
Phytochemical Characterization, Antioxidant, and Anti-Proliferative Activities of Wild and Cultivated Nigella damascena Species Collected in Sicily (Italy)
 , , , , , , , ,
, , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
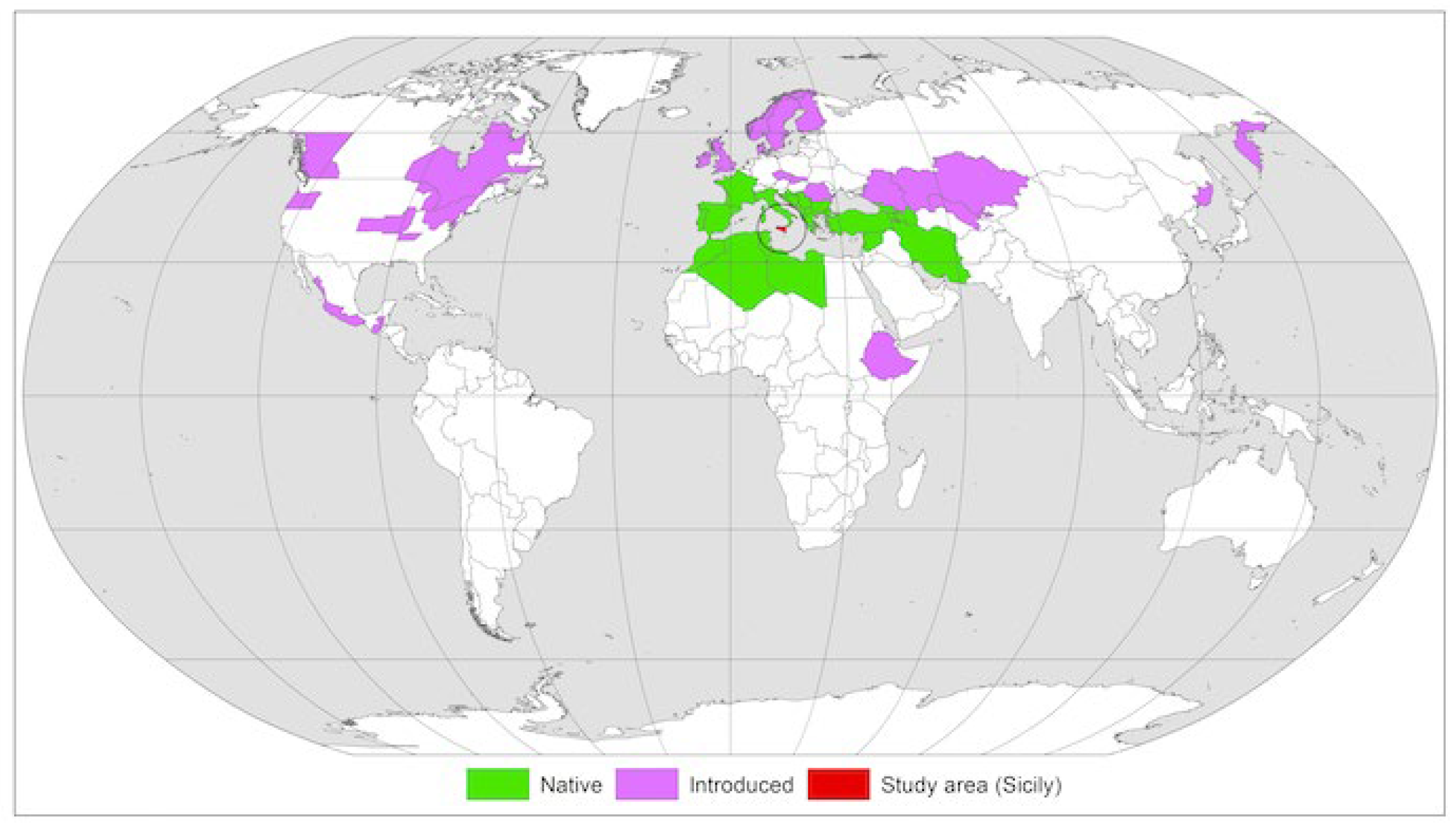
2.1. Plant Materials
2.2. Extraction Procedure
2.3. Metabolite Profiling
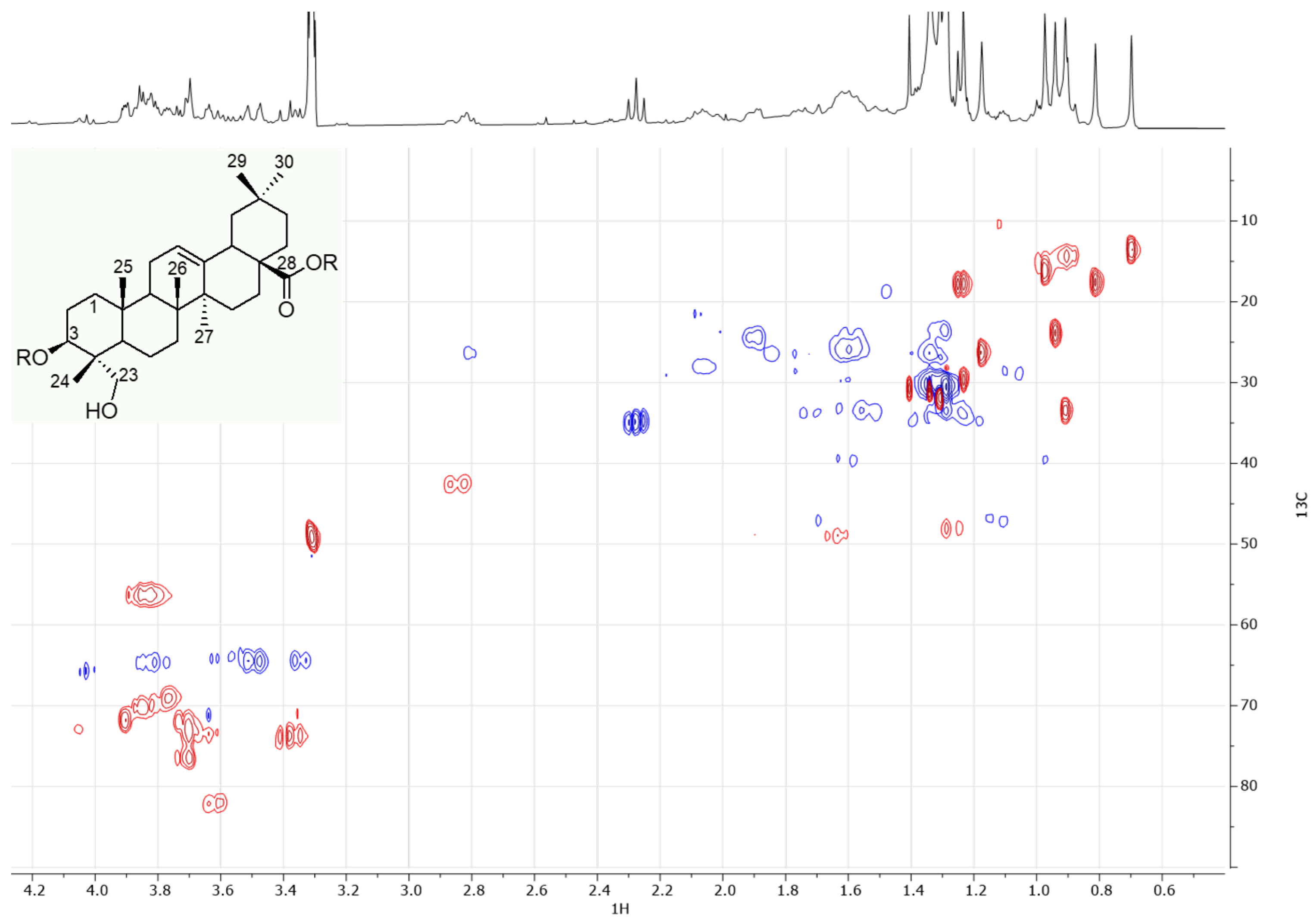
2.3.1. NMR Experiments
2.3.2. UHPLC-ESI-QqTOF HR MS/MS Experiments
2.4. Cell Lines and Culture Conditions
2.4.1. MTT Assay
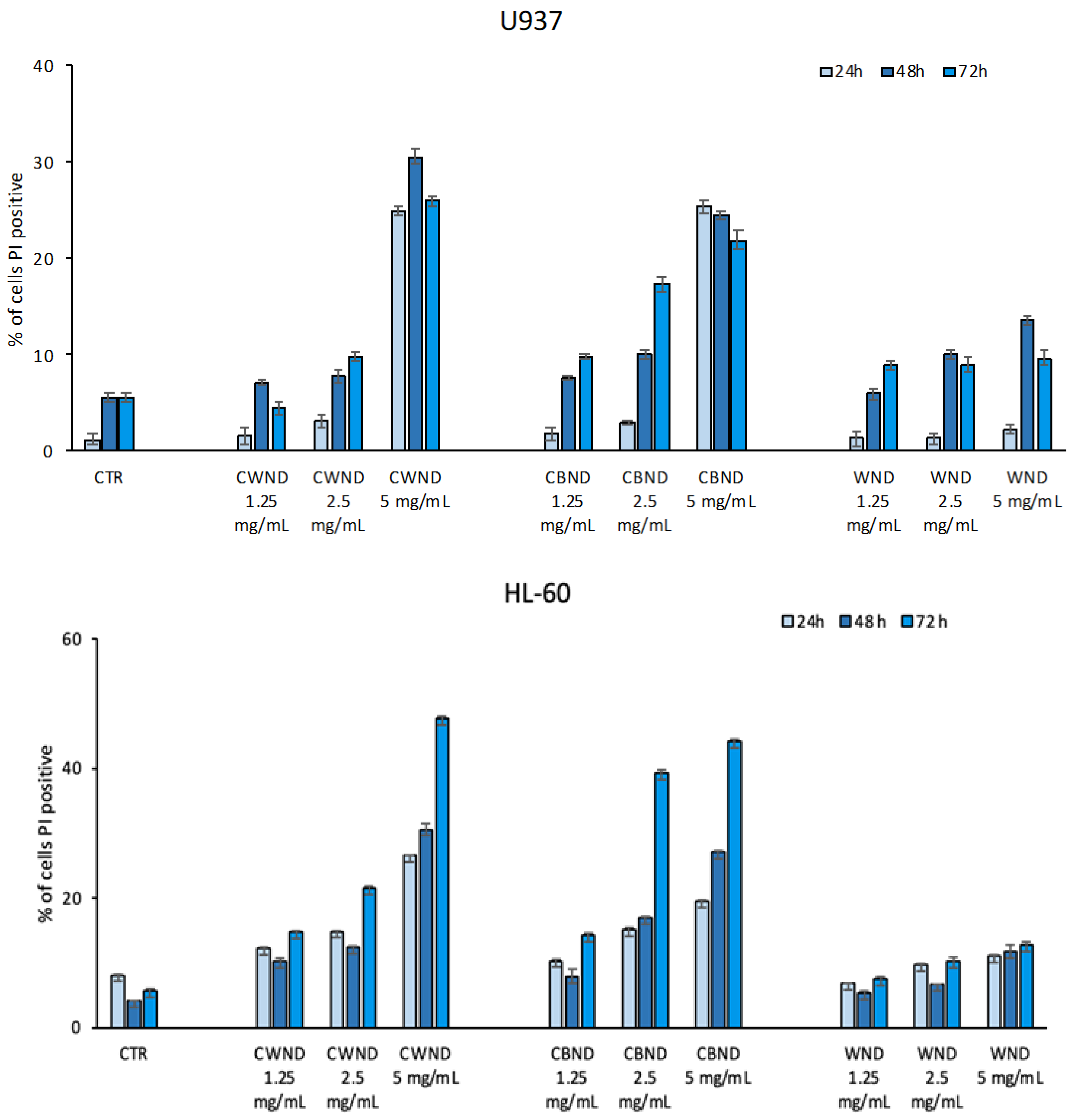
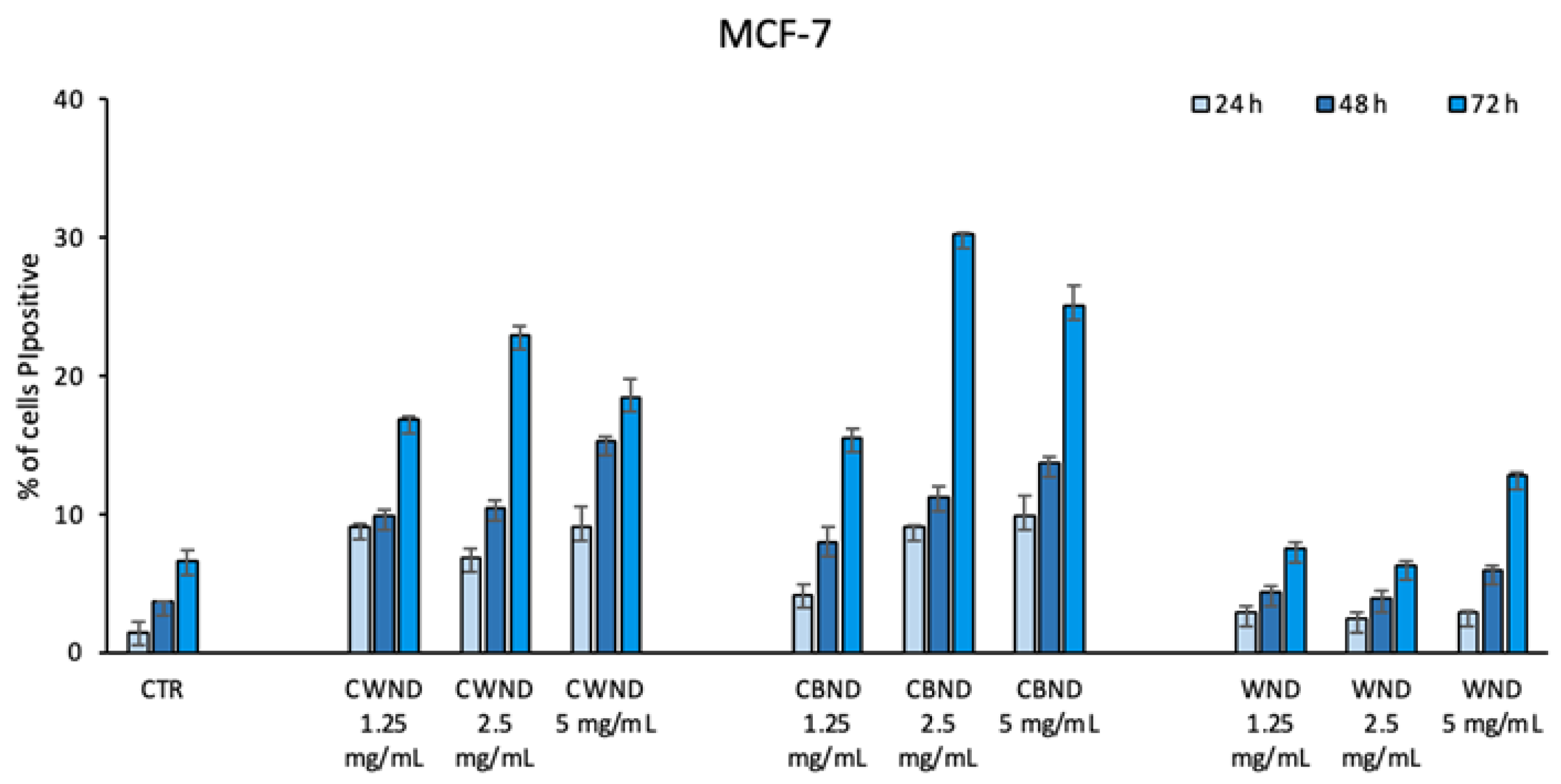
2.4.2. FACS Analysis
2.4.3. Polymorphonuclear Leukocytes (PMN) Isolation
2.5. Reactive Oxygen Species (ROS) Generation and Antioxidant Enzymes Measured in PMN Cells
2.6. Statistics
3. Results
3.1. Chemical Characterization of the Extracts
3.2. Biological Properties
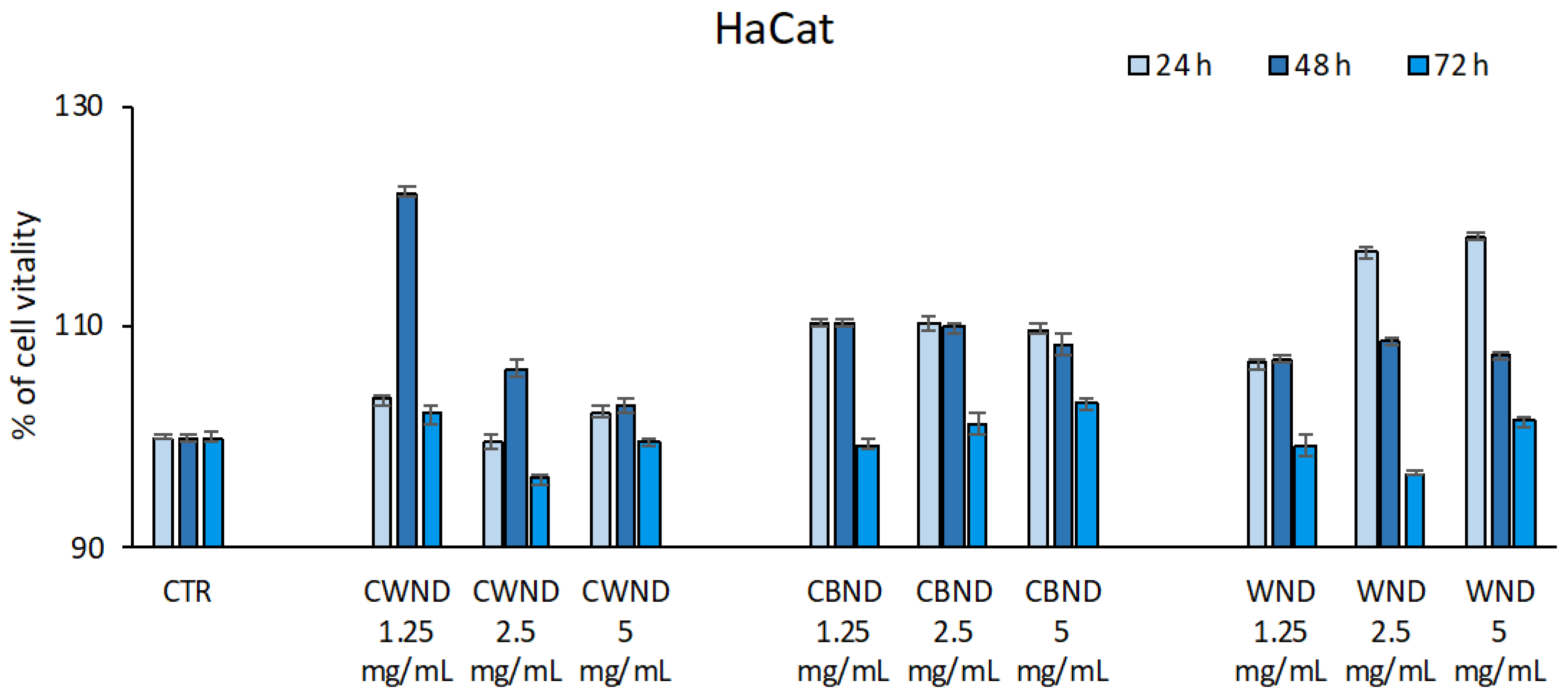
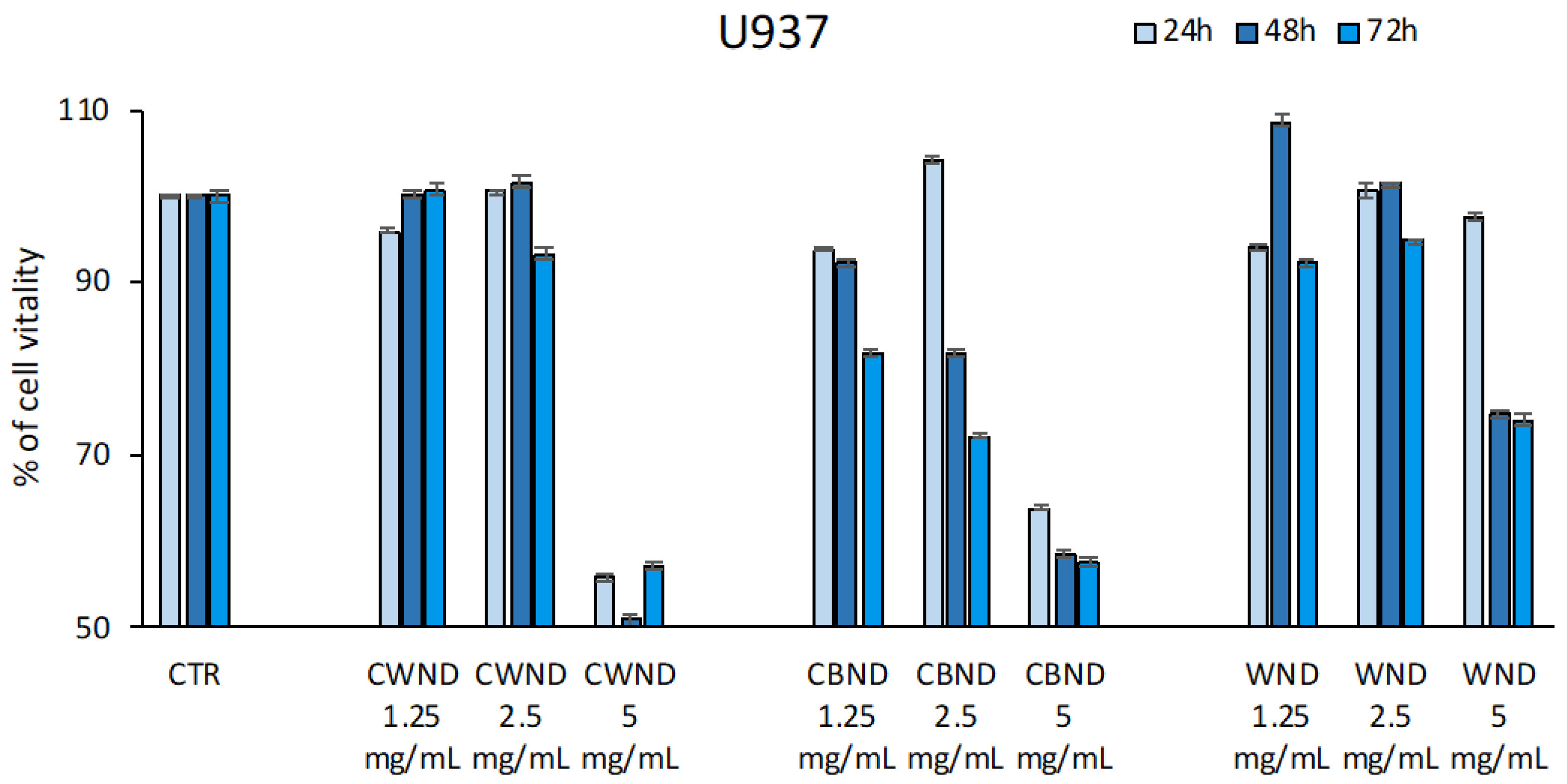
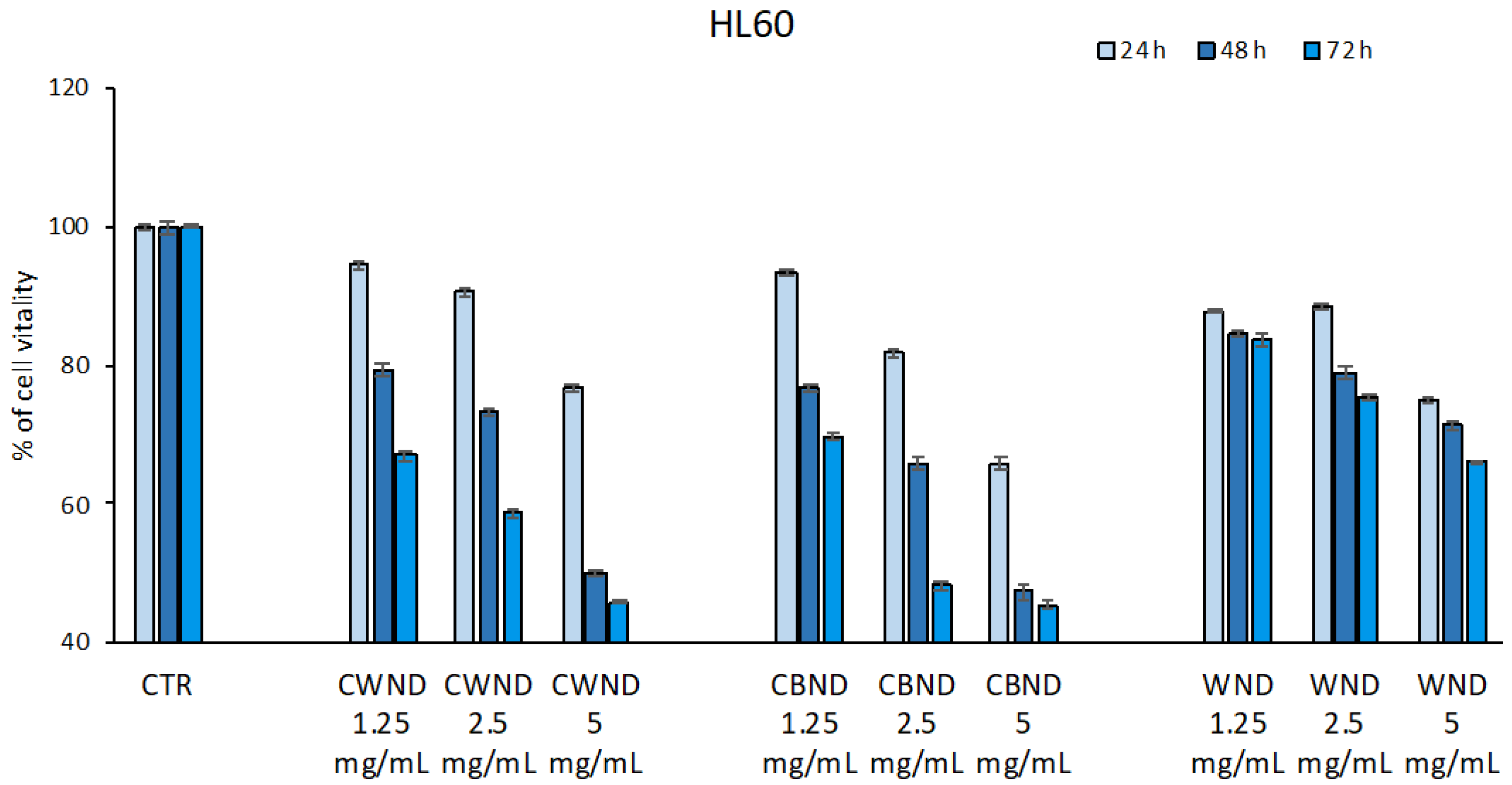
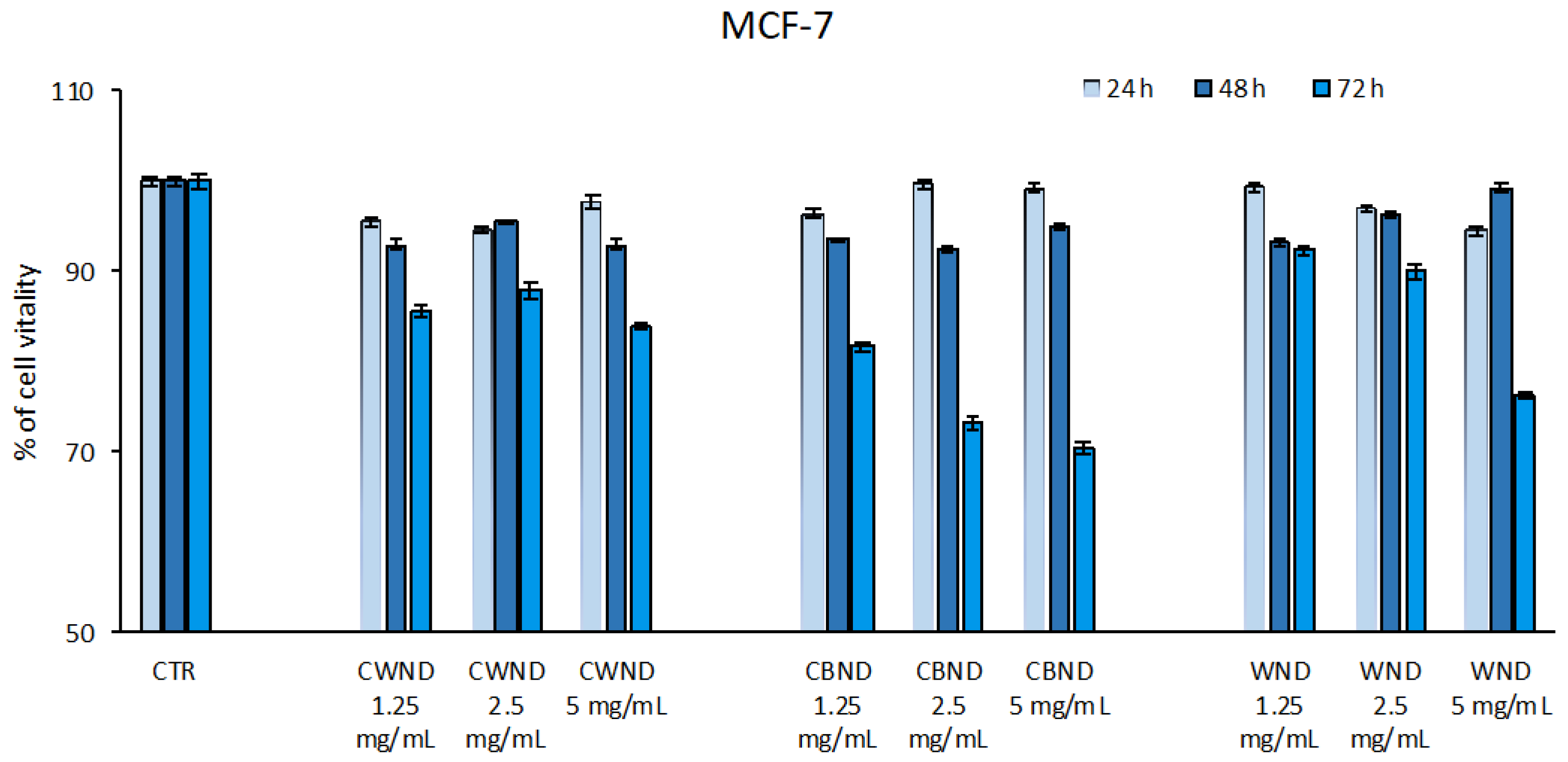
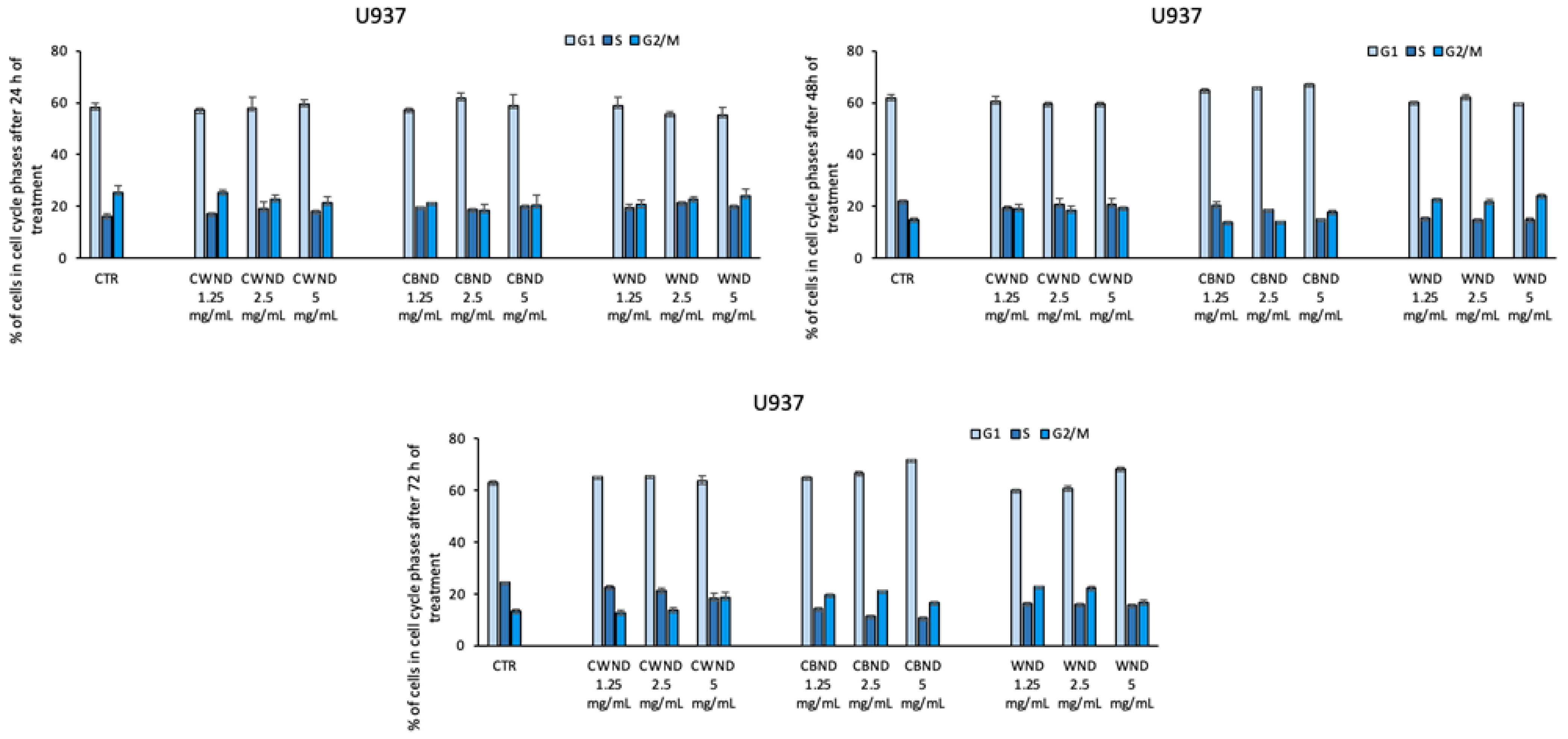
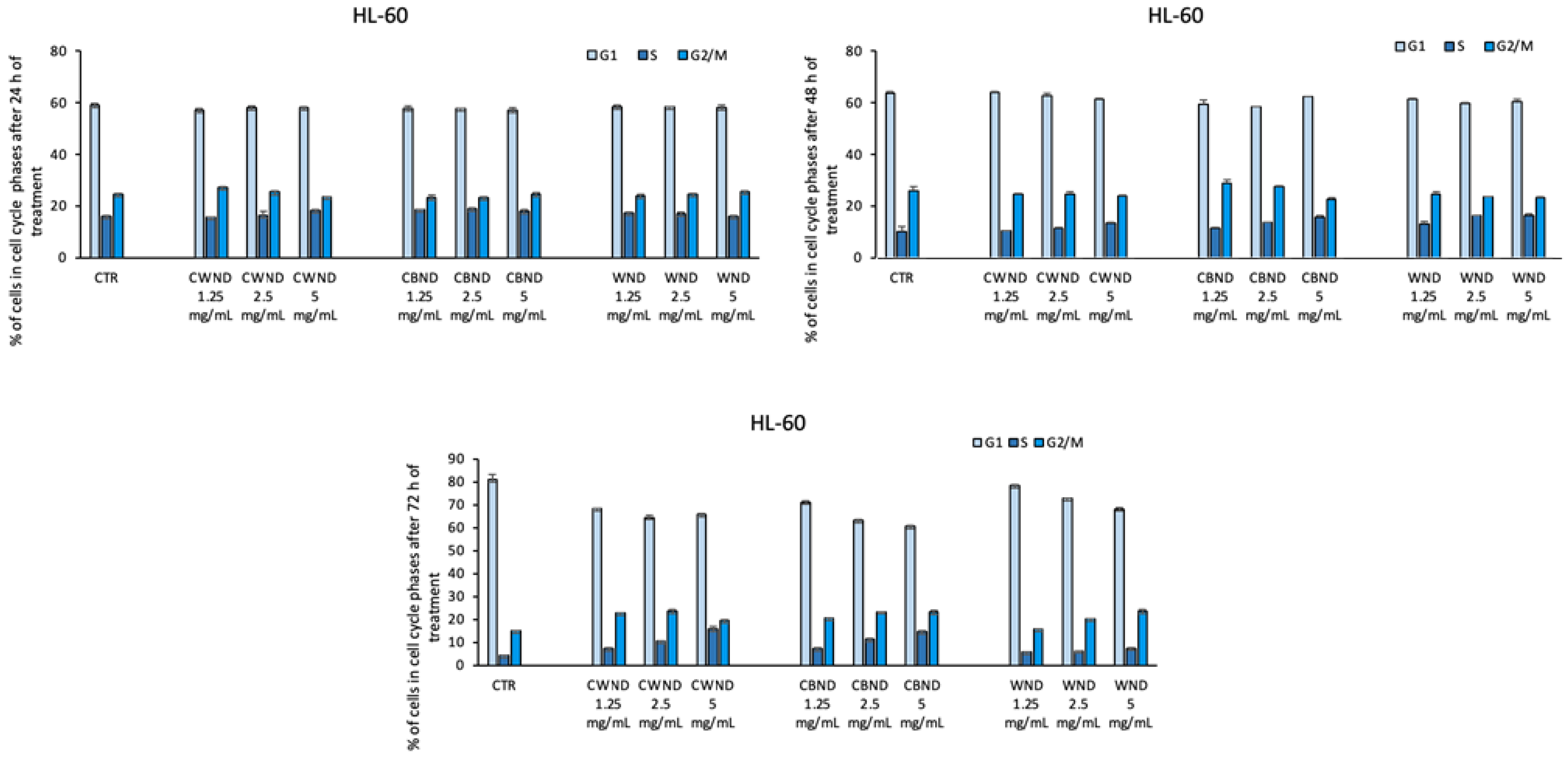
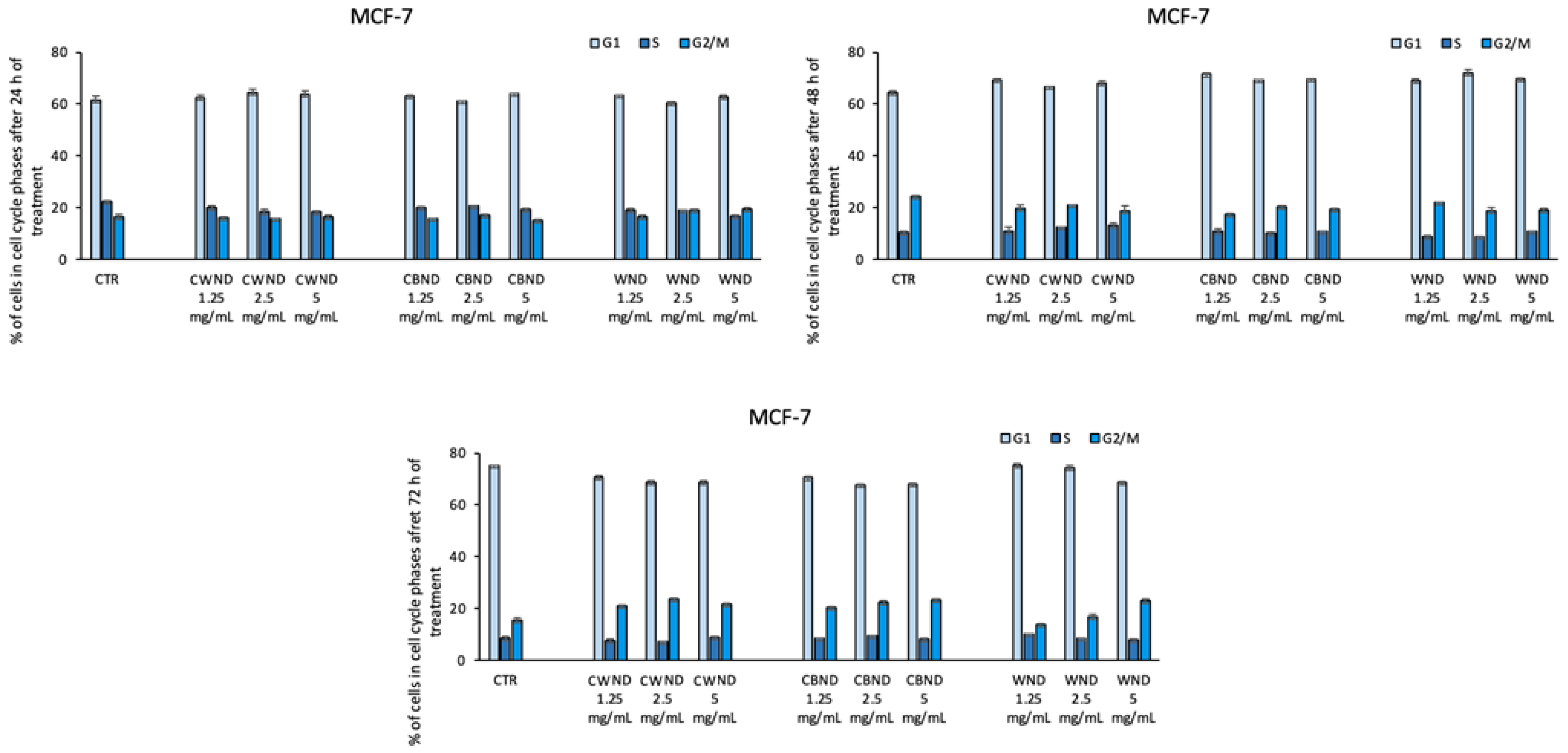
3.2.1. Biological Effect of CWND, CBND, and WND in Hematological and Solid Cancer Cells
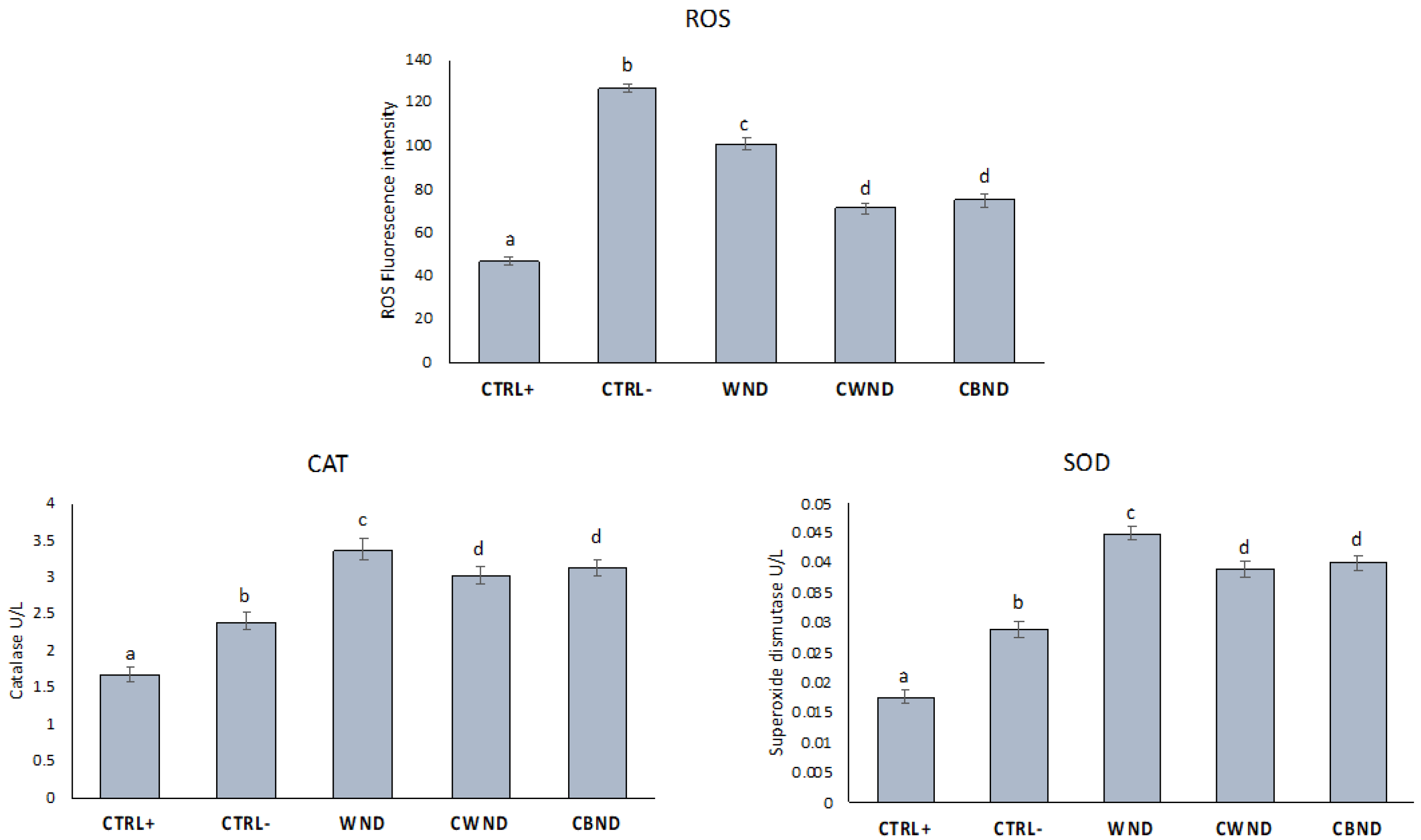
3.2.2. ROS Generation and Antioxidant Enzymes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet [WWW Document]. 2023. Available online: http://www.plantsoftheworldonline.org/ (accessed on 22 November 2023).
- ESRI. ESRI Data & Maps Environmental Systems Research Institute, Redlands (CA) USA. 2002. Available online: https://www.esri.com/en-us/home (accessed on 22 November 2023).
- Badalamenti, N.; Modica, A.; Bazan, G.; Marino, P.; Bruno, M. The ethnobotany, phytochemistry, and biological properties of Nigella damascena—A review. Phytochemistry 2022, 198, 113165. [Google Scholar] [CrossRef]
- Heiss, A.G.; Stika, H.P.; De Zorzi, N.; Jursa, M. Nigella in the mirror of time a brief attempt to draw a genus’ ethnohistorical portrait. In Festschrift für Helmut Johannes Kroll Zum 65; Geburtstag; Wachholtz Verlag: Hamburg, Germany, 2012; pp. 147–169. [Google Scholar]
- Jabbour, F.; Pasquier, P.E.D.; Chazalviel, L.; Guilloux, M.L.; Conde E Silva, N.; Deveaux, Y.; Manicacci, D.; Galipot, P.; Heiss, A.G.; Damerval, C. Evolution of the distribution area of the Mediterranean Nigella damascena and a likely multiple molecular origin of its perianth dimorphism. Flora 2021, 274, 151735. [Google Scholar] [CrossRef]
- Huxley, A.; Griffiths, M. The New Royal Horticultural Society Dictionary of Gardening. In Dictionary of Gardening; MacMillan, P., Ed.; MacMillan Press: London, UK, 1992; Volume 4, pp. 154–168. [Google Scholar]
- Pignatti, S. Flora D’italia, 2nd ed.; Editor Edagricole: Milan, Italy, 2017; Volume 1. [Google Scholar]
- Linné, C.V. Species Plantarum, Exhibentes Plantas Rite Cognitas, ad Genera Relatas, Cum Differentiis Specificis, Nominibus Trivialibus, Synonymis Selectis, Locis Natalibus, Secundum Systema Sexuale Digestas; Impensis Laurentii Salvii, Holmiae: Stockholm, Sweden, 1753. [Google Scholar] [CrossRef]
- Fiori, A. Nuova Flora Analitica D’italia; Tipografia di M. Ricci: Firenze, Italy, 1923. [Google Scholar] [CrossRef]
- Toma, C.C.; Olah, N.K.; Vlase, L.; Mogoșan, C.; Mocan, A. Comparative studies on polyphenolic composition, antioxidant and diuretic effects of Nigella sativa L. (Black Cumin) and Nigella damascena L. (lady-in-a-Mist) seeds. Molecules 2015, 20, 9560–9574. [Google Scholar] [CrossRef]
- Ballero, M.; Fresu, I. Application of medicinal plants in Seui’s Barbagia (Central Sardinia, Italy). Fitoterapia 1993, 64, 141–150. [Google Scholar]
- Agradi, E.; Fico, G.; Cillo, F.; Francisci, C.; Tomè, F. Estrogenic activity of phenolic compounds from Nigella damascena evaluated using a recombinant yeast screen. Planta Med. 2001, 67, 553–555. [Google Scholar] [CrossRef] [PubMed]
- Fico, G.; Braca, A.; Tomè, F.; Morelli, I. Phenolic derivatives from Nigella damascena seeds. Pharm. Biol. 2000, 38, 371–373. [Google Scholar] [CrossRef]
- Heiss, A.G.; Oeggl, K. The oldest evidence of Nigella damascena L. (Ranunculaceae) and its possible introduction to central Europe. Veget. Hist. Archaeobot. 2005, 14, 562–570. [Google Scholar] [CrossRef]
- Geraci, A.; Polizzano, V.; Schicchi, R. Ethnobotanical uses of wild taxa as galactagogues in Sicily (Italy). Acta Soc. Bot. Pol. 2018, 87, 1–18. [Google Scholar] [CrossRef]
- Aleo, N.; Amato, F.; Aleo, M. Le piante tossiche della flora trapanese (Sicilia). Quad. Bot. Amb. Appl. 2011, 11, 31–49. [Google Scholar]
- Rodrìguez, A.D.; González, E.; Ramìrez, C. The structural chemistry, reactivity, and total synthesis of dolabellane diterpenes. Tetrahedron 1998, 54, 11683–11729. [Google Scholar] [CrossRef]
- Mohamed, K.M.; Ohtani, K.; Kasai, R.; Yamadi, K. Dolabellane diterpene glucosides from Chrozophora obliqua. Phytochemistry 1994, 37, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, K.M.; Ohtani, K.; Kasai, R.; Yamasaki, K. 3-Hydroxy-3-methylglutaryl dolabellane diterpenes from Chrozophora obliqua. Phytochemistry 1995, 39, 151–161. [Google Scholar] [CrossRef]
- Cao, X.W.; Wang, F.J.; Liew, O.W.; Zhao, J. Analysis of triterpenoid saponins reveals insights into structural features associated with potent protein drug enhancement effects. Mol. Pharm. 2020, 17, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Nakamura, S.; Asada, Y.; Yamashita, M.; Matsuda, H. Oxazonigelladine and dolabellane-type diterpene constituents from Nigella damascena seeds. Tetrahedron 2017, 73, 7054–7060. [Google Scholar] [CrossRef]
- Ogawa, K.; Nakamura, S.; Hosokawa, K.; Ishimaru, H.; Saito, N.; Ryu, K.; Fujimuro, M.; Nakashima, S.; Matsuda, H. New diterpenes from Nigella damascena seeds and their antiviral activities against herpes simplex virus type-1. J. Nat. Med. 2018, 72, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Sieniawska, E.; Michel, P.; Mroczek, T.; Granica, S.; Skalicka-Wožniak, K. Nigella damascena L. essential oil and its main constituents, damascenine and β-elemene modulate inflammatory response of human neutrophils ex vivo. Food Chem. Toxicol. 2019, 125, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Edris, A.E. Anti-cancer properties of Nigella spp. essential oils and their major constituents, thymoquinone and β-elemene. Curr. Clin. Pharmacol. 2009, 4, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Sarno, F.; Pepe, G.; Termolino, P.; Carafa, V.; Massaro, C.; Merciai, F.; Altucci, L. Trifolium repens blocks proliferation in chronic myelogenous leukemia via the BCR-ABL/STAT5 pathway. Cells 2020, 9, 379. [Google Scholar] [CrossRef] [PubMed]
- Badalamenti, N.; Russi, S.; Bruno, M.; Maresca, V.; Vaglica, A.; Ilardi, V.; Zanfardino, A.; Di Napoli, M.; Varcamonti, M.; Cianciullo, P.; et al. Dihydrophenanthrenes from a Sicilian accession of Himantoglossum robertianum (Loisel.) P. delforge showed antioxidant, antimicrobial, and antiproliferative activities. Plants 2021, 10, 2776. [Google Scholar] [CrossRef] [PubMed]
- Manna, A.; Saha, P.; Sarkar, A.; Mukhopadhyay, D.; Bauri, A.K.; Kumar, D.; Das, P.; Chattopadhyay, S.; Chatterjee, M. Malabaricone-A induces a redox imbalance that mediates apoptosis in U937 Cell Line. PLoS ONE 2012, 7, e36938. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, P.O.; Pala, D.; Silva, C.T.; de Souza, M.O.; do Amaral, J.F.; Vieira, R.A.L.; de Freitas Folly, G.A.; Volp, A.C.P.; de Freitas, R.N. Açai (Euterpe oleracea Mart.) pulp dietary intake improves cellular antioxidantenzymes and biomarkers of serum in healthy women. Nutrition 2016, 32, 674–680. [Google Scholar] [CrossRef] [PubMed]
- Scognamiglio, M.; Fiumano, V.; D’Abrosca, B.; Esposito, A.; Choi, Y.H.; Verpoorte, R.; Fiorentino, A. Chemical interactions between plants in Mediterranean vegetation: The influence of selected plant extracts on Aegilops geniculata metabolome. Phytochemistry 2014, 106, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Scognamiglio, M.; Schneider, B. Identification of potential allelochemicals from donor plants and their synergistic effects on the metabolome of Aegilops geniculata. Front. Plant Sci. 2020, 11, 1046. [Google Scholar] [CrossRef] [PubMed]
- Sayaka, M.; Kazuyoshi, T.; Yukie, O.; Seiji, O.; Tunehiro, M.; Hajime, M. Functional and structural characterization of a flavonoid glucoside 1,6-glucosyltransferase from Catharanthus roseus. Plant Cell Physiol. 2009, 50, 1401–1415. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Aziz, S.M.S.A.; Aboshady, A.I.; Ibrahim, M.A.R.; Ibrahim, M.F.M.; Alenezi, M.A.; Darwish, D.B.E.; Al-Qahtani, S.M.; Al-Harbi, N.A.; Darwish, H.; et al. Isolation and identification of flavonoids from black cumin (Nigella sativa) by HPLC-MS and in silico molecular interactions of their major compounds with Fusarium oxysporum trypsin-like serine protease. Separations 2023, 10, 360. [Google Scholar] [CrossRef]
- Farag, M.A.; Haidy, A.; Gad, A.G.H.; Ludger, A.W. Metabolomics driven analysis of six Nigella species seeds via UPLC-qTOF-MS and GC–MS coupled to chemometrics. Food Chem. 2014, 151, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Yoshimitsu, H.; Nishida, M.; Okawa, M.; Nohara, T. Four new triterpene glycosides from Nigella damascena. Chem. Pharm. Bull. 2007, 55, 488–491. [Google Scholar] [CrossRef] [PubMed]
- Pendes, A.; Delaude, C. Structure elucidation of an acetylated saponin of Blighia welwitschii by NMR spectroscopy. Carbohydr. Res. 1994, 263, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Mukhamedziev, M.M.; Alimbaeva, P.K.; Gorovits, T.T.; Abubakirov, N.K. Structure of dipsacoside B—A triterpene glycoside from Dipsacus azureus. Chem. Nat. Compd. 1971, 7, 145–149. [Google Scholar] [CrossRef]
- Braca, A.; Autore, G.; De Simone, F.; Marzocco, S.; Morelli, I.; Venturella, F.; De Tommasi, N. Cytotoxic saponins from Schfflera rotundifolia. Planta Med. 2004, 70, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Woo, W.S. Triterpene glycosides from the roots of Patrinia scabiosifolia. Planta Med. 1987, 52, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.I.; Won, K.J.; Kim, D.Y.; Kim, H.B.; Li, Y.; Lee, H.M. Chemical composition of Patrinia scabiosifolia flower absolute and its migratory and proliferative activities in human keratinocytes. Chem. Biodivers. 2019, 16, e1900252. [Google Scholar] [CrossRef] [PubMed]
- Kawai, H.; Kuroyanagi, M.; Umehara, K.; Akira Ueno, A.; Satake, M. Studies on the saponins of Lonicera japonica Thunb. Chem. Pharm. Bull. 1988, 36, 4769–4775. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.C.; Zhang, Q.W.; Zhao, S.X.; Che, C.T. Four new oleanane saponins from Anemone anhuiensis. Chem. Pharm. Bull. 2001, 49, 632–634. [Google Scholar] [CrossRef] [PubMed]
- Parveen, A.; Farooq, M.A.; Kyunn, W.W. A new oleanane type saponin from the aerial parts of Nigella sativa with antioxidant and anti-diabetic potential. Molecules 2020, 25, 2171. [Google Scholar] [CrossRef] [PubMed]
- Toki, K.; Saito, N.; Nogami, A.; Tatsuzawa, F.; Shigihara, A.; Honda, T. Flavonoid glycosides isolated from the blue flowers of Nigella damascena. Heterocycles 2009, 78, 2287–2294. [Google Scholar] [CrossRef]
- Tanikawa, N.; Homma, K.; Tatsuzawa, F. Flavonoids of the rose-pink, blue, and white flowers of Nigella damascena L. (Ranunculaceae). Sci. Hortic. 2019, 257, 108609. [Google Scholar] [CrossRef]
- Kokoska, L.; Havlik, J.; Valterova, I.; Nepovim, A.; Rada, V.; Vanek, T. Chemical composition of the essential oil of Nigella orientalis L. seeds. Flavour Fragr. J. 2005, 20, 419–420. [Google Scholar] [CrossRef]
- Moretti, A.; D’Antuono, L.F.; Elementi, S. Essential oils of Nigella sativa L. and Nigella damascena L. seed. J. Essent. Oil Res. 2004, 16, 182–183. [Google Scholar] [CrossRef]
- Xu, K.; Zhan Shu, Z.; Xu, Q.M.; Liu, Y.L.; Li, X.R.; Wang, Y.L.; Yang, S.L. Cytotoxic activity of Pulsatilla chinensis saponins and their structure—Activity relationship. J. Asian Nat. Prod. Res. 2013, 15, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.B.; Zhang, X.J.; Zhang, R.H.; Zhu, L.L.; Pu, D.B.; Li, X.L.; Li, H.L.; Xu, M.; Xiao, W.L. In vitro human dihydroorotate dehydrogenase inhibitory, antiinflammatory and cytotoxic activities of alkaloids from the seeds of Nigella glandulifera. Planta Med. 2018, 84, 1013–1021. [Google Scholar] [CrossRef]
- Al-Jumaili, A.; Kumar, A.; Bazaka, K.; Jacob, M.V. Plant secondary metabolite-derived polymers: A potential approach to develop antimicrobial films. Polymers 2018, 10, 515. [Google Scholar] [CrossRef] [PubMed]
- Salmani, J.M.; Asgharpanah, J.; Zendehdel, K. Cytotoxic effects of Nigella damascena Extract on Human Breast and Colorectal Cancer Cell Lines. Asian Pac. J. Cancer Prev. 2014, 15, 3829–3833. [Google Scholar]
- Karimi, A.; Moradi, M.T.; Alidadi, S.; Hashemi, L.; Ghaffari, S.H. The effect of Nigella sativa extract on human glioblastoma multiforme cells: A laboratory study. Iran. J. Cancer Prev. 2013, 6, 218–224. [Google Scholar]
- Badalamenti, N.; Ilardi, V.; Rosselli, S.; Bruno, M. The ethnobotany, phytochemistry and biological properties of genus Ferulago—A review. J. Ethnopharmacol. 2021, 274, 114050. [Google Scholar] [CrossRef] [PubMed]
- Sytar, O. Phenolic acids in the inflorescences of different varieties of buckwheat and their antioxidant activity. J. King Saud Univ. Sci. 2015, 27, 136–142. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Alhamad, N.M.; Gammoh, S.; Ereifej, K.; Alodat, M.; Hussein, N.M.; Kubow, S.; Torley, P.J. Antioxidant and antihypertensive properties of phenolic–protein complexes in extracted protein fractions from Nigella damascena and Nigella arvensis. Food Hydrocoll. 2016, 56, 84–92. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Ramezani, M.; Salmani, G.A. Antinociceptive, anti-inflammatory and acute toxicity effects of Zataria multiflora Boiss extracts in mice and rats. J. Ethnopharmacol. 2007, 109, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Dalli, M.; Bekkouch, O.; Azizi, S.E.; Azghar, A.; Gseyra, N.; Kim, J. Nigella sativa L. Phytochemistry and pharmacological activities: A Review. Biomolecules 2021, 12, 20. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Fayyad, M.W. Recent advances on the anti-cancer properties of Nigella sativa, a widely used food additive. Integr. Med. 2016, 7, 173–180. [Google Scholar] [CrossRef]
- Miceli, M.; Bontempo, P.; Nebbioso, A.; Altucci, L. Natural compounds in epigenetics: A current view. Food Chem. Toxicol. 2014, 73, 71–83. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Rt | Tentative Assignment | Molecular Formula | [M-H]− Found | [M-2H]2− | RDB | Error (ppm) |
|---|---|---|---|---|---|---|---|
| 1 | 3.791 | Quercetin tetrahexoside | C39H50O27 | 949.2459 | 15 | −0.8 | |
| 2 | 4.010 | Quercetin dihexoside deoxyhexoside | C33H40O21 | 771.1993 | 14 | 0.5 | |
| 3 | 4.269 | Kaempferol dihexoside deoxyhexoside (1) | C33H40O20 | 755.2046 | 14 | 0.8 | |
| 4 | 4.308 | Kaempferol dihexoside | C27H30O16 | 609.1460 | 13 | −0.2 | |
| 5 | 4.315 | Myricetin dihexosylpentoside | C32H38O22 | 773.1751 | 14 | −4.0 | |
| 6 | 4.373 | Kaempferol dihexoside deoxyhexoside (2) | C33H40O20 | 755.2046 | 14 | −1.6 | |
| 7 | 4.409 | Quercetin trihexoside | C33H40O22 | 787.1940 | 14 | 0.2 | |
| 8 | 4.450 | Quercetin hexosylpentoside deoxyhexoside | C32H38O20 | 741.1884 | 14 | 0.9 | |
| 9 | 4.452 | Quercetin (hydromethylglutaryl) dihexoside deoxyhexoside | C39H48O25 | 915.2399 | 16 | −1.4 | |
| 10 | 4.562 | Quercetin dihexoside | C27H30O17 | 625.1426 | 13 | 2.5 | |
| 11 | 4.740 | Kaempferol (hydromethylglutaryl) dihexoside deoxyhexoside (1) | C39H48O24 | 899.2499 | 16 | 1.6 | |
| 12 | 4.883 | Kaempferol trihexoside | C33H40O21 | 771.1979 | 14 | −1.3 | |
| 13 | 4.885 | Quercetin (hydromethylglutaryl) hexosylpentoside deoxyhexoside | C38H46O24 | 885.2285 | 16 | −2.4 | |
| 14 | 4.889 | Kaempferol (hydromethylglutaryl) dihexoside deoxyhexoside (2) | C39H48O24 | 899.2452 | 16 | −1.2 | |
| 15 | 4.954 | Quercetin hexosylpentoside | C26H28O16 | 595.1308 | 13 | 0.6 | |
| 16 | 4.957 | Quercetin dihexosylpentoside | C32H38O21 | 757.1839 | 14 | 0.8 | |
| 17 | 5.032 | Kaempferol dihexoside | C27H30O16 | 609.1463 | 13 | 0.3 | |
| 18 | 5.113 | Quercetin (hydromethylglutaryl) trihexoside | C39H48O26 | 931.2346 | 16 | −1.4 | |
| 19 | 5.231 | Quercetin caffeoyltrihexoside (1) | C42H46O25 | 949.2246 | 474.1091 | 24 | −1.0 |
| 20 | 5.339 | Quercetin hexosyldeoxyhexoside | C27H30O16 | 609.1463 | 13 | 0.3 | |
| 21 | 5.471 | Quercetin hexoside | C21H20O12 | 463.0881 | 12 | −0.2 | |
| 22 | 5.505 | Quercetin caffeoyltrihexoside (2) | C42H46O25 | 949.2248 | 474.1094 | 24 | −0.8 |
| 23 | 5.560 | Quercetin (hydroxymethylglutaryl) hexosylpentoside | C32H36O20 | 739.1730 | 15 | −1.9 | |
| 24 | 5.572 | Quercetin (hydroxymethylglutaryl) dihexosylpentoside | C38H46O25 | 901.2248 | 16 | −0.8 | |
| 25 | 5.861 | Quercetin feruloyltrihexoside (1) | C43H48O25 | 963.2393 | 20 | −2.0 | |
| 26 | 6.065 | Quercetin coumaroyltrihexoside (1) | C42H46O24 | 933.2306 | 466.1118 | 20 | −1.9 |
| 27 | 6.058 | Quercetin coumaroyltrihexoside (2) | C42H46O24 | 933.2303 | 20 | −0.4 | |
| 28 | 6.142 | Quercetin feruloyl trihexoside (2) | C43H48O25 | 963.2393 | 20 | −0.5 | |
| 29 | 6.209 | Isorhamnetin dihexoside | C28H32O16 | 623.1623 | 13 | 0.9 | |
| 30 | 6.338 | Kaempferol feruloyl trihexoside | C43H48O24 | 947.2463 | 20 | 0 | |
| 31 | 6.941 | Quercetin deoxyhexoside | C21H20O11 | 447.0942 | 12 | 2.0 | |
| 32 | 7.525 | Kaempferol deoxyhexoside | C21H20O10 | 431.0993 | 12 | 2.2 |
| Peak | Rt | Tentative Assignment | Molecular Formula | [M-H]− Found | [M-H]2− Found | RDB | Error (ppm) |
|---|---|---|---|---|---|---|---|
| 1′ | 7.734 | 3- hexosyl deoxyhexosyl hederagenin 28-hexosyl ester | C48H78O18 | 941.5114 | 10 | −0.1 | |
| 2′ | 8.061 | 3-hexosyl pentosyl hederagenin 28-hexosyl ester | C47H76O18 | 927.4963 | 10 | 0.4 | |
| 3′ | 8.350 | 3-hexosyl deoxyhexosyl pentosyl hederagenin 28-dihexosyl ester | C61H100O31 | 1235.6059 | 663.3057 | 10 | 0.8 |
| 4′ | 8.490 | 3-deoxyhexosyl pentosyl hederagenin 28-hexosyl ester | C47H76O17 | 911.5018 | 10 | 0.9 | |
| 5′ | 8.928 | 3-hexosyl deoxyhexosyl pentosyl oleanolic acid | C47H76O16 | 895.5038 | 10 | −1.4 | |
| 6′ | 9.140 | 3-deoxyhexosyl pentosyl oleanolic acid 28-hexosyl ester | C47H76O16 | 895.5055 | 10 | −0.6 | |
| 7′ | 9.416 | 3-deoxyhexosyl pentosyl oleanolic acid 28-dihexosyl ester | C53H86O21 | 1057.5585 | 11 | −0.4 | |
| 8′ | 9.813 | 3-hexosyl deoxyhexosyl hederagenin 28-hexosyl ester | C48H78O18 | 941.5119 | 10 | 0.4 | |
| 9′ | 10.365 | 3-hexosyl deoxyhexosyl hederagenin pentosyl ester (1) | C47H76O17 | 911.5000 | 10 | −1.1 | |
| 10′ | 10.502 | 3-hexosyl deoxyhexosyl hederagenin pentosyl ester (2) | C47H76O17 | 911.5017 | 10 | 0.8 | |
| 11′ | 10.769 | 3-hexosyl deoxyhexosyl dehydrohederagenin pentosyl ester | C47H74O17 | 909.4855 | 10 | 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scognamiglio, M.; Maresca, V.; Basile, A.; Pacifico, S.; Fiorentino, A.; Bruno, M.; Badalamenti, N.; Kapelusz, M.; Marino, P.; Capasso, L.; et al. Phytochemical Characterization, Antioxidant, and Anti-Proliferative Activities of Wild and Cultivated Nigella damascena Species Collected in Sicily (Italy). Antioxidants 2024, 13, 402. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040402
Scognamiglio M, Maresca V, Basile A, Pacifico S, Fiorentino A, Bruno M, Badalamenti N, Kapelusz M, Marino P, Capasso L, et al. Phytochemical Characterization, Antioxidant, and Anti-Proliferative Activities of Wild and Cultivated Nigella damascena Species Collected in Sicily (Italy). Antioxidants. 2024; 13(4):402. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040402
Chicago/Turabian StyleScognamiglio, Monica, Viviana Maresca, Adriana Basile, Severina Pacifico, Antonio Fiorentino, Maurizio Bruno, Natale Badalamenti, Marta Kapelusz, Pasquale Marino, Lucia Capasso, and et al. 2024. "Phytochemical Characterization, Antioxidant, and Anti-Proliferative Activities of Wild and Cultivated Nigella damascena Species Collected in Sicily (Italy)" Antioxidants 13, no. 4: 402. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13040402