Involvement of Glutaredoxin and Thioredoxin Systems in the Nitrogen-Fixing Symbiosis between Legumes and Rhizobia

Abstract
:1. Introduction
2. The Glutaredoxin and Thioredoxin Systems of Plant Partner
2.1. The Glutaredoxin System
2.2. The Thioredoxin System
3. The Glutaredoxin and Thioredoxin Systems of Bacterial Partner
3.1. The Gutaredoxin System
3.2. The Thioredoxin System
3.3. Transcriptional Regulation of Trx and Grx Systems in S. meliloti
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, W.; Shi, J.; Xie, Q.; Jiang, Y.; Yu, N.; Wang, E. Nutrient exchange and regulation in arbuscular mycorrhizal symbiosis. Mol. Plant 2017, 10, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.M.; Uroz, S.; Barker, D.G. Ancestral alliances: Plant mutualistic symbioses with fungi and bacteria. Science 2017, 356. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, J.; Zhu, H. Genetic and Molecular Mechanisms Underlying Symbiotic Specificity in Legume-Rhizobium Interactions. Front. Plant Sci. 2018, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E. Speak, friend, and enter: Signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Maroti, G.; Kondorosi, E. Nitrogen-fixing Rhizobium-legume symbiosis: Are polyploidy and host peptide-governed symbiont differentiation general principles of endosymbiosis? Front. Microbiol. 2014, 5, 326. [Google Scholar] [PubMed]
- Hirsch, A.M. Developmental biology of legume nodulation. New Phytol. 1992, 122, 211–237. [Google Scholar] [CrossRef] [Green Version]
- Pierre, O.; Hopkins, J.; Combier, M.; Baldacci, F.; Engler, G.; Brouquisse, R.; Herouart, D.; Boncompagni, E. Involvement of papain and legumain proteinase in the senescence process of Medicago truncatula nodules. New Phytol. 2014, 202, 849–863. [Google Scholar] [CrossRef]
- Kondorosi, E.; Kondorosi, A. Endoreduplication and activation of the anaphase-promoting complex during symbiotic cell development. FEBS Lett. 2004, 567, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Vinardell, J.M.; Fedorova, E.; Cebolla, A.; Kevei, Z.; Horvath, G.; Kelemen, Z.; Tarayre, S.; Roudier, F.; Mergaert, P.; Kondorosi, A. Endoreduplication mediated by the anaphase-promoting complex activator CCS52A is required for symbiotic cell differentiation in Medicago truncatula nodules. Plant Cell 2003, 15, 2093–2105. [Google Scholar] [CrossRef]
- Kondorosi, E.; Mergaert, P.; Kereszt, A. A paradigm for endosymbiotic life: Cell differentiation of Rhizobium bacteria provoked by host plant factors. Annu. Rev. Microbiol. 2013, 67, 611–628. [Google Scholar] [CrossRef]
- Mergaert, P.; Nikovics, K.; Kelemen, Z.; Maunoury, N.; Vaubert, D.; Kondorosi, A.; Kondorosi, E. A novel family in Medicago truncatula consisting of more than 300 nodule-specific genes coding for small, secreted polypeptides with conserved cysteine motifs. Plant Physiol. 2003, 132, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Horvath, B.; Domonkos, A.; Kereszt, A.; Szucs, A.; Abraham, E.; Ayaydin, F.; Boka, K.; Chen, Y.; Chen, R.; Murray, J.D.; et al. Loss of the nodule-specific cysteine rich peptide, NCR169, abolishes symbiotic nitrogen fixation in the Medicago truncatula dnf7 mutant. Proc. Natl. Acad. Sci. USA 2015, 112, 15232–15237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Chen, Y.; Xi, J.; Waters, C.; Chen, R.; Wang, D. An antimicrobial peptide essential for bacterial survival in the nitrogen-fixing symbiosis. Proc. Natl. Acad. Sci. USA 2015, 112, 15238–15243. [Google Scholar] [CrossRef] [PubMed]
- Puppo, A.; Pauly, N.; Boscari, A.; Mandon, K.; Brouquisse, R. Hydrogen peroxide and nitric oxide: Key regulators of the Legume-Rhizobium and mycorrhizal symbioses. Antioxid. Redox Signal. 2013, 18, 2202–2219. [Google Scholar] [CrossRef] [PubMed]
- Hichri, I.; Boscari, A.; Castella, C.; Rovere, M.; Puppo, A.; Brouquisse, R. Nitric oxide: A multifaceted regulator of the nitrogen-fixing symbiosis. J. Exp. Bot. 2015, 66, 2877–2887. [Google Scholar] [CrossRef]
- Frendo, P.; Matamoros, M.A.; Alloing, G.; Becana, M. Thiol-based redox signaling in the nitrogen-fixing symbiosis. Front. Plant Sci. 2013, 4, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, C.W.; Alloing, G.; Mandon, K.; Frendo, P. Redox regulation of differentiation in symbiotic nitrogen fixation. Biochim. Biophys. Acta 2015, 1850, 1469–1478. [Google Scholar] [CrossRef]
- Klapheck, S. Homoglutathione: Isolation, quantification and occurrence in legumes. Physiol. Plant. 1988, 74, 727–732. [Google Scholar] [CrossRef]
- Frendo, P.; Jimenez, M.J.; Mathieu, C.; Duret, L.; Gallesi, D.; Van de Sype, G.; Herouart, D.; Puppo, A. A Medicago truncatula homoglutathione synthetase is derived from glutathione synthetase by gene duplication. Plant Physiol. 2001, 126, 1706–1715. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Clemente, M.R.; Sato, S.; Asamizu, E.; Tabata, S.; Ramos, J.; Moran, J.F.; Stiller, J.; Gresshoff, P.M.; Becana, M. Molecular analysis of the pathway for the synthesis of thiol tripeptides in the model legume Lotus japonicus. Mol. Plant Microbe Interact. 2003, 16, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Matamoros, M.A.; Moran, J.F.; Iturbe-Ormaetxe, I.; Rubio, M.C.; Becana, M. Glutathione and homoglutathione synthesis in legume root nodules. Plant Physiol. 1999, 121, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, L.R.H. Glutathione metabolism in plants. In Sulphur Nutrition and Assimilation in Higher Plants; DeKok, S.I., Rennenberg, H., Brunold, C., Rauser, W., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1993; pp. 109–124. [Google Scholar]
- Meyer, A.J.; May, M.J.; Fricker, M. Quantitative in vivo measurement of glutathione in Arabidopsis cells. Plant J. 2001, 27, 67–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalloniati, C.; Krompas, P.; Karalias, G.; Udvardi, M.K.; Rennenberg, H.; Herschbach, C.; Flemetakis, E. Nitrogen-fixing nodules are an important source of reduced sulfur, which triggers global changes in sulfur metabolism in Lotus japonicus. Plant Cell 2015, 27, 2384–2400. [Google Scholar] [CrossRef] [PubMed]
- Thal, B.; Braun, H.P.; Eubel, H. Proteomic analysis dissects the impact of nodulation and biological nitrogen fixation on Vicia faba root nodule physiology. Plant Mol. Biol. 2018, 97, 233–251. [Google Scholar] [CrossRef] [PubMed]
- Krusell, L.; Krause, K.; Ott, T.; Desbrosses, G.; Kramer, U.; Sato, S.; Nakamura, Y.; Tabata, S.; James, E.K.; Sandal, N.; et al. The sulfate transporter SST1 is crucial for symbiotic nitrogen fixation in Lotus japonicus root nodules. Plant Cell 2005, 17, 1625–1636. [Google Scholar] [CrossRef]
- Wienkoop, S.; Saalbach, G. Proteome analysis. Novel proteins identified at the peribacteroid membrane from Lotus japonicus root nodules. Plant Physiol. 2003, 131, 1080–1090. [Google Scholar] [CrossRef]
- Clarke, V.C.; Loughlin, P.C.; Gavrin, A.; Chen, C.; Brear, E.M.; Day, D.A.; Smith, P.M. Proteomic analysis of the soybean symbiosome identifies new symbiotic proteins. Mol. Cell Proteom. 2015, 14, 1301–1322. [Google Scholar] [CrossRef]
- Macnicol, P.K.; Randall, P.J. Changes in the levels of major sulfur metabolites and free amino acids in pea cotyledons recovering from sulfur deficiency. Plant Physiol. 1987, 83, 354–359. [Google Scholar] [CrossRef]
- Iturbe-Ormaetxe, I.; Heras, B.; Matamoros, M.A.; Ramos, J.; Moran, J.F.; Becana, M. Cloning and functional characterization of a homoglutathione synthetase from pea nodules. Physiol. Plant 2002, 115, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Galant, A.; Preuss, M.; Cameron, J.; Jez, J. Plant Glutathione Biosynthesis: Diversity in Biochemical Regulation and Reaction Products. Front. Plant Sci. 2011, 2, 45. [Google Scholar] [CrossRef]
- Frendo, P.; Mathieu, C.; Van de Sype, G.; Herouart, D.; Puppo, A. Characterisation of a cDNA encoding gamma-glutamylcysteine synthetase in Medicago truncatula. Free Radic. Res. 1999, 31, S213–S218. [Google Scholar] [CrossRef]
- Dalton, D.A.; Baird, L.M.; Langeberg, L.; Taugher, C.Y.; Anyan, W.R.; Vance, C.P.; Sarath, G. Subcellular localization of oxygen defense enzymes in Soybean (Glycine max [L.] Merr.) root nodules. Plant Physiol. 1993, 102, 481–489. [Google Scholar] [CrossRef]
- Evans, P.J.; Gallesi, D.; Mathieu, C.; Hernandez, M.J.; de Felipe, M.; Halliwell, B.; Puppo, A. Oxidative stress occurs during soybean nodule senescence. Planta 1999, 208, 73–79. [Google Scholar] [CrossRef]
- Groten, K.; Vanacker, H.; Dutilleul, C.; Bastian, F.; Bernard, S.; Carzaniga, R.; Foyer, C.H. The roles of redox processes in pea nodule development and senescence. Plant Cell Environ. 2005, 28, 1293–1304. [Google Scholar] [CrossRef] [Green Version]
- Escuredo, P.R.; Minchin, F.R.; Gogorcena, Y.; Iturbe-Ormaetxe, I.; Klucas, R.V.; Becana, M. Involvement of activated oxygen in nitrate-Induced senescence of pea root nodules. Plant Physiol. 1996, 110, 1187–1195. [Google Scholar] [CrossRef]
- Gogorcena, Y.; Gordon, A.J.; Escuredo, P.R.; Minchin, F.R.; Witty, J.F.; Moran, J.F.; Becana, M. N2 fixation, carbon metabolism and oxidative damage in nodules of dark stressed common bean plants. Plant Physiol. 1997, 113, 1193–1201. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Baird, L.M.; Escuredo, P.R.; Dalton, D.A.; Minchin, F.R.; Iturbe-Ormaetxe, I.; Rubio, M.C.; Moran, J.F.; Gordon, A.J.; Becana, M. Stress induced legume root nodule senescence. Physiological, biochemical and structural alterations. Plant Physiol. 1999, 121, 97–112. [Google Scholar] [CrossRef]
- Marino, D.; Frendo, P.; Ladrera, R.; Zabalza, A.; Puppo, A.; Arrese-Igor, C.; Gonzalez, E.M. Nitrogen fixation control under drought stress. Localized or systemic? Plant Physiol. 2007, 143, 1968–1974. [Google Scholar] [CrossRef]
- Naya, L.; Ladrera, R.; Ramos, J.; Gonzalez, E.M.; Arrese-Igor, C.; Minchin, F.R.; Becana, M. The response of carbon metabolism and antioxidant defenses of alfalfa nodules to drought stress and to the subsequent recovery of plants. Plant Physiol. 2007, 144, 1104–1114. [Google Scholar] [CrossRef]
- Frendo, P.; Harrison, J.; Norman, C.; Hernandez Jimenez, M.J.; Van de Sype, G.; Gilabert, A.; Puppo, A. Glutathione and homoglutathione play a critical role in the nodulation process of Medicago truncatula. Mol. Plant Microbe Interact. 2005, 18, 254–259. [Google Scholar] [CrossRef]
- Vernoux, T.; Wilson, R.C.; Seeley, K.A.; Reichheld, J.P.; Muroy, S.; Brown, S.; Maughan, S.C.; Cobbett, C.S.; Van Montagu, M.; Inze, D.; et al. The ROOT MERISTEMLESS1/CADMIUM SENSITIVE2 gene defines a glutathione-dependent pathway involved in initiation and maintenance of cell division during postembryonic root development. Plant Cell 2000, 12, 97–110. [Google Scholar] [CrossRef]
- Reichheld, J.P.; Khafif, M.; Riondet, C.; Droux, M.; Bonnard, G.; Meyer, Y. Inactivation of thioredoxin reductases reveals a complex interplay between thioredoxin and glutathione pathways in Arabidopsis development. Plant Cell 2007, 19, 1851–1865. [Google Scholar] [CrossRef]
- Schippers, J.H.; Foyer, C.H.; van Dongen, J.T. Redox regulation in shoot growth, SAM maintenance and flowering. Curr. Opin. Plant Biol. 2016, 29, 121–128. [Google Scholar] [CrossRef]
- Pucciariello, C.; Innocenti, G.; Van de Velde, W.; Lambert, A.; Hopkins, J.; Clement, M.; Ponchet, M.; Pauly, N.; Goormachtig, S.; Holsters, M.; et al. (Homo)glutathione depletion modulates host gene expression during the symbiotic interaction between Medicago truncatula and Sinorhizobium meliloti. Plant Physiol. 2009, 151, 1186–1196. [Google Scholar] [CrossRef]
- El Msehli, S.; Lambert, A.; Baldacci-Cresp, F.; Hopkins, J.; Boncompagni, E.; Smiti, S.A.; Herouart, D.; Frendo, P. Crucial role of (homo)glutathione in nitrogen fixation in Medicago truncatula nodules. New Phytol. 2011, 192, 496–506. [Google Scholar] [CrossRef]
- Rouhier, N.; Couturier, J.; Johnson, M.K.; Jacquot, J.P. Glutaredoxins: Roles in iron homeostasis. Trends Biochem. Sci. 2010, 35, 43–52. [Google Scholar] [CrossRef]
- Meyer, Y.; Belin, C.; Delorme-Hinoux, V.; Reichheld, J.P.; Riondet, C. Thioredoxin and glutaredoxin systems in plants: Molecular mechanisms, crosstalks, and functional significance. Antioxid. Redox Signal. 2012, 17, 1124–1160. [Google Scholar] [CrossRef]
- Roux, B.; Rodde, N.; Jardinaud, M.F.; Timmers, T.; Sauviac, L.; Cottret, L.; Carrère, S.; Sallet, E.; Courcelle, E.; Moreau, S.; et al. An integrated analysis of plant and bacterial gene expression in symbiotic root nodules using laser-capture microdissection coupled to RNA sequencing. Plant J. 2014, 77, 817–837. [Google Scholar] [CrossRef] [Green Version]
- Gelhaye, E.; Rouhier, N.; Gerard, J.; Jolivet, Y.; Gualberto, J.; Navrot, N.; Ohlsson, P.I.; Wingsle, G.; Hirasawa, M.; Knaff, D.B.; et al. A specific form of thioredoxin h occurs in plant mitochondria and regulates the alternative oxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 14545–14550. [Google Scholar] [CrossRef] [Green Version]
- Renard, M.; Alkhalfioui, F.; Schmitt-Keichinger, C.; Ritzenthaler, C.; Montrichard, F. Identification and characterization of thioredoxin h isoforms differentially expressed in germinating seeds of the model legume Medicago truncatula. Plant Physiol. 2011, 155, 1113–1126. [Google Scholar] [CrossRef] [Green Version]
- Meyer, Y.; Buchanan, B.B.; Vignols, F.; Reichheld, J.P. Thioredoxins and glutaredoxins: Unifying elements in redox biology. Annu. Rev. Genet. 2009, 43, 335–367. [Google Scholar] [CrossRef]
- Pulido, P.; Cazalis, R.; Cejudo, F.J. An antioxidant redox system in the nucleus of wheat seed cells suffering oxidative stress. Plant J. 2009, 57, 132–145. [Google Scholar] [CrossRef] [Green Version]
- Marchal, C.; Delorme-Hinoux, V.; Bariat, L.; Siala, W.; Belin, C.; Saez-Vasquez, J.; Riondet, C.; Reichheld, J.P. NTR/NRX define a new thioredoxin system in the nucleus of Arabidopsis thaliana cells. Mol. Plant 2014, 7, 30–44. [Google Scholar] [CrossRef]
- Alkhalfioui, F.; Renard, M.; Frendo, P.; Keichinger, C.; Meyer, Y.; Gelhaye, E.; Hirasawa, M.; Knaff, D.B.; Ritzenthaler, C.; Montrichard, F. A novel type of thioredoxin dedicated to symbiosis in legumes. Plant Physiol. 2008, 148, 424–435. [Google Scholar] [CrossRef]
- Tovar-Mendez, A.; Matamoros, M.A.; Bustos-Sanmamed, P.; Dietz, K.J.; Cejudo, F.J.; Rouhier, N.; Sato, S.; Tabata, S.; Becana, M. Peroxiredoxins and NADPH-dependent thioredoxin systems in the model legume Lotus japonicus. Plant Physiol. 2011, 156, 1535–1547. [Google Scholar] [CrossRef]
- Lee, M.Y.; Shin, K.H.; Kim, Y.K.; Suh, J.Y.; Gu, Y.Y.; Kim, M.R.; Hur, Y.S.; Son, O.; Kim, J.S.; Song, E.; et al. Induction of thioredoxin is required for nodule development to reduce reactive oxygen species levels in soybean roots. Plant Physiol. 2005, 139, 1881–1889. [Google Scholar] [CrossRef]
- Du, H.; Kim, S.; Nam, K.H.; Lee, M.S.; Son, O.; Lee, S.H.; Cheon, C.I. Identification of uricase as a potential target of plant thioredoxin: Implication in the regulation of nodule development. Biochem. Biophys. Res. Commun. 2010, 397, 22–26. [Google Scholar] [CrossRef]
- Ribeiro, C.W.; Baldacci-Cresp, F.; Pierre, O.; Larousse, M.; Benyamina, S.; Lambert, A.; Hopkins, J.; Castella, C.; Cazareth, J.; Alloing, G.; et al. Regulation of differentiation of nitrogen-fixing bacteria by microsymbiont targeting of plant thioredoxin s1. Curr. Biol. 2017, 27, 250–256. [Google Scholar] [CrossRef]
- Rouhier, N.; Jacquot, J.P. The plant multigenic family of thiol peroxidases. Free Radic. Biol. Med. 2005, 38, 1413–1421. [Google Scholar] [CrossRef]
- Castella, C.; Mirtziou, I.; Seassau, A.; Boscari, A.; Montrichard, F.; Papadopoulou, K.; Rouhier, N.; Puppo, A.; Brouquisse, R. Post-translational modifications of Medicago truncatula glutathione peroxidase 1 induced by nitric oxide. Nitric Oxide 2017, 68, 125–136. [Google Scholar] [CrossRef]
- Ramos, J.; Matamoros, M.A.; Naya, L.; James, E.K.; Rouhier, N.; Sato, S.; Tabata, S.; Becana, M. The glutathione peroxidase gene family of Lotus japonicus: Characterization of genomic clones, expression analyses and immunolocalization in legumes. New Phytol. 2009, 181, 103–114. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Saiz, A.; Penuelas, M.; Bustos-Sanmamed, P.; Mulet, J.M.; Barja, M.V.; Rouhier, N.; Moore, M.; James, E.K.; Dietz, K.J.; et al. Function of glutathione peroxidases in legume root nodules. J. Exp. Bot. 2015, 66, 2979–2990. [Google Scholar] [CrossRef] [Green Version]
- Matamoros, M.A.; Fernandez-Garcia, N.; Wienkoop, S.; Loscos, J.; Saiz, A.; Becana, M. Mitochondria are an early target of oxidative modifications in senescing legume nodules. New Phytol. 2013, 197, 873–885. [Google Scholar] [CrossRef] [Green Version]
- Groten, K.; Dutilleul, C.; van Heerden, P.D.; Vanacker, H.; Bernard, S.; Finkemeier, I.; Dietz, K.J.; Foyer, C.H. Redox regulation of peroxiredoxin and proteinases by ascorbate and thiols during pea root nodule senescence. FEBS Lett. 2006, 580, 1269–1276. [Google Scholar] [CrossRef] [Green Version]
- Harrison, J.; Jamet, A.; Muglia, C.I.; Van de Sype, G.; Aguilar, O.M.; Puppo, A.; Frendo, P. Glutathione plays a fundamental role in growth and symbiotic capacity of Sinorhizobium meliloti. J. Bacteriol. 2005, 187, 168–174. [Google Scholar] [CrossRef]
- Muglia, C.; Comai, G.; Spegazzini, E.; Riccillo, P.M.; Aguilar, O.M. Glutathione produced by Rhizobium tropici is important to prevent early senescence in common bean nodules. FEMS Microbiol. Lett. 2008, 23, 191–198. [Google Scholar] [CrossRef]
- Tate, R.; Cermola, M.; Riccio, A.; Diez-Roux, G.; Patriarca, E.J. Glutathione is required by Rhizobium etli for glutamine utilization and symbiotic effectiveness. Mol. Plant Microbe Interact. 2012, 25, 331–340. [Google Scholar] [CrossRef]
- Sobrevals, L.; Muller, P.; Fabra, A.; Castro, S. Role of glutathione in the growth of Bradyrhizobium sp. (peanut microsymbiont) under different environmental stresses and in symbiosis with the host plant. Can. J. Microbiol. 2006, 52, 609–616. [Google Scholar] [CrossRef]
- Tang, G.; Li, N.; Liu, Y.; Yu, L.; Yan, J.; Luo, L. Sinorhizobium meliloti Glutathione Reductase Is Required for both Redox Homeostasis and Symbiosis. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef]
- Cheng, G.; Karunakaran, R.; East, A.K.; Munoz-Azcarate, O.; Poole, P.S. Glutathione affects the transport activity of Rhizobium leguminosarum 3841 and is essential for efficient nodulation. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef]
- Benyamina, S.M.; Baldacci-Cresp, F.; Couturier, J.; Chibani, K.; Hopkins, J.; Bekki, A.; de Lajudie, P.; Rouhier, N.; Jacquot, J.P.; Alloing, G.; et al. Two Sinorhizobium meliloti glutaredoxins regulate iron metabolism and symbiotic bacteroid differentiation. Environ. Microbiol. 2013, 15, 795–810. [Google Scholar] [CrossRef]
- Spatzal, T.; Aksoyoglu, M.; Zhang, L.; Andrade, S.L.; Schleicher, E.; Weber, S.; Rees, D.C.; Einsle, O. Evidence for interstitial carbon in nitrogenase FeMo cofactor. Science 2011, 334, 940. [Google Scholar] [CrossRef]
- Sasaki, S.; Minamisawa, K.; Mitsui, H. A Sinorhizobium meliloti RpoH-regulated gene is involved in iron-sulfur protein metabolism and effective plant symbiosis under intrinsic iron limitation. J. Bacteriol. 2016, 198, 2297–2306. [Google Scholar] [CrossRef]
- Prinz, W.A.; Aslund, F.; Holmgren, A.; Beckwith, J. The role of the thioredoxin and glutaredoxin pathways in reducing protein disulfide bonds in the Escherichia coli cytoplasm. J. Biol. Chem. 1997, 272, 15661–15667. [Google Scholar] [CrossRef]
- Castro-Sowinski, S.; Matan, O.; Bonafede, P.; Okon, Y. A thioredoxin of Sinorhizobium meliloti CE52G is required for melanin production and symbiotic nitrogen fixation. Mol. Plant Microbe Interact. 2007, 20, 986–993. [Google Scholar] [CrossRef]
- Vargas, C.; Wu, G.; Davies, A.E.; Downie, J.A. Identification of a gene encoding a thioredoxin-like product necessary for cytochrome c biosynthesis and symbiotic nitrogen fixation in Rhizobium leguminosarum. J. Bacteriol. 1994, 176, 4117–4123. [Google Scholar] [CrossRef]
- Abicht, H.K.; Scharer, M.A.; Quade, N.; Ledermann, R.; Mohorko, E.; Capitani, G.; Hennecke, H.; Glockshuber, R. How periplasmic thioredoxin TlpA reduces bacterial copper chaperone ScoI and cytochrome oxidase subunit II (CoxB) prior to metallation. J. Biol. Chem. 2014, 289, 32431–32444. [Google Scholar] [CrossRef]
- Luo, L.; Yao, S.Y.; Becker, A.; Ruberg, S.; Yu, G.Q.; Zhu, J.B.; Cheng, H.P. Two new Sinorhizobium meliloti LysR-type transcriptional regulators required for nodulation. J. Bacteriol. 2005, 187, 4562–4572. [Google Scholar] [CrossRef]
- Imlay, J.A. Transcription factors that defend bacteria against reactive oxygen species. Annu. Rev. Microbiol. 2015, 69, 93–108. [Google Scholar] [CrossRef]
- Lu, D.; Tang, G.; Wang, D.; Luo, L. The Sinorhizobium meliloti LysR family transcriptional factor LsrB is involved in regulation of glutathione biosynthesis. Acta Biochim. Biophys. Sin. (Shanghai) 2013, 45, 882–888. [Google Scholar] [CrossRef]
- Tang, G.; Xing, S.; Wang, S.; Yu, L.; Li, X.; Staehelin, C.; Yang, M.; Luo, L. Regulation of cysteine residues in LsrB proteins from Sinorhizobium meliloti under free-living and symbiotic oxidative stress. Environ. Microbiol. 2017, 19, 5130–5145. [Google Scholar] [CrossRef]
- Tang, G.; Wang, Y.; Luo, L. Transcriptional regulator LsrB of Sinorhizobium meliloti positively regulates the expression of genes involved in lipopolysaccharide biosynthesis. Appl. Environ. Microbiol. 2014, 80, 5265–5273. [Google Scholar] [CrossRef]
- Ono, Y.; Mitsui, H.; Sato, T.; Minamisawa, K. Two RpoH homologs responsible for the expression of heat shock protein genes in Sinorhizobium meliloti. Mol. Gen. Genet. 2001, 264, 902–912. [Google Scholar] [CrossRef]
- De Lucena, D.K.; Puhler, A.; Weidner, S. The role of sigma factor RpoH1 in the pH stress response of Sinorhizobium meliloti. BMC Microbiol. 2010, 10, 265. [Google Scholar] [CrossRef]
- Barnett, M.J.; Bittner, A.N.; Toman, C.J.; Oke, V.; Long, S.R. Dual RpoH sigma factors and transcriptional plasticity in a symbiotic bacterium. J. Bacteriol. 2012, 194, 4983–4994. [Google Scholar] [CrossRef]
- Lehman, A.P.; Long, S.R. OxyR-dependent transcription response of Sinorhizobium meliloti to oxidative stress. J. Bacteriol. 2018, 200. [Google Scholar] [CrossRef]
- Mitsui, H.; Sato, T.; Sato, Y.; Ito, N.; Minamisawa, K. Sinorhizobium meliloti RpoH1 is required for effective nitrogen-fixing symbiosis with alfalfa. Mol. Genet. Genom. 2004, 271, 416–425. [Google Scholar] [CrossRef]
- Kereszt, A.; Mergaert, P.; Montiel, J.; Endre, G.; Kondorosi, E. Impact of plant peptides on symbiotic nodule development and functioning. Front. Plant Sci. 2018, 9, 1026. [Google Scholar] [CrossRef]
- Tiricz, H.; Szucs, A.; Farkas, A.; Pap, B.; Lima, R.M.; Maroti, G.; Kondorosi, E.; Kereszt, A. Antimicrobial nodule-specific cysteine-rich peptides induce membrane depolarization-associated changes in the transcriptome of Sinorhizobium meliloti. Appl. Environ. Microbiol. 2013, 79, 6737–6746. [Google Scholar] [CrossRef]
- Gon, S.; Faulkner, M.J.; Beckwith, J. In vivo requirement for glutaredoxins and thioredoxins in the reduction of the ribonucleotide reductases of Escherichia coli. Antioxid. Redox Signal. 2006, 8, 735–742. [Google Scholar] [CrossRef]
- Taga, M.E.; Walker, G.C. Sinorhizobium meliloti requires a cobalamin-dependent ribonucleotide reductase for symbiosis with its plant host. Mol. Plant Microbe Interact. 2010, 23, 1643–1654. [Google Scholar] [CrossRef] [Green Version]
- Ezraty, B.; Aussel, L.; Barras, F. Methionine sulfoxide reductases in prokaryotes. Biochim. Biophys. Acta 2005, 1703, 221–229. [Google Scholar] [CrossRef]
- Fontenelle, C.; Blanco, C.; Arrieta, M.; Dufour, V.; Trautwetter, A. Resistance to organic hydroperoxides requires ohr and ohrR genes in Sinorhizobium meliloti. BMC Microbiol. 2011, 11, 100. [Google Scholar] [CrossRef] [Green Version]
- Barranco-Medina, S.; Lazaro, J.J.; Dietz, K.J. The oligomeric conformation of peroxiredoxins links redox state to function. FEBS Lett. 2009, 583, 1809–1816. [Google Scholar] [CrossRef] [Green Version]
- Cussiol, J.R.; Alegria, T.G.; Szweda, L.I.; Netto, L.E. Ohr (organic hydroperoxide resistance protein) possesses a previously undescribed activity, lipoyl-dependent peroxidase. J. Biol. Chem. 2010, 285, 21943–21950. [Google Scholar] [CrossRef]
- Djordjevic, M.A.; Chen, H.C.; Natera, S.; Van Noorden, G.; Menzel, C.; Taylor, S.; Renard, C.; Geiger, O.; the Sinorhizobium DNA Sequencing Consortium; Weiller, G.F. A global analysis of protein expression profiles in Sinorhizobium meliloti: Discovery of new genes for nodule occupancy and stress adaptation. Mol. Plant Microbe Interact. 2003, 16, 508–524. [Google Scholar] [CrossRef]
- Dombrecht, B.; Heusdens, C.; Beullens, S.; Verreth, C.; Mulkers, E.; Proost, P.; Vanderleyden, J.; Michiels, J. Defence of Rhizobium etli bacteroids against oxidative stress involves a complexly regulated atypical 2-Cys peroxiredoxin. Mol. Microbiol. 2005, 55, 1207–1221. [Google Scholar] [CrossRef]



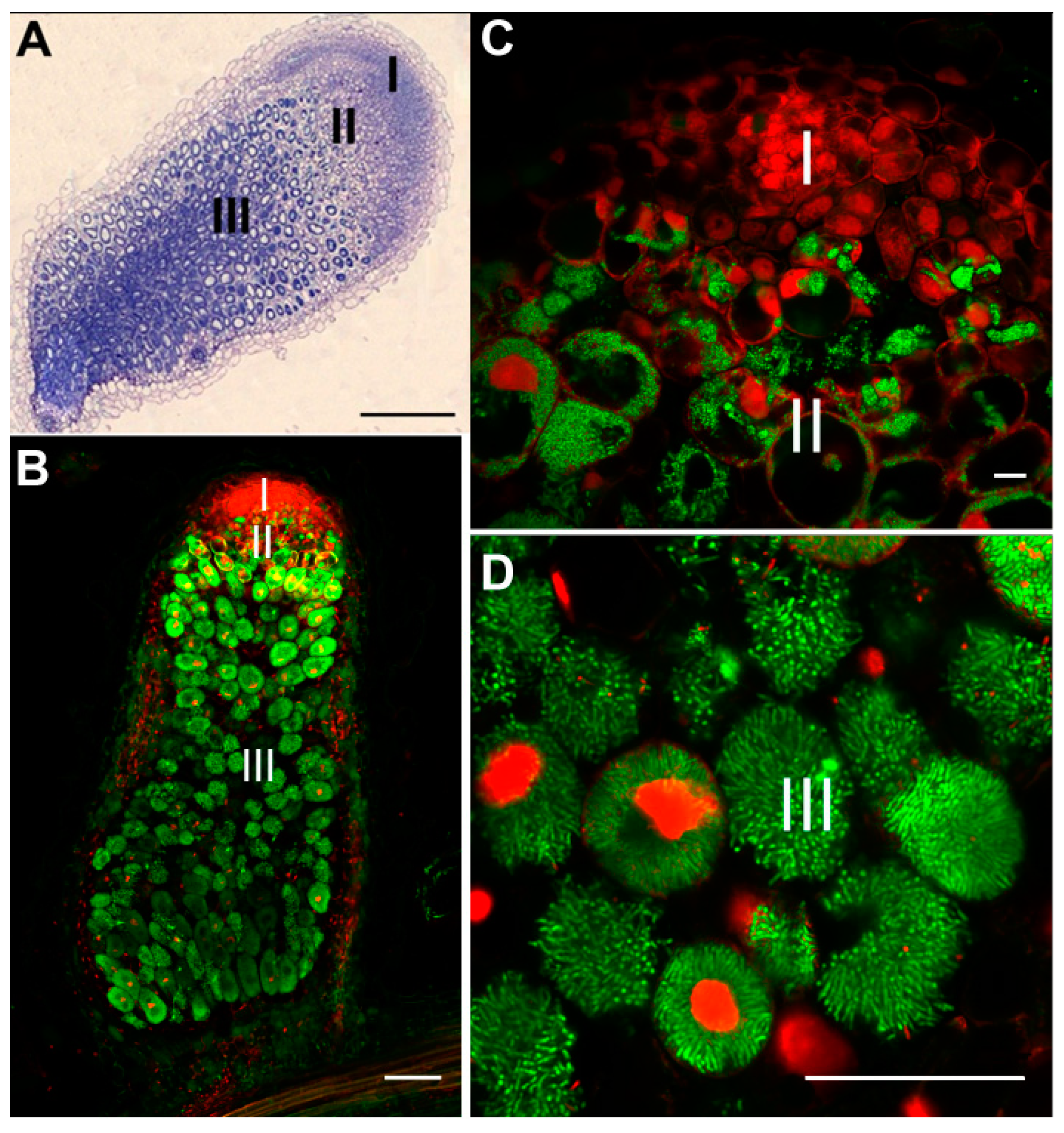{kind=link}
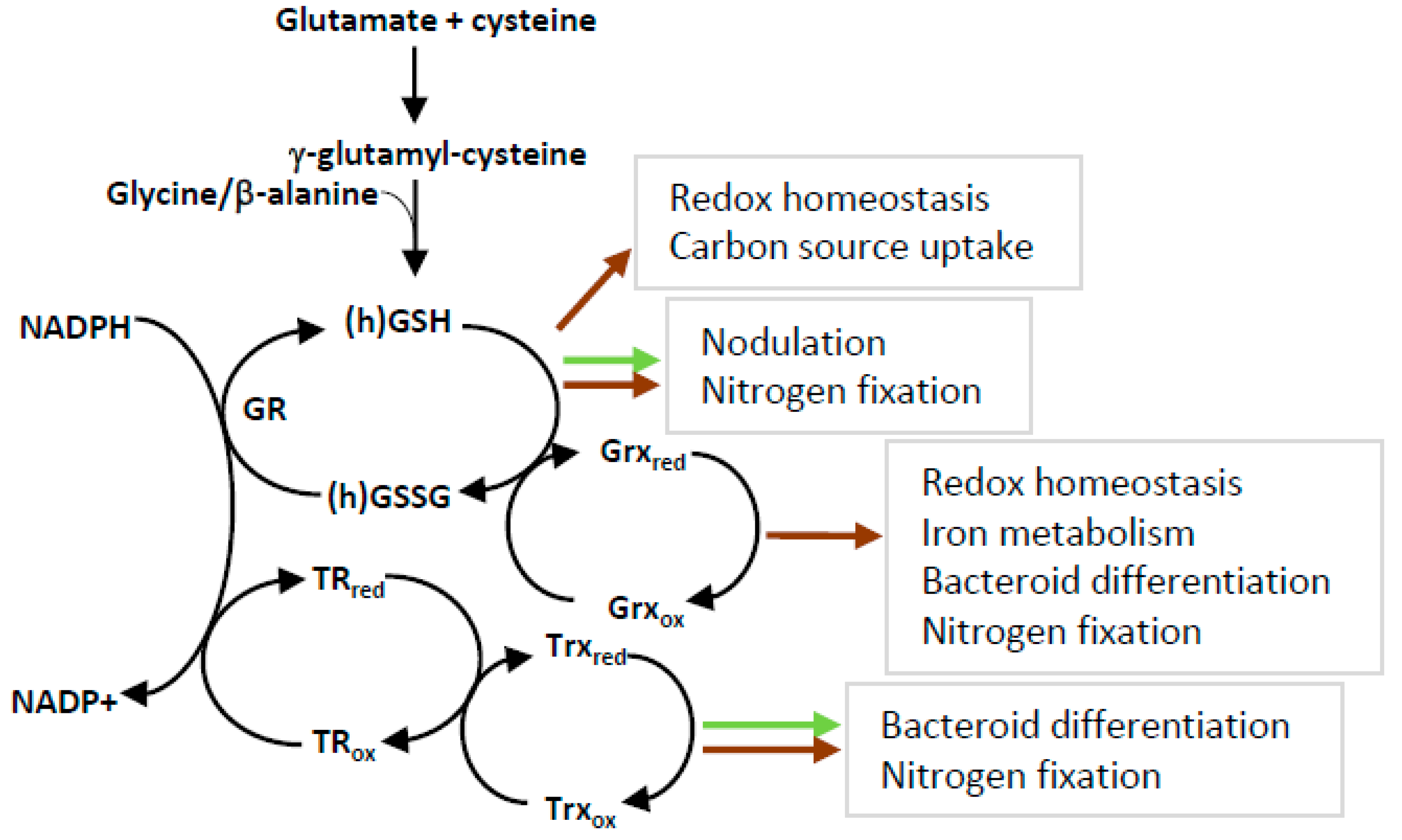{kind=link}
| Gene Name | Root | Nodule | I | IId | IIp | IZ II-III | III |
|---|---|---|---|---|---|---|---|
| Sulfate transporter | |||||||
| Medtr7g095430 | 1307 | 1297 | 7 | 2 | 4 | 19 | 9 |
| Medtr4g084620 | 298 | 176 | 13 | 9 | 43 | 44 | 29 |
| Mt0062_10115 | 54 | 548 | 30 | 154 | 289 | 235 | 361 |
| Medtr4g011970 | 633 | 1052 | 307 | 1721 | 3064 | 1094 | 1406 |
| Medtr3g087730 | 4 | 876 | 118 | 530 | 1088 | 323 | 92 |
| Medtr5g061860 | 87 | 1449 | 4 | 14 | 22 | 1313 | 1742 |
| Medtr6g086170 | 45 | 48,527 | 14 | 291 | 566 | 6898 | 12,537 |
| Medtr3g073730 | 220 | 20 | 2 | 4 | 1 | 1 | 0 |
| Medtr5g061880 | 200 | 43 | 15 | 14 | 2 | 3 | 3 |
| Medtr2g102243 | 322 | 193 | 19 | 18 | 1 | 1 | 0 |
| Medtr2g008470 | 1406 | 253 | 1 | 1 | 1 | 7 | 8 |
| Medtr3g087740 | 2150 | 854 | 19 | 17 | 16 | 4 | 13 |
| Mt0050_00072 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr4g084640 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mt0008_01149 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mt0008_11083 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr4g063825 | 0 | 6 | 1 | 0 | 0 | 0 | 0 |
| Mt0006_10002 | 1 | 0 | 2 | 3 | 0 | 0 | 0 |
| Medtr2g082610 | 3 | 5 | 1 | 2 | 0 | 0 | 0 |
| Medtr3g073780 | 9 | 1 | 0 | 0 | 0 | 0 | 1 |
| Medtr1g071530 | 345 | 184 | 2 | 2 | 0 | 1 | 1 |
| Medtr7g022870 | 464 | 449 | 48 | 52 | 17 | 20 | 39 |
| Gene name | Putative redox site | Root | Nodule | I | IId | IIp | IZ II-III | III |
|---|---|---|---|---|---|---|---|---|
| Glutaredoxins | ||||||||
| Class I | ||||||||
| Medtr7g035245 | YCPFC | 2612 | 2354 | 665 | 163 | 156 | 238 | 167 |
| Medtr1g069255 | WCSYC | 121 | 153 | 54 | 41 | 135 | 109 | 92 |
| Medtr3g077560 | YCGYC | 1314 | 366 | 2 | 1 | 1 | 2 | 0 |
| Medtr3g077570 | YCGYC | 201 | 148 | 27 | 16 | 3 | 3 | 15 |
| Medtr2g038560 | YCPYC | 1573 | 1200 | 425 | 682 | 837 | 509 | 297 |
| Medtr5g021090 | YCPYC | 1284 | 1444 | 76 | 97 | 63 | 72 | 50 |
| Class II | ||||||||
| Medtr2g103130 | QCGFS | 1332 | 1390 | 380 | 266 | 323 | 546 | 302 |
| Medtr4g079110 | GCCMS | 968 | 1834 | 62 | 36 | 1 | 0 | 0 |
| Medtr7g079520 | QCGFS | 771 | 640 | 170 | 143 | 103 | 107 | 62 |
| Medtr4g088905 | KCGFS | 1444 | 1940 | 277 | 252 | 194 | 97 | 105 |
| Medtr4g016930 | LCGSF | 124 | 218 | 57 | 66 | 70 | 65 | 71 |
| Class III | ||||||||
| Medtr7g026770 | TCCMC | 13 | 13 | 21 | 5 | 0 | 0 | 0 |
| Medtr3g104510 | SCCMC | 16 | 32 | 1 | 3 | 24 | 53 | 96 |
| Medtr1g088910 | SCYMC | 62 | 260 | 2 | 0 | 0 | 0 | 0 |
| Medtr1g015890 | SCCMC | 144 | 340 | 100 | 1437 | 562 | 164 | 258 |
| Medtr2g090755 | GCCMS | 78 | 387 | 10 | 4 | 1 | 2 | 3 |
| Medtr2g014760 | GCCLC | 71 | 467 | 33 | 19 | 8 | 4 | 10 |
| Medtr1g088925 | SCCLC | 474 | 1 | 1 | 1 | 0 | 0 | 0 |
| Medtr1g088920 | LCCLC | 460 | 3 | 1 | 0 | 0 | 0 | 0 |
| Medtr7g108200 | SCCLC | 1650 | 308 | 2 | 1 | 0 | 0 | 0 |
| Medtr4g119030 | SCCMS | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr2g048970 | SCCMS | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr2g019950 | SCGMS | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr4g119050 | SCCMS | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr7g108250 | TCCLS | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr7g108220 | SCYMC | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr7g108250 | TCCLS | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr7g022690 | SCCMC | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr5g077550 | DCCFS | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Medtr1g088905 | TCCLS | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Mt0001_10735 | SCCMS | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Medtr7g022710 | SCCMC | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Medtr7g022550 | SCCMC | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Medtr7g108260 | TCPMS | 2 | 4 | 0 | 0 | 0 | 0 | 0 |
| Medtr2g019900 | SCCMC | 16 | 84 | 0 | 0 | 0 | 0 | 0 |
| Medtr7g108210 | SCYMC | 30 | 30 | 1 | 0 | 0 | 0 | 0 |
| S. meliloti Genes | Bacterial Gene Expression in M. truncatula Nodules | Transcription Factors | Inducing Conditions | References | |||||
|---|---|---|---|---|---|---|---|---|---|
| Total reads LCM | % FI | % FIIp | % FIId | % IZ | % FIII | ||||
| Grx system | |||||||||
| SMc00825 (gshA) | 3819 | 16 | 22 | 30 | 16 | 16 | LsrB | GSSG | [82,83] |
| SMc00419 (gshB) | 6433 | 8 | 9 | 24 | 53 | 6 | LsrB, RpoH1 | [82,87] | |
| SMc00154 (gor) | 4477 | 19 | 12 | 28 | 24 | 17 | LsrB, RpoH1 | [82,87] | |
| SMc02443 (Smgrx1) | 9123 | 7 | 6 | 20 | 16 | 51 | RpoH1 | low pH | [86] |
| NCR247, NCR335 | [91] | ||||||||
| SMc00538 (Smgrx2) | 10138 | 24 | 24 | 21 | 22 | 9 | |||
| SMa0280 (Smgrx3) | 1571 | 17 | 18 | 22 | 22 | 21 | HS | [87] | |
| Trx system | |||||||||
| SMc02761 (trxA) | 5519 | 13 | 10 | 19 | 21 | 37 | RpoH1 | HS | [87] |
| NCR247, NCR335 | [91] | ||||||||
| SMc03801 | 2780 | 20 | 18 | 31 | 14 | 17 | RpoH1 | ||
| SMc01224 (trxB) | 5394 | 18 | 17 | 28 | 13 | 24 | RpoH1 | low pH, HS | [86,87] |
| Grx/Trx targets | |||||||||
| SMc02885 (msrA1) | 2016 | 11 | 8 | 17 | 32 | 33 | RpoH1 | low pH, HS | [86,87] |
| NCR247, NCR335 | [91] | ||||||||
| SMc02467 (msrA2) | 1690 | 5 | 14 | 22 | 43 | 16 | |||
| SMa1896 (msrA3) | 551 | 20 | 12 | 26 | 20 | 21 | RpoH1 | HS | [87] |
| NCR247, NCR335 | [91] | ||||||||
| H2O2 | [88] | ||||||||
| SMc00117 (msrB1) | 729 | 24 | 13 | 19 | 23 | 20 | RpoH1 | HS | [87] |
| SMa1894 (msrB2) | 107 | 0 | 14 | 17 | 24 | 45 | RpoH1 | HS | [87] |
| NCR247, NCR335 | [91] | ||||||||
| H2O2 | [88] | ||||||||
| SMc01724 (msrB3) | 795 | 0 | 19 | 23 | 18 | 40 | |||
| SMc01237 (nrdJ) | 33647 | 9 | 7 | 4 | 4 | 76 | |||
| SMb20964 | 1308 | 29 | 9 | 9 | 7 | 45 | OxyR | H2O2 | [88] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alloing, G.; Mandon, K.; Boncompagni, E.; Montrichard, F.; Frendo, P. Involvement of Glutaredoxin and Thioredoxin Systems in the Nitrogen-Fixing Symbiosis between Legumes and Rhizobia. Antioxidants 2018, 7, 182. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox7120182
Alloing G, Mandon K, Boncompagni E, Montrichard F, Frendo P. Involvement of Glutaredoxin and Thioredoxin Systems in the Nitrogen-Fixing Symbiosis between Legumes and Rhizobia. Antioxidants. 2018; 7(12):182. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox7120182
Chicago/Turabian StyleAlloing, Geneviève, Karine Mandon, Eric Boncompagni, Françoise Montrichard, and Pierre Frendo. 2018. "Involvement of Glutaredoxin and Thioredoxin Systems in the Nitrogen-Fixing Symbiosis between Legumes and Rhizobia" Antioxidants 7, no. 12: 182. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox7120182