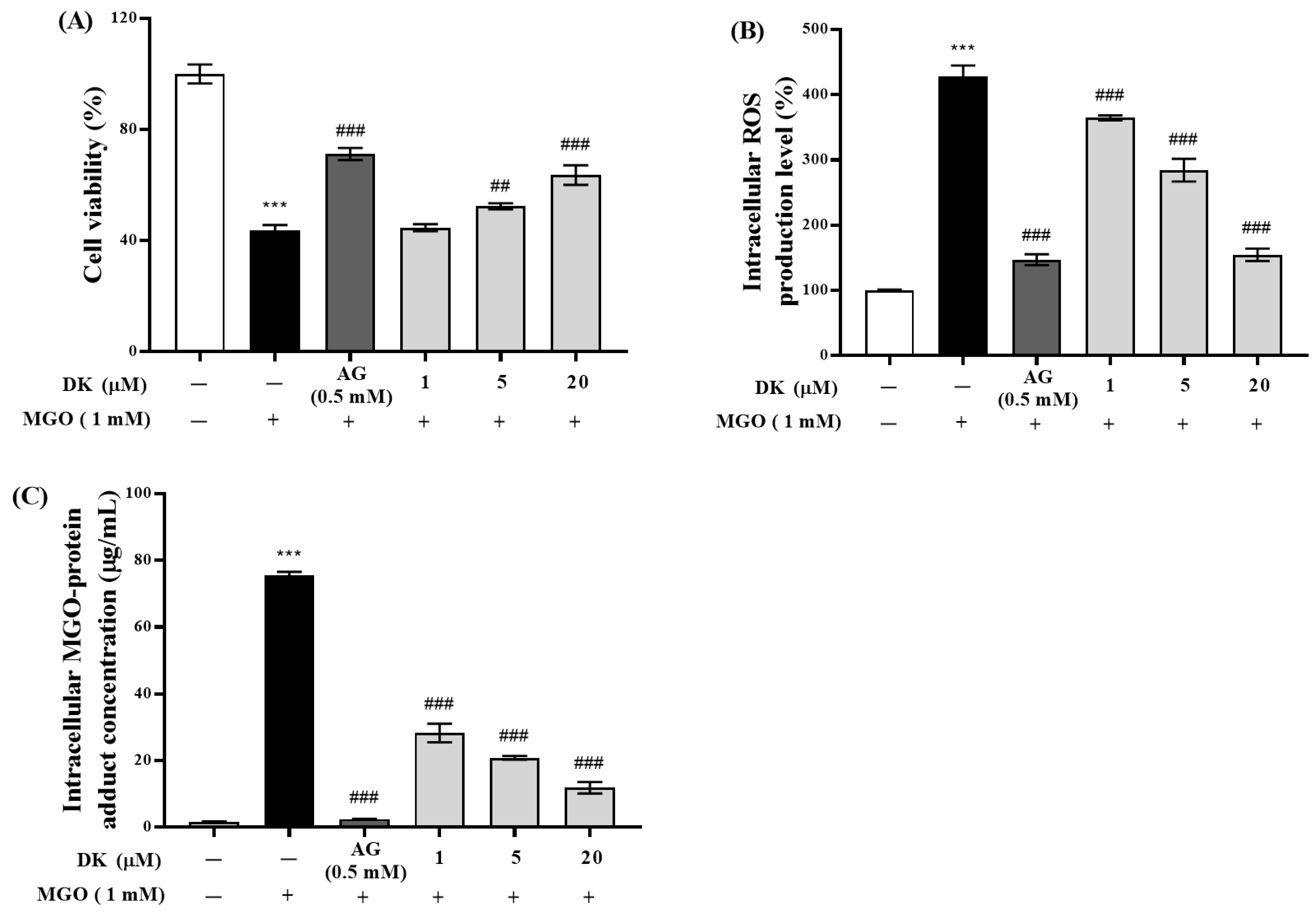
3.2. Protective Effect of DK against MGO-Induced Renal Damage
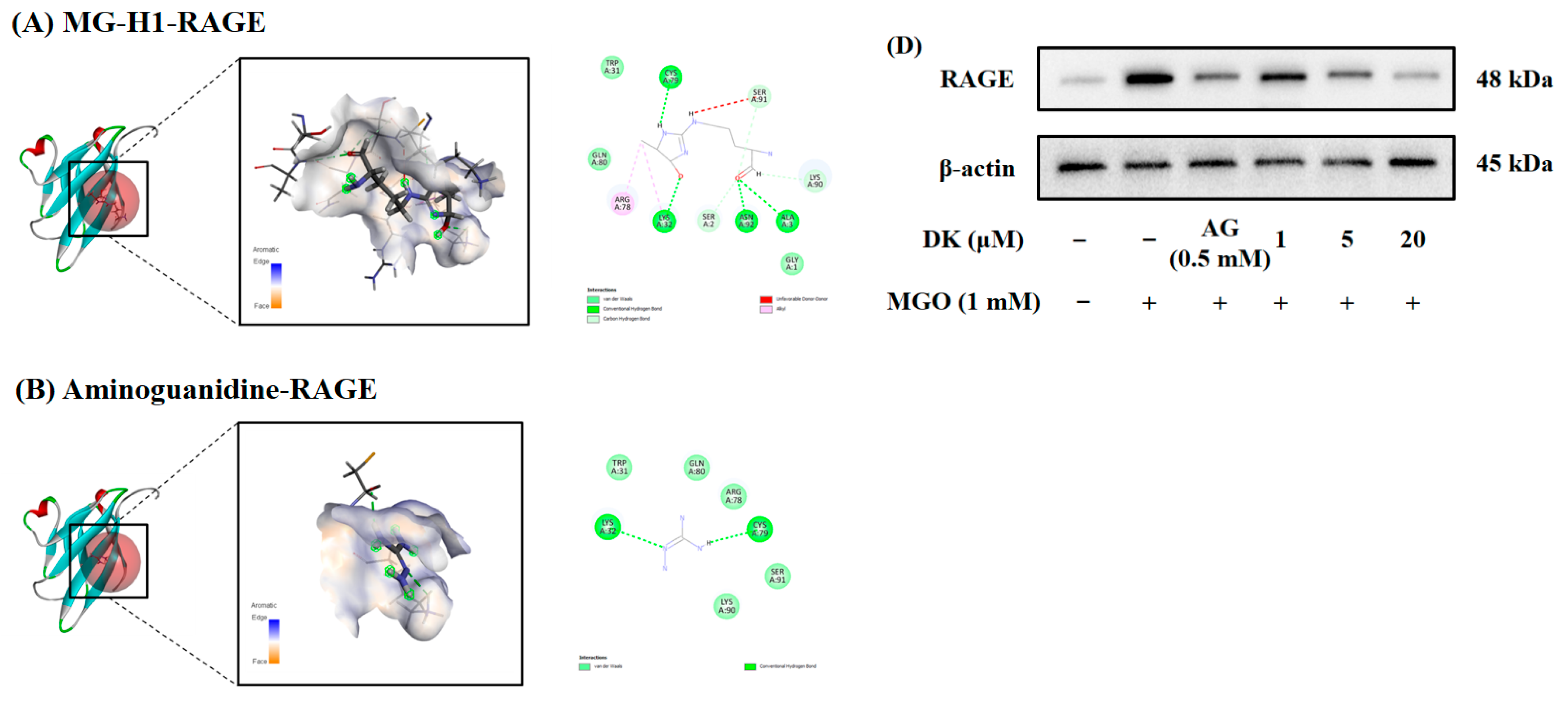
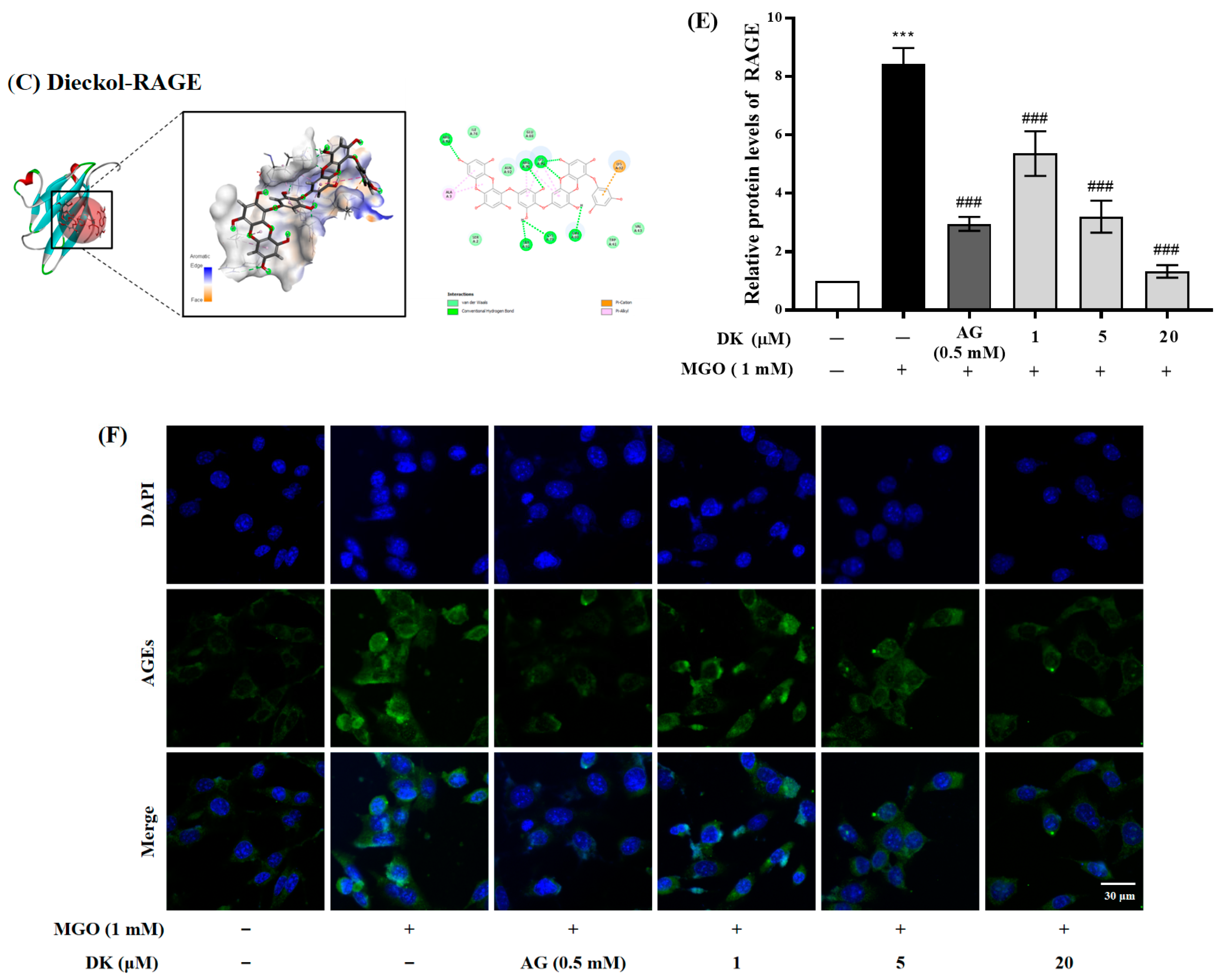
3.3. Molecular Docking Analtsis and Effect of DK on AGE/RAGE Axis in Renal Cells
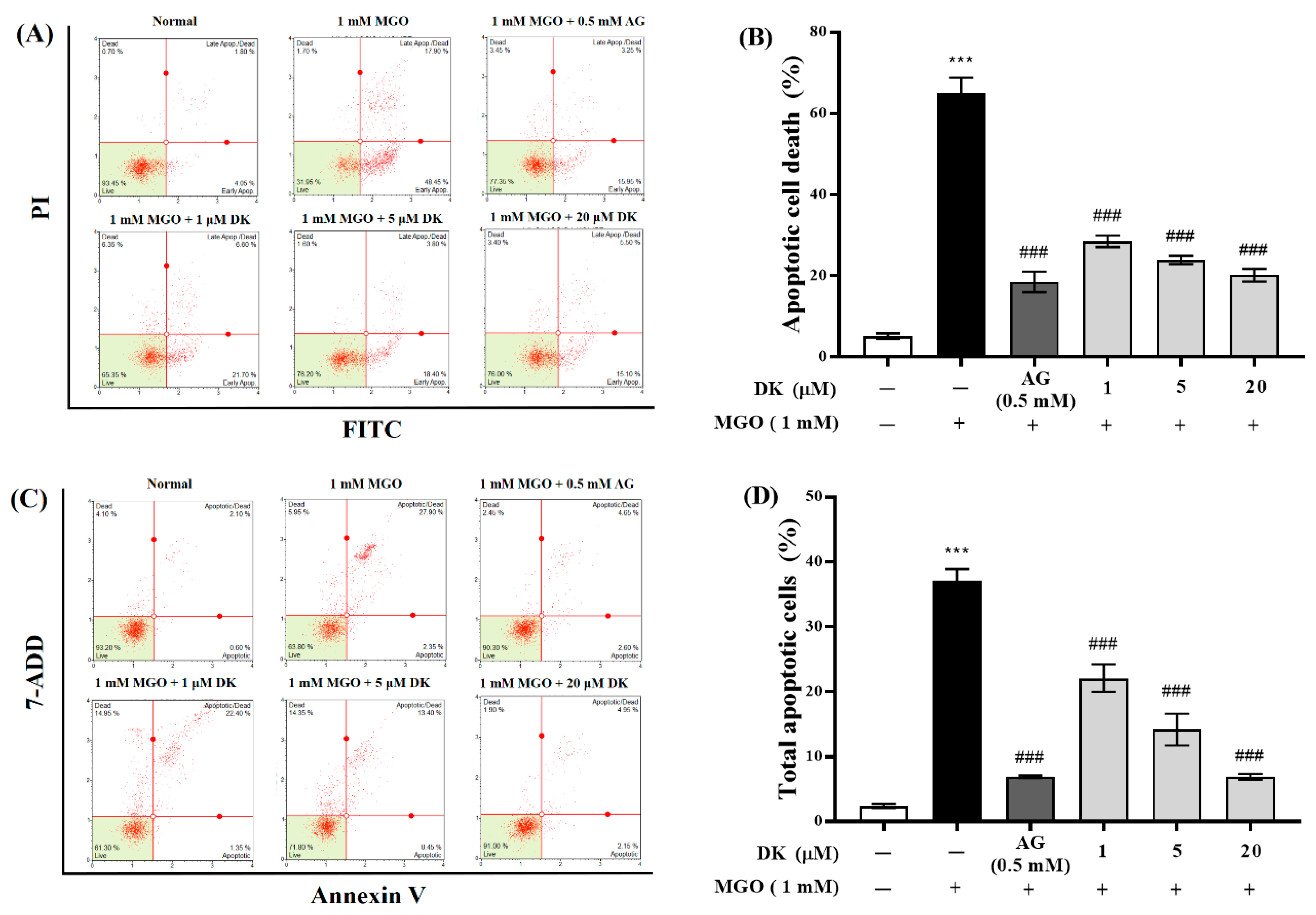
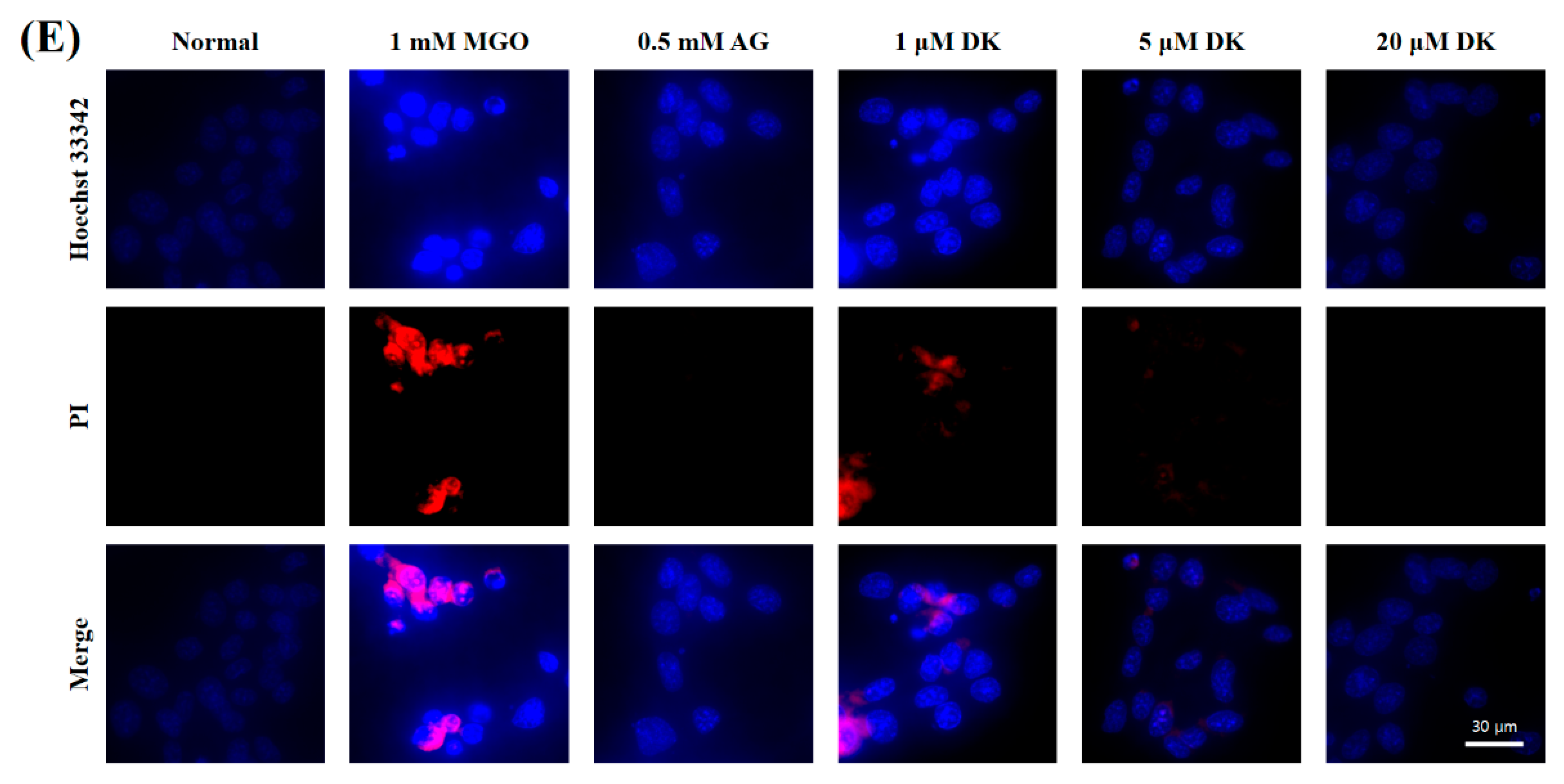
3.4. Preventive Ability of DK against MGO-Induced Apoptotic Cell Death
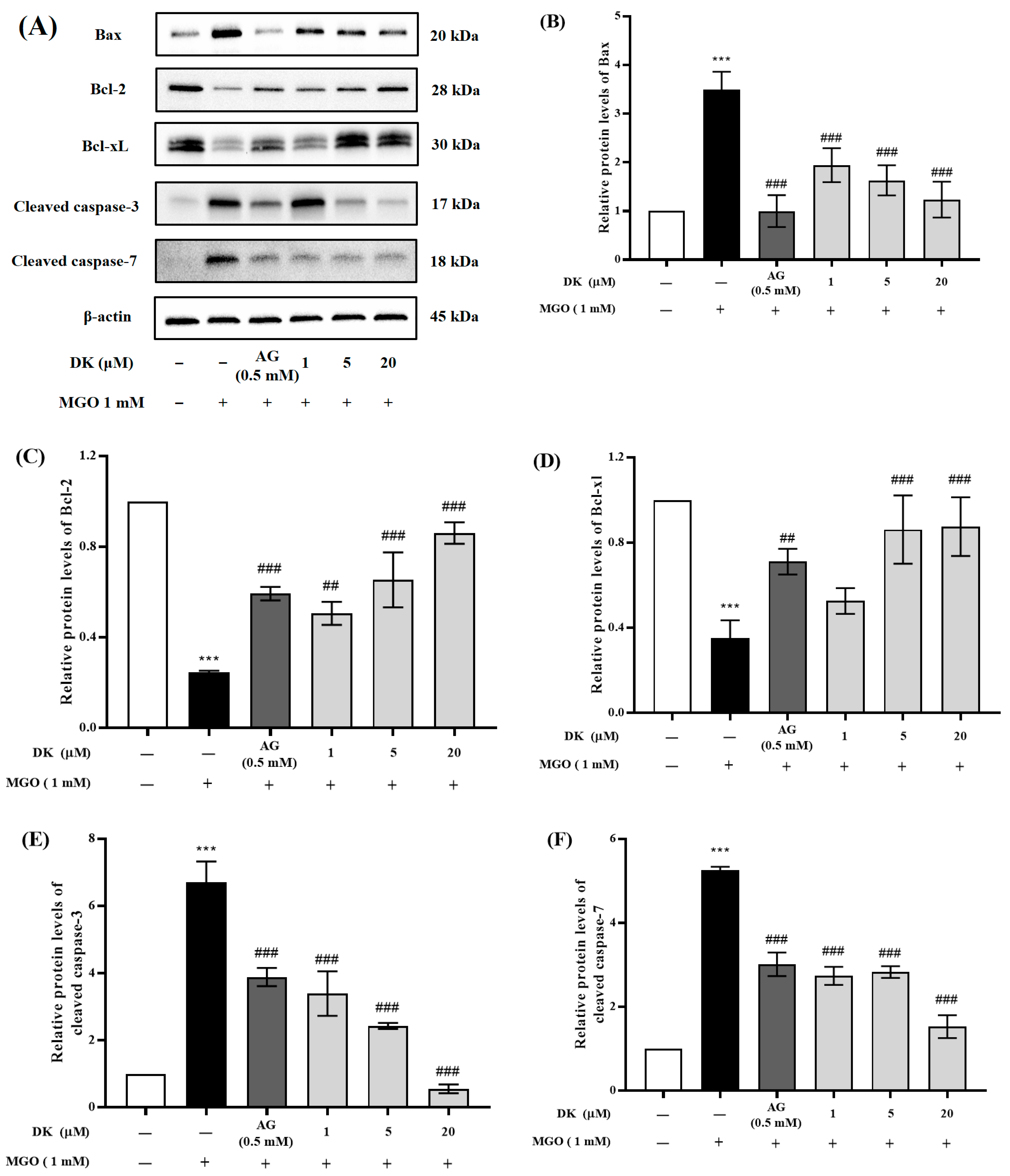
3.5. Effect of DK on the Apoptosis-Related Protein Expression
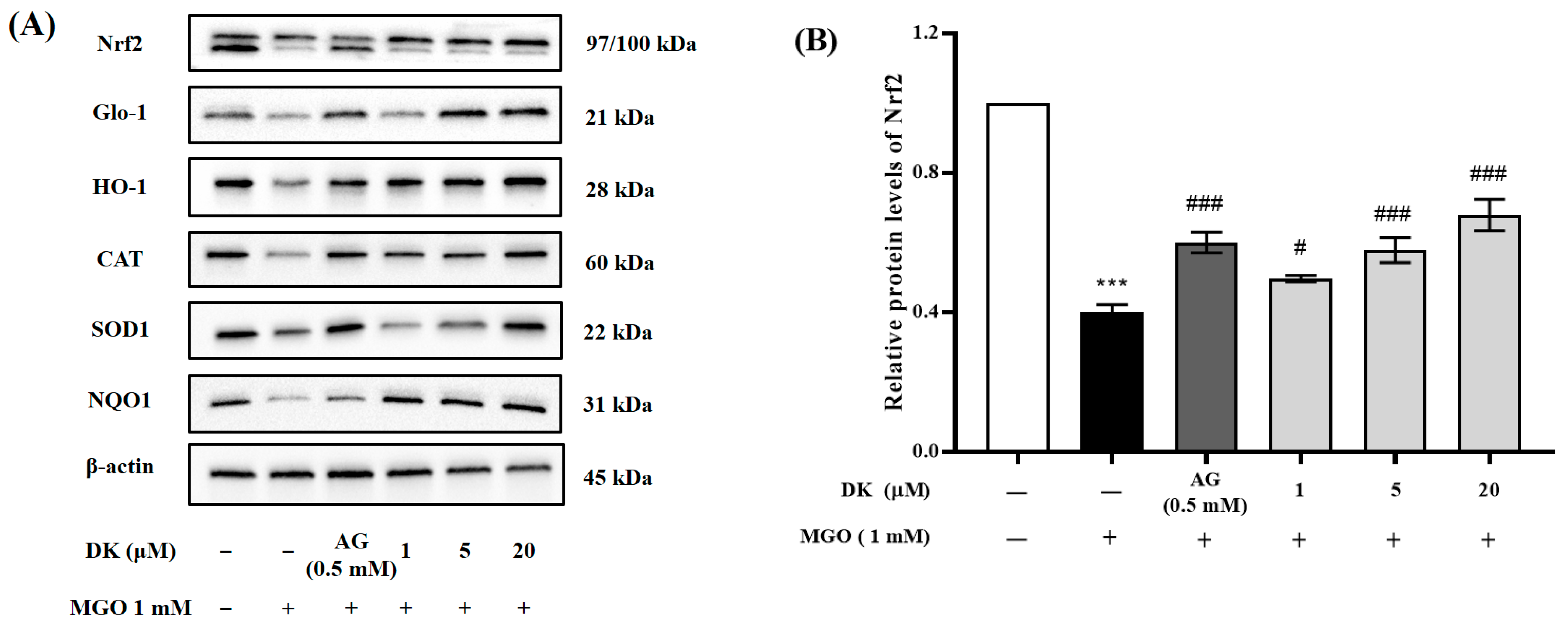
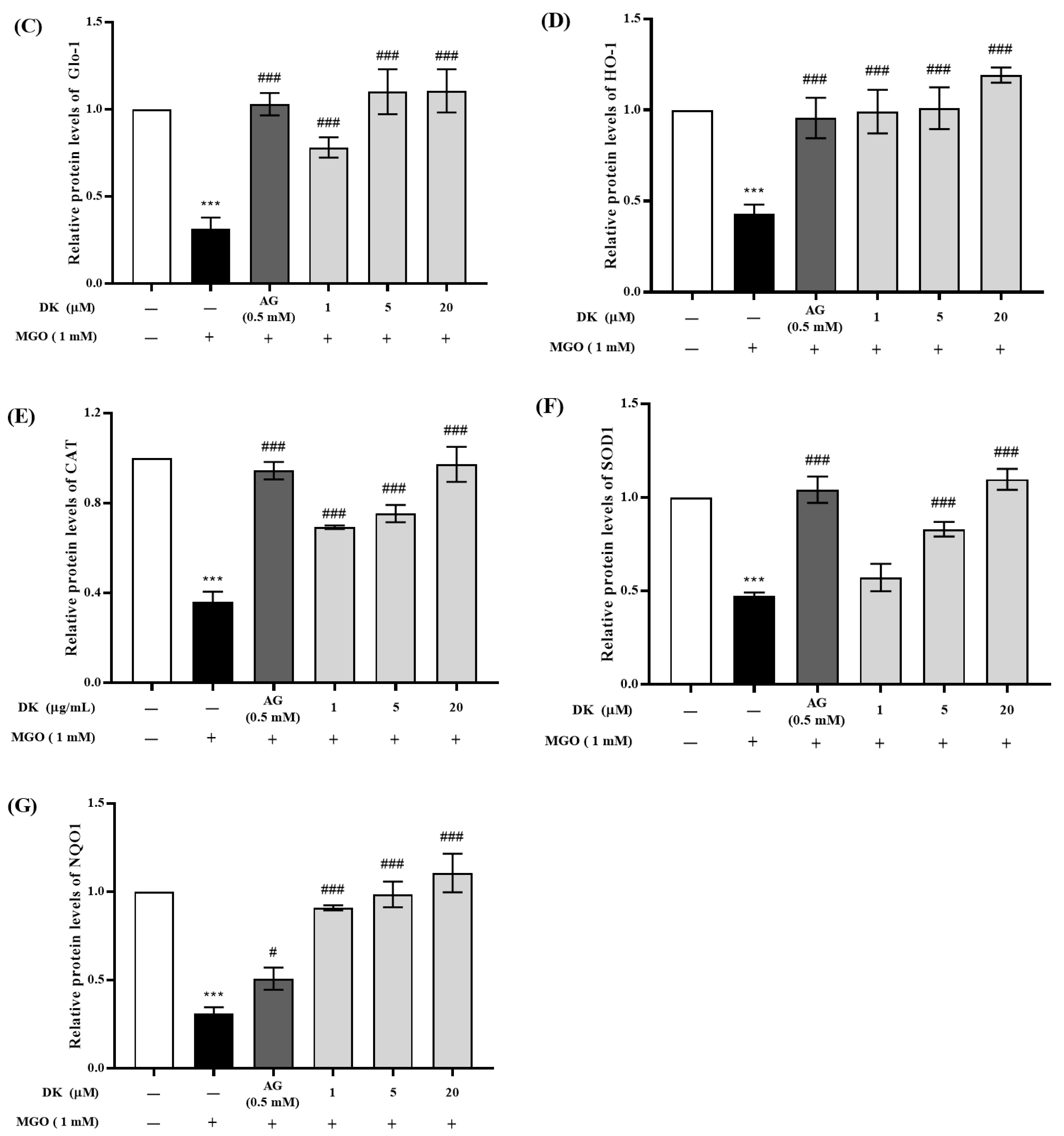
3.6. Effect of DK on Nrf2/Glo-1/ARE Signaling Pathway
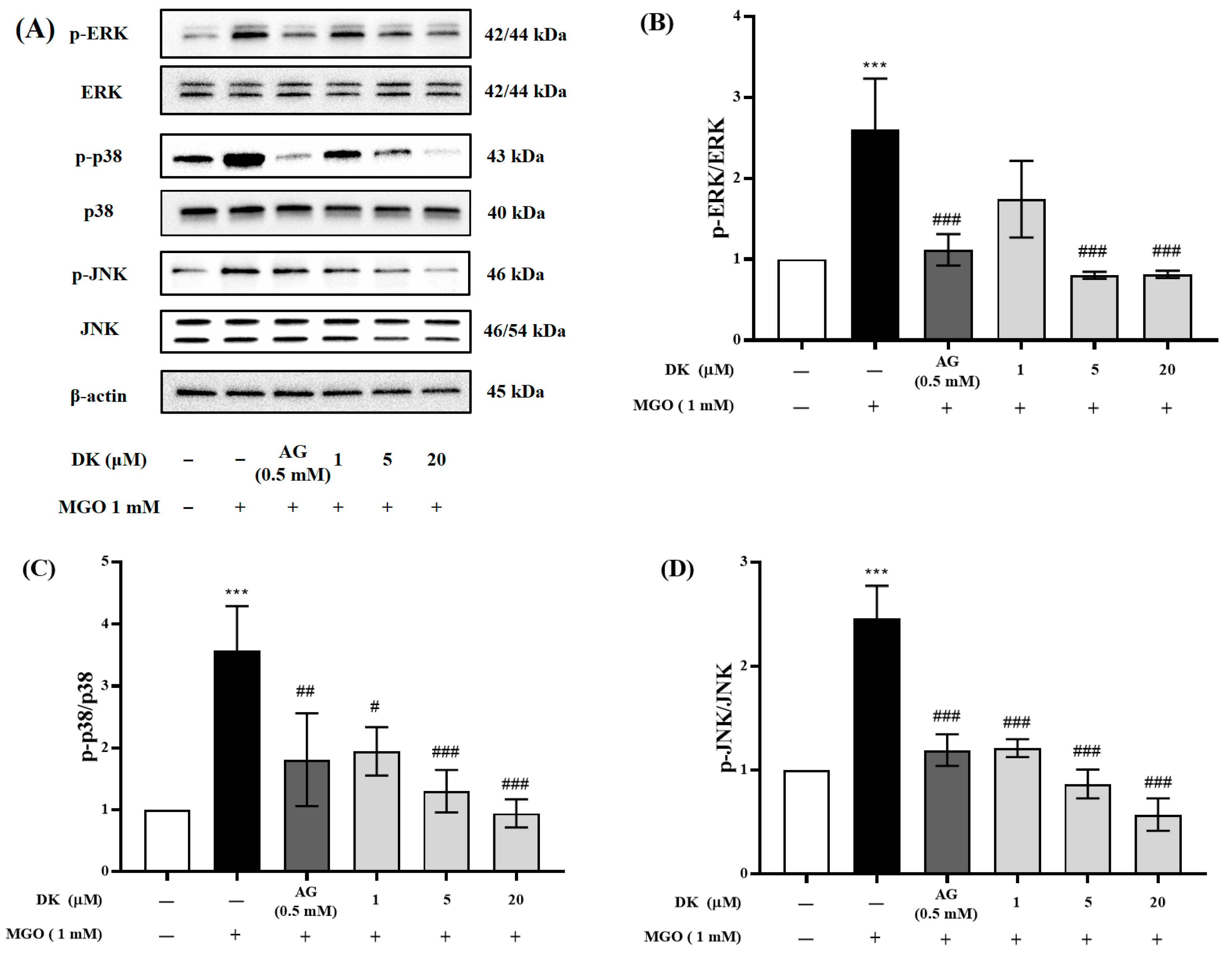
3.7. Effect of DK on MAPKs’ Phosphorylation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rabbani, N.; Thornalley, P.J. Advanced glycation end products in the pathogenesis of chronic kidney disease. Kidney Int. 2018, 93, 803–813. [Google Scholar] [CrossRef] [Green Version]
- Golej, J.; Hoeger, H.; Radner, W.; Unfried, G.; Lubec, G. Oral administration of methylglyoxal leads to kidney collagen accumulation in the mouse. Life Sci. 1998, 63, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N. Advanced glycation endproducts-role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 2005, 67, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Rowan, S.; Bejarano, E.; Taylor, A. Mechanistic targeting of adcanced glycation end-products in age-related disease. Biochim. Biophys. Acta. Mol.- Mol. Basis Dis. 2018, 1864, 3631–3643. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Galiniak, S.; Bartosz, G. Kinetics of glycoxidation of bovine serum albumin by methylglyoxal and glyoxal and its prevention by various compounds. Molecules 2014, 19, 4880–4896. [Google Scholar] [CrossRef] [Green Version]
- Brings, S.; Fleming, T.; Freichel, M.; Muckenthaler, M.U.; Herzig, S.; Nawroth, P.P. Dicarbonyls and advanced glycation end-products in the development of diabetic complications and targets for intervention. Int. J. Mole. Sci. 2017, 18, 984. [Google Scholar] [CrossRef] [Green Version]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a highly reactive dicarbonyl compound, in diabetes, its vascular complications, and other age-related disease. Physiol. Rev. 2020, 100, 407–461. [Google Scholar] [CrossRef]
- Rodrigues, L.; Matafome, P.; Crisóstomo, J.; Santos-Silva, D.; Sena, C.; Pereira, P.; Seiça, R. Advanced glycation end products and diabetic nephropathy: A comparative study using diabetic and normal rats with methylglyoxal-induced glycation. J. Physiol. Biochem. 2013, 70, 173–184. [Google Scholar] [CrossRef]
- Wijesinghe, W.A.J.P.; Jeon, Y.-J. Exploiting biological activities of brown seaweed Ecklonia cava for potential industrial applications: A review. Int. J. Food Sci. Nutr. 2012, 63, 225–235. [Google Scholar] [CrossRef]
- Wijesekara, I.; Yoon, N.Y.; Kim, S.-K. Phlorotannins from Ecklonia cava (Phaeophyceae): Biological activities and potential health benefits. BioFactors 2010, 36, 408–414. [Google Scholar] [CrossRef]
- Lee, S.-H.; Han, J.-S.; Heo, S.-J.; Hwang, J.-Y.; Jeon, Y.-J. Protective effects of dieckol isolated from Ecklonia cava against high glucose-induced oxidative stress in human umbilical vein endothelial cells. Toxicol. Vitr. 2010, 24, 375–381. [Google Scholar] [CrossRef]
- Kiho, T.; Kato, M.; Usui, S.; Hirano, K. Effect of buformin and metformin on formation of advanced glycation end products by methylglyoxal. Clin. Chim. Acta 2005, 358, 139–145. [Google Scholar] [CrossRef]
- Do, M.H.; Choi, J.; Kim, Y.; Park, H.-Y.; Park, Y.; Ha, S.K.; Hur, J. Schizonepeta tenuifolia reduces methylglyoxal-induced cytotoxicity and oxidative stress in mesangial cells. J. Funct. Foods 2019, 62, 103531. [Google Scholar] [CrossRef]
- Cho, C.H.; Jung, Y.S.; Kim, J.M.; Nam, T.G.; Lee, S.-H.; Cho, H.S.; Song, M.C.; Heo, H.J.; Kim, D.-O. Neuroprotective effects of Actinidia eriantha cv. Bidan kiwifruit on amyloid beta-induced neuronal damages in PC-12 cells and ICR mice. J. Funct. Foods 2021, 79, 104398. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Santos, R.N.D.; Oliva, G.; Andricopulo, A.D. Molecular docking and structure-based drug design strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef] [Green Version]
- Twarda-Clapa, A.; Olczak, A.; Bialkowska, A.M.; Koziolkiewicz, M. Advanced glycation end-products (AGEs): Formation, chemistry, classification, receptors, and disease related to AGEs. Cells 2022, 11, 1312. [Google Scholar] [CrossRef]
- Sharma, C.; Kaur, A.; Thind, S.S.; Singh, B.; Raina, S. Advanced glycation end-products (AGEs): An emerging concern for processed food industries. J. Food Sci. Technol. 2015, 52, 7561–7576. [Google Scholar] [CrossRef] [Green Version]
- Vlassara, H.; Palace, M.R. Diabetes and advanced glycation endproducts. J. Intern. Med. 2022, 251, 87–101. [Google Scholar] [CrossRef]
- Tan, A.L.Y.; Forbes, J.M.; Cooper, M.E. AGE, RAGE, and ROS in diabetic nephropathy. Semin. Nephrol. 2007, 27, 130–143. [Google Scholar] [CrossRef]
- Sanajou, D.; Haghjo, A.G.; Argani, H.; Aslani, S. AGE-RAGE acis blockade in diabetic nephropathy: Current status and future directions. Eur. J. Pharmacol. 2018, 833, 158–164. [Google Scholar] [CrossRef]
- Yoon, S.-R.; Shim, S.-M. Inhibitory effect of polyphenols in Houttuynia cordata on advanced glycation end-products (AGEs) by trapping methylglyoxal. LWT-Food Sci. Technol. 2015, 61, 158–163. [Google Scholar] [CrossRef]
- Lo, C.-Y.; Hsiao, W.-T.; Chen, X.-Y. Efficiency of trapping methylglyoxal by phenols and phenolic acids. J. Food Sci. 2011, 76, H90–H96. [Google Scholar] [CrossRef]
- Zhu, D.; Wang, L.; Zhou, Q.; Yan, S.; Li, Z.; Sheng, J.; Zhang, W. (+)-Catechin ameliorates diabetic nephropathy by trapping methylglyoxal in type 2 diabetic mice. Mol. Nutr. Food Res. 2014, 58, 2249–2260. [Google Scholar] [CrossRef]
- Lv, L.; Shao, X.; Chen, H.; Ho, C.-T.; Sang, S. Genistein inhibits advanced glycation end product formation by trapping methylglyoxal. Chem. Res. Toxicol. 2011, 24, 579–586. [Google Scholar] [CrossRef]
- Bednarska, K.; Kuś, P.; Fecka, I. Investigation of the phytochemical composition, antioxidant activity, and methylglyoxal trapping effect of Galega officinalis L. Herb in vitro. Molecules 2020, 25, 5810. [Google Scholar] [CrossRef]
- Cho, C.H.; Youm, G.H.; Kim, M.; Kim, S.; Song, E.-J.; Nam, Y.-D.; Lee, S.-H. Evaluation of the relationship between bioactive components in seaweeds and advanced glycation end-products inhibitory activities using principal component analysis. Plant Food Hum. Nutr. 2021, 76, 326–333. [Google Scholar] [CrossRef]
- Cho, C.H.; Lee, C.-J.; Kim, M.-G.; Ryu, B.; Je, J.-G.; Kim, Y.; Lee, S.-H. Therapeutic potential of phlorotannin-rich Ecklonia cava extract on methylglyoxa-induced diabetic nephropathy in in vitro model. Mar. Drugs 2022, 20, 355. [Google Scholar] [CrossRef]
- Rabbani, N.; Thornalley, P.J. Methylglyoxal, glyoxalase 1 and the dicarbonyl proteome. Amino Acids 2012, 42, 1133–1142. [Google Scholar] [CrossRef]
- Berner, A.K.; Brouwers, O.; Pringle, R.; Klaassen, I.; Colhoun, L.; McVicar, C.; Brockbant, S.; Curry, J.W.; Miyata, T.; Brownlee, M.; et al. Protection against methylglyoxal-derived AGEs by regulation of glyoxalase 1 prevent retinal neuroglial and vasodegenerative pathology. Diabetologia 2012, 55, 845–854. [Google Scholar] [CrossRef]
- He, Y.; Yang, Z.; Pi, J.; Cai, T.; Xia, Y.; Cao, X.; Liu, J. EGCG attenuates the neurotoxicity of methylglyoxal via regulating MAPK and the downstream signaling pathways and inhibiting advanced glycation end products formation. Food Chem. 2022, 384, 132358. [Google Scholar] [CrossRef]
- Nishimoto, S.; Koike, S.; Inoue, N.; Suzuki, T.; Ogasawara, Y. Activation of Nrf2 attenuates carbonyl stress induced by methylglyoxal in human neuroblastoma cells: Increase in GSH levels is a critical event for the detoxification mechanism. Biochem. Biophys. Res. Commun. 2017, 483, 874–879. [Google Scholar] [CrossRef]
- Adelusi, T.I.; Du, L.; Hao, M.; Zhou, X.; Xuan, Q.; Apu, C.; Sun, Y.; Lu, Q.; Yin, X. Keap1/Nrf2/ARE signaling unfolds therapeutic targets for redox imbalanced-mediated disease and diabetic nephropathy. Biomed. Pharmacother. 2020, 123, 109732. [Google Scholar] [CrossRef]
- Kellow, N.J.; Savige, G.S. Dietary advanced glycation end-products restriction for the attenuation of insulin resistance, oxidative stress and endothelial dysfunction: A systematic review. Eur. J. Clin. Nutr. 2013, 67, 239–248. [Google Scholar] [CrossRef] [Green Version]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, C.-H.; Yoo, G.; Kim, M.; Kurniawati, U.D.; Choi, I.-W.; Lee, S.-H. Dieckol, Derived from the Edible Brown Algae Ecklonia cava, Attenuates Methylglyoxal-Associated Diabetic Nephropathy by Suppressing AGE–RAGE Interaction. Antioxidants 2023, 12, 593. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12030593
Cho C-H, Yoo G, Kim M, Kurniawati UD, Choi I-W, Lee S-H. Dieckol, Derived from the Edible Brown Algae Ecklonia cava, Attenuates Methylglyoxal-Associated Diabetic Nephropathy by Suppressing AGE–RAGE Interaction. Antioxidants. 2023; 12(3):593. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12030593
Chicago/Turabian StyleCho, Chi-Heung, Guijae Yoo, Mingyeong Kim, Ulfah Dwi Kurniawati, In-Wook Choi, and Sang-Hoon Lee. 2023. "Dieckol, Derived from the Edible Brown Algae Ecklonia cava, Attenuates Methylglyoxal-Associated Diabetic Nephropathy by Suppressing AGE–RAGE Interaction" Antioxidants 12, no. 3: 593. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12030593