NOX2ko Mice Show Largely Increased Expression of a Mutated NOX2 mRNA Encoding an Inactive NOX2 Protein

 , ,
, ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Mice and Approval of Animal Studies
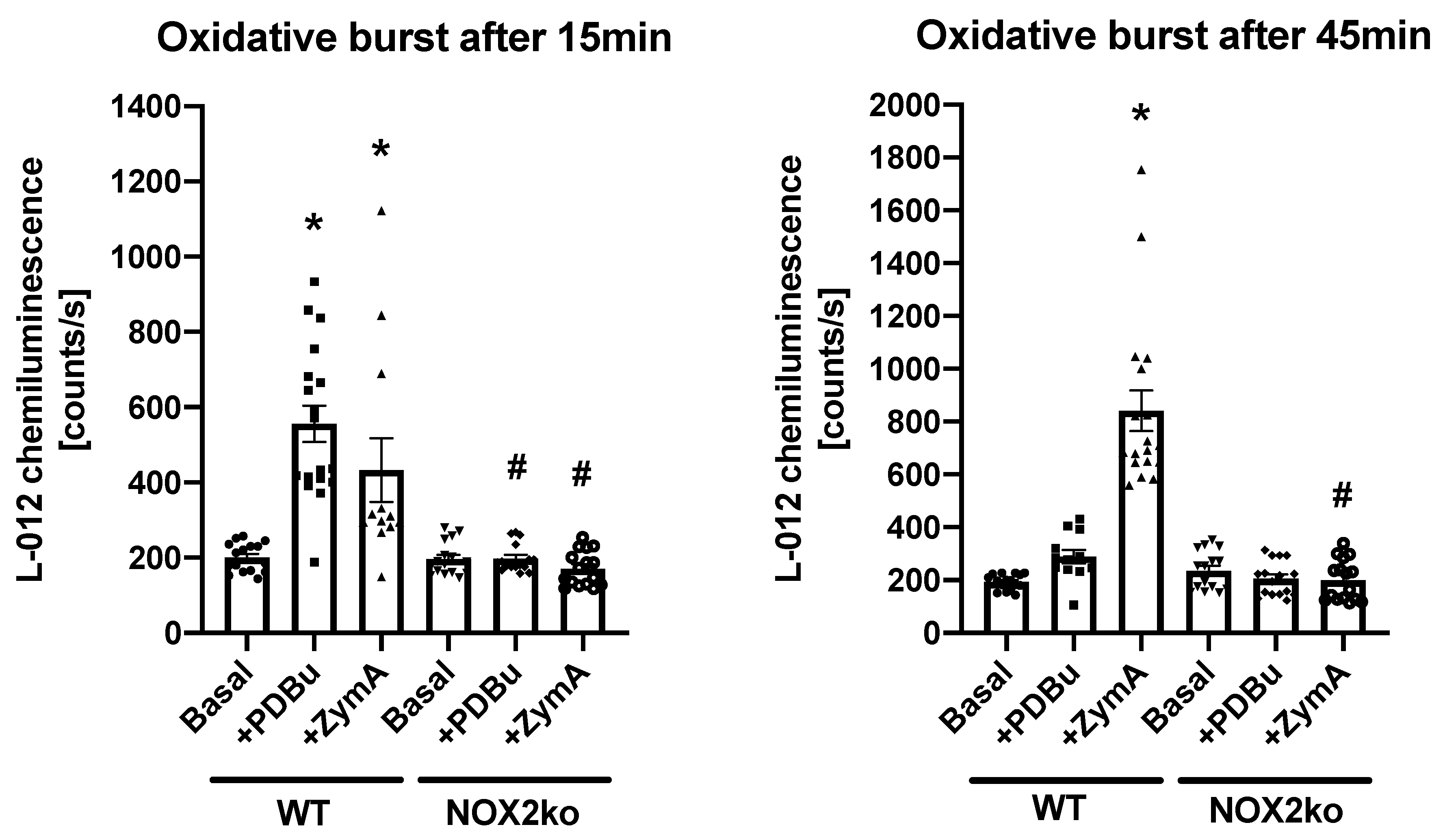
2.3. Measurement of Oxidative Burst in Whole Blood
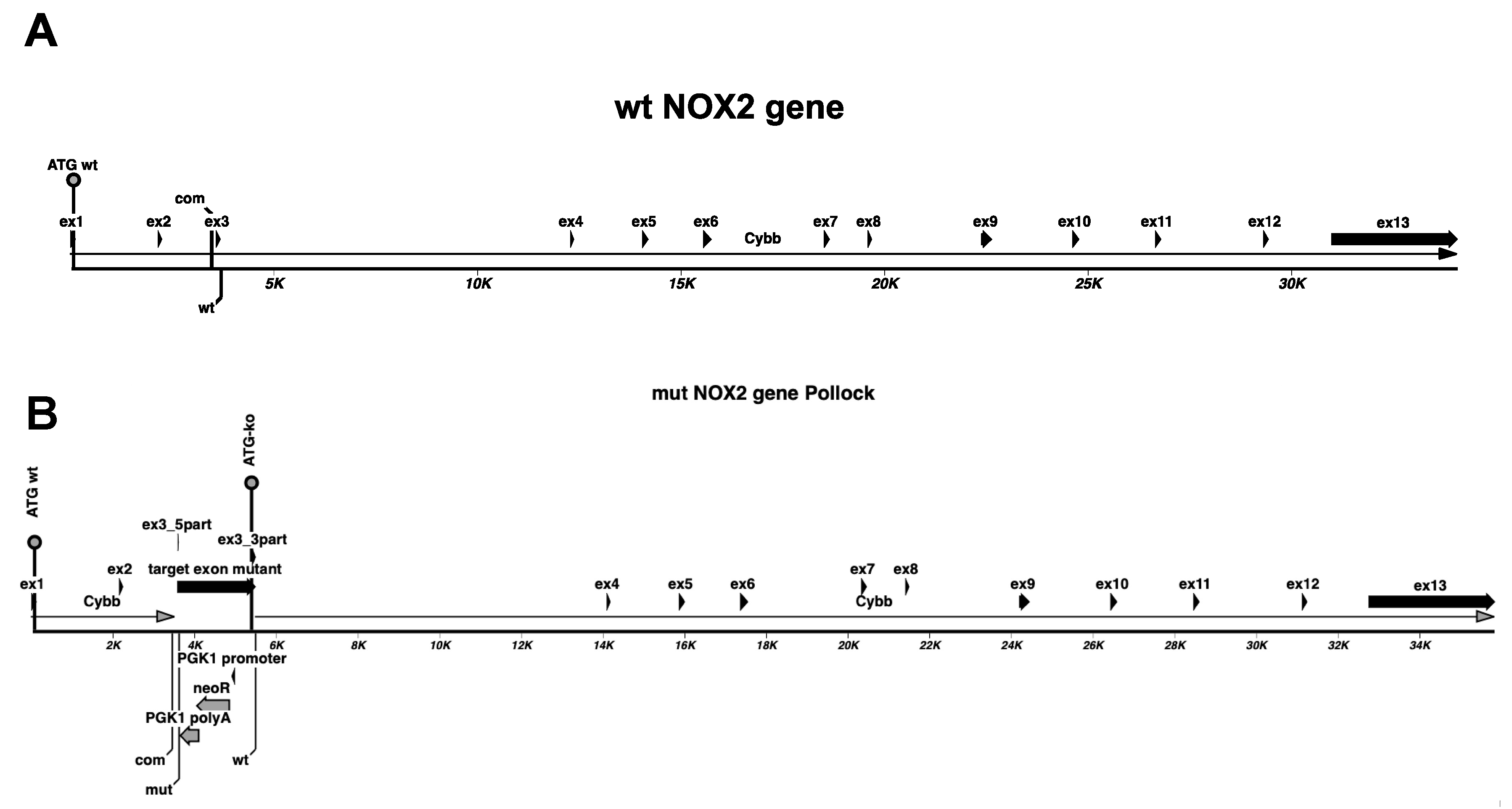
2.4. Analysis of NOX2ko mRNA by RT-PCR
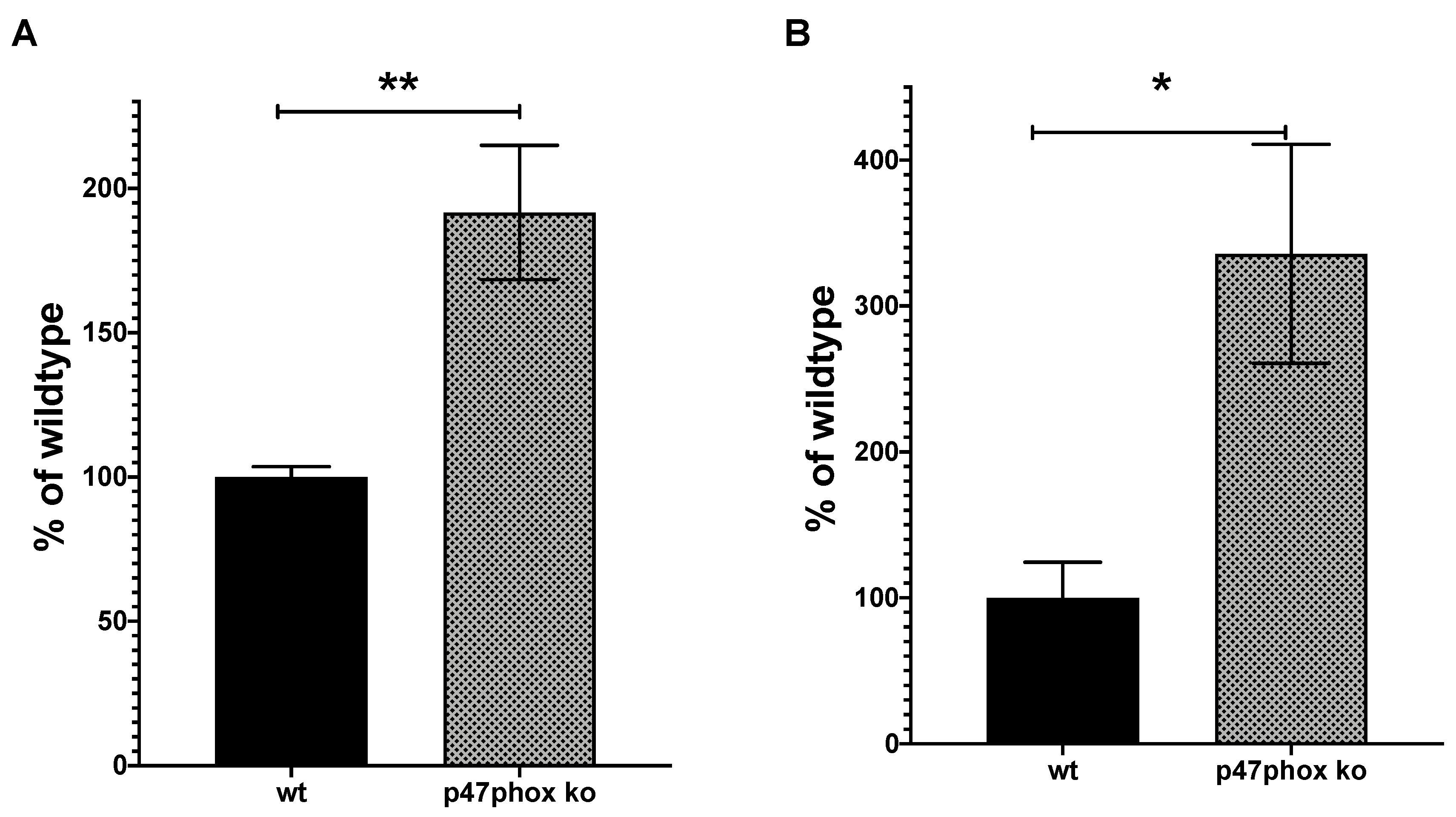
2.5. Analysis of NOX2ko mRNA Structure and Expression Values Using NGS Data
2.6. Real-Time Reverse Transcription Polymerase Chain Reaction Analysis
2.7. Western Blot Experiments
2.8. Immunohistochemistry
2.9. Statistics
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 3′- or 5′-UTR | 3′- or 5′-untranslated region |
| cds | coding sequence |
| ATG | translational start codon |
| Cybb | cybb/NOX2/gp9phox gene sequence |
| Ex | exon |
| neoR | neomycin resistance gene |
| PGK1 | phosphoglycerate kinase 1 |
| NOX2ko | mice with mutated NOX2 gene |
| NOX2wt | wildtype animals |
| PDBu | Phorbol 12,13-Dibutyrate |
| polyA | polyadenylation signal |
| qRT-PCR | quantitative real time reverse transcription polymerase chain reaction |
| WB | Western blot |
| ZymA | Zymosan A |
References
- Prieto-Bermejo, R.; Romo-González, M.; Pérez-Fernández, A.; Ijurko, C.; Hernández-Hernández, Á. Reactive oxygen species in haematopoiesis: Leukaemic cells take a walk on the wild side. J. Exp. Clin. Cancer Res. 2018, 37, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Egea, J.; Fabregat, I.; Frapart, Y.M.; Ghezzi, P.; Görlach, A.; Kietzmann, T.; Kubaichuk, K.; Knaus, U.G.; Lopez, M.G.; Olaso-Gonzalez, G.; et al. European contribution to the study of ROS: A summary of the findings and prospects for the future from the COST action BM1203 (EU-ROS). Redox Biol. 2017, 13, 94–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosaad, Y.M. Hematopoietic stem cells: An overview. Transfus. Apher. Sci. 2014, 51, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Valdivia, A.; Perez-Alvarez, S.; Aroca-Aguilar, J.-D.; Ikuta, I.; Jordan, J. Superoxide dismutases: A physiopharmacological update. J. Physiol. Biochem. 2009, 65, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Ewenzel, P.; Kossmann, S.; Münzel, T.; Daiber, A. Redox regulation of cardiovascular inflammation–Immunomodulatory function of mitochondrial and Nox-derived reactive oxygen and nitrogen species. Free Radic. Biol. Med. 2017, 109, 48–60. [Google Scholar] [CrossRef]
- Daiber, A.; Steven, S.; Weber, A.; Shuvaev, V.V.; Muzykantov, V.R.; Laher, I.; Li, H.; Lamas, S.; Münzel, T. Targeting vascular (endothelial) dysfunction. Br. J. Pharm. 2016, 174, 1591–1619. [Google Scholar] [CrossRef]
- Kauppila, T.E.; Kauppila, J.H.; Larsson, N.-G. Mammalian Mitochondria and Aging: An Update. Cell Metab. 2017, 25, 57–71. [Google Scholar] [CrossRef]
- Zeng, M.Y.; Miralda, I.; Armstrong, C.L.; Uriarte, S.M.; Bagaitkar, J. The roles of NADPH oxidase in modulating neutrophil effector responses. Mol. Oral Microbiol. 2019, 34, 27–38. [Google Scholar] [CrossRef]
- Daiber, A.; August, M.; Baldus, S.; Wendt, M.; Oelze, M.; Sydow, K.; Kleschyov, A.L.; Munzel, T. Measurement of NAD(P)H oxidase-derived superoxide with the luminol analogue L-012. Free Radic. Biol. Med. 2004, 36, 101–111. [Google Scholar] [CrossRef]
- Steven, S.; Oelze, M.; Hanf, A.; Kröller-Schön, S.; Kashani, F.; Roohani, S.; Welschof, P.; Kopp, M.; Gödtel-Armbrust, U.; Xia, N.; et al. The SGLT2 inhibitor empagliflozin improves the primary diabetic complications in ZDF rats. Redox Biol. 2017, 13, 370–385. [Google Scholar] [CrossRef]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef]
- Lassègue, B.; Martín, A.S.; Griendling, K.K. Biochemistry, Physiology, and Pathophysiology of NADPH Oxidases in the Cardiovascular System. Circ. Res. 2012, 110, 1364–1390. [Google Scholar] [CrossRef]
- Lambeth, J.D.; Neish, A.S. Nox Enzymes and New Thinking on Reactive Oxygen: A Double-Edged Sword Revisited. Annu. Rev. Pathol. Mech. Dis. 2014, 9, 119–145. [Google Scholar] [CrossRef]
- Altenhöfer, S.; Radermacher, K.A.; Kleikers, P.W.M.; Wingler, K.; Schmidt, H.H.H.W. Evolution of NADPH Oxidase Inhibitors: Selectivity and Mechanisms for Target Engagement. Antioxid. Redox Signal. 2015, 23, 406–427. [Google Scholar] [CrossRef]
- Kröller-Schön, S.; Daiber, A.; Steven, S.; Oelze, M.; Frenis, K.; Kalinovic, S.; Heimann, A.; Schmidt, F.P.; Pinto, A.; Kvandová, M.; et al. Crucial role for Nox2 and sleep deprivation in aircraft noise-induced vascular and cerebral oxidative stress, inflammation, and gene regulation. Eur. Heart J. 2018, 39, 3528–3539. [Google Scholar] [CrossRef]
- Stein, J.; Steven, S.; Bros, M.; Sudowe, S.; Hausding, M.; Oelze, M.; Münzel, T.; Grabbe, S.; Reske-Kunz, A.B.; Daiber, A. Role of Protein Kinase C and Nox2-Derived Reactive Oxygen Species Formation in the Activation and Maturation of Dendritic Cells by Phorbol Ester and Lipopolysaccharide. Oxidative Med. Cell. Longev. 2017, 2017, 1–12. [Google Scholar] [CrossRef]
- Brandt, M.; Garlapati, V.; Oelze, M.; Sotiriou, E.; Knorr, M.; Kröller-Schön, S.; Kossmann, S.; Schönfelder, T.; Morawietz, H.; Schulz, E.; et al. NOX2 amplifies acetaldehyde-mediated cardiomyocyte mitochondrial dysfunction in alcoholic cardiomyopathy. Sci. Rep. 2016, 6, 32554. [Google Scholar] [CrossRef]
- Kuntic, M.; Oelze, M.; Steven, S.; Kröller-Schön, S.; Stamm, P.; Kalinovic, S.; Frenis, K.; Vujacic-Mirski, K.; Jimenez, M.T.B.; Kvandova, M.; et al. Short-term e-cigarette vapour exposure causes vascular oxidative stress and dysfunction: Evidence for a close connection to brain damage and a key role of the phagocytic NADPH oxidase (NOX-2). Eur. Heart J. 2019, 41, 2472–2483. [Google Scholar] [CrossRef] [Green Version]
- Pollock, J.D.; Williams, D.A.; Gifford, M.A.; Li, L.L.; Du, X.; Fisherman, J.; Orkin, S.H.; Doerschuk, C.M.; Dinauer, M.C. Mouse model of X–linked chronic granulomatous disease, an inherited defect in phagocyte superoxide production. Nat. Genet. 1995, 9, 202–209. [Google Scholar] [CrossRef]
- Zielonka, J.; Lambeth, J.D.; Kalyanaraman, B. On the use of L-012, a luminol-based chemiluminescent probe, for detecting superoxide and identifying inhibitors of NADPH oxidase: A reevaluation. Free Radic. Biol. Med. 2013, 65, 1310–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oelze, M.; Kröller-Schön, S.; Welschof, P.; Jansen, T.; Hausding, M.; Mikhed, Y.; Stamm, P.; Mader, M.; Zinßius, E.; Agdauletova, S.; et al. The Sodium-Glucose Co-Transporter 2 Inhibitor Empagliflozin Improves Diabetes-Induced Vascular Dysfunction in the Streptozotocin Diabetes Rat Model by Interfering with Oxidative Stress and Glucotoxicity. PLoS ONE 2014, 9, e112394. [Google Scholar] [CrossRef] [PubMed]
- Bollmann, F.; Wu, Z.; Oelze, M.; Siuda, D.; Xia, N.; Henke, J.; Daiber, A.; Li, H.; Stumpo, D.J.; Blackshear, P.J.; et al. Endothelial Dysfunction in Tristetraprolin-deficient Mice Is Not Caused by Enhanced Tumor Necrosis Factor-α Expression. J. Biol. Chem. 2014, 289, 15653–15665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adane, B.; Ye, H.; Khan, N.; Pei, S.; Minhajuddin, M.; Stevens, B.M.; Jones, C.L.; D’Alessandro, A.; Reisz, J.A.; Zaberezhnyy, V.; et al. The Hematopoietic Oxidase NOX2 Regulates Self-Renewal of Leukemic Stem Cells. Cell Rep. 2019, 27, 238–254.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Münzel, T.; Daiber, A.; Steven, S.; Tran, L.P.; Ullmann, E.; Kossmann, S.; Schmidt, F.P.; Oelze, M.; Xia, N.; Li, H.; et al. Effects of noise on vascular function, oxidative stress, and inflammation: Mechanistic insight from studies in mice. Eur. Heart J. 2017, 38, 2838–2849. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Harrison, C.B.; Selemidis, S.; Guida, E.; King, P.T.; Sobey, C.G.; Drummond, G.R. Nox2beta: A novel splice variant of nox2 that regulates nadph oxidase activity in macrophages. PLoS ONE 2012, 7, e48326. [Google Scholar] [CrossRef]
- Jackson, S.H.; Gallin, J.I.; Holland, S.M. The p47phox mouse knock-out model of chronic granulomatous disease. J. Exp. Med. 1995, 182, 751–758. [Google Scholar] [CrossRef]
- Bendall, J.K.; Rinze, R.; Adlam, D.; Tatham, A.L.; De Bono, J.; Channon, K.M. Endothelial Nox2 Overexpression Potentiates Vascular Oxidative Stress and Hemodynamic Response to Angiotensin II. Circ. Res. 2007, 100, 1016–1025. [Google Scholar] [CrossRef] [Green Version]
- Murdoch, C.E.; Alom-Ruiz, S.P.; Wang, M.; Zhang, M.; Walker, S.; Yu, B.; Brewer, A.; Shah, A.M. Role of endothelial Nox2 NADPH oxidase in angiotensin II-induced hypertension and vasomotor dysfunction. Basic Res. Cardiol. 2011, 106, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Chrissobolis, S.; Bánfi, B.; Sobey, C.G.; Faraci, F.M. Role of Nox isoforms in angiotensin II-induced oxidative stress and endothelial dysfunction in brain. J. Appl. Physiol. 2012, 113, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dikalov, S.I.; Nazarewicz, R.R.; Bikineyeva, A.; Hilenski, L.; Lassègue, B.; Griendling, K.K.; Harrison, D.G.; Dikalova, A.E. Nox2-Induced Production of Mitochondrial Superoxide in Angiotensin II-Mediated Endothelial Oxidative Stress and Hypertension. Antioxid. Redox Signal. 2014, 20, 281–294. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, P.; Knorr, M.; Kossmann, S.; Stratmann, J.; Hausding, M.; Schuhmacher, S.; Karbach, S.H.; Schwenk, M.; Yogev, N.; Schulz, E.; et al. Lysozyme M–Positive Monocytes Mediate Angiotensin II–Induced Arterial Hypertension and Vascular Dysfunction. Circulation 2011, 124, 1370–1381. [Google Scholar] [CrossRef] [Green Version]
- Looi, Y.H.; Grieve, D.J.; Siva, A.; Walker, S.J.; Anilkumar, N.; Cave, A.C.; Marber, M.S.; Monaghan, M.J.; Shah, A.M. Involvement of Nox2 NADPH Oxidase in Adverse Cardiac Remodeling After Myocardial Infarction. Hypertension 2008, 51, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Somasuntharam, I.; Boopathy, A.V.; Khan, R.S.; Martinez, M.D.; Brown, M.E.; Murthy, N.; Davis, M.E. Delivery of Nox2-NADPH oxidase siRNA with polyketal nanoparticles for improving cardiac function following myocardial infarction. Biomaterials 2013, 34, 7790–7798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, F.L.; Lu, X.; Strutt, B.; Hill, D.J.; Feng, Q. Nox2 deficiency protects against streptozotocin-induced beta-cell destruction and development of diabetes in mice. Diabetes 2010, 59, 2603–2611. [Google Scholar] [CrossRef] [Green Version]
- Dao, V.T.-V.; Elbatreek, M.H.; Altenhöfer, S.; Casas, A.I.; Pachado, M.P.; Neullens, C.T.; Knaus, U.G.; Schmidt, H.H.H.W. Isoform-selective NADPH oxidase inhibitor panel for pharmacological target validation. Free Radic. Biol. Med. 2020, 148, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Dao, V.T.-V.; Casas, A.I.; Maghzal, G.J.; Seredenina, T.; Kaludercic, N.; Robledinos-Anton, N.; Di Lisa, F.; Stocker, R.; Ghezzi, P.; Jaquet, V.; et al. Pharmacology and Clinical Drug Candidates in Redox Medicine. Antioxid. Redox Signal. 2015, 23, 1113–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griendling, K.K.; Ushio-Fukai, M. Redox control of vascular smooth muscle proliferation. J. Lab. Clin. Med. 1998, 132, 9–15. [Google Scholar] [CrossRef]
- Schröder, K.; Helmcke, I.; Palfi, K.; Krause, K.-H.; Busse, R.; Brandes, R.P. Nox1 Mediates Basic Fibroblast Growth Factor-Induced Migration of Vascular Smooth Muscle Cells. Arter. Thromb. Vasc. Biol. 2007, 27, 1736–1743. [Google Scholar] [CrossRef] [Green Version]
- Mofarrahi, M.; Brandes, R.P.; Görlach, A.; Hanze, J.; Terada, L.S.; Quinn, M.T.; Mayaki, D.; Petrof, B.; Hussain, S.N. Regulation of Proliferation of Skeletal Muscle Precursor Cells By NADPH Oxidase. Antioxid. Redox Signal. 2008, 10, 559–574. [Google Scholar] [CrossRef]
- Schröder, K.; Kohnen, A.; Aicher, A.; Liehn, E.A.; Büchse, T.; Stein, S.; Weber, C.; Dimmeler, S.; Brandes, R.P. NADPH Oxidase Nox2 Is Required for Hypoxia-Induced Mobilization of Endothelial Progenitor Cells. Circ. Res. 2009, 105, 537–544. [Google Scholar] [CrossRef]
- Donkó, Á.; Morand, S.; Korzeniowska, A.; Boudreau, H.E.; Zana, M.; Hunyady, L.; Geiszt, M.; Leto, T.L. Hypothyroidism-associated missense mutation impairs NADPH oxidase activity and intracellular trafficking of Duox2. Free Radic. Biol. Med. 2014, 73, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Rokutan, K.; Kawahara, T.; Kuwano, Y.; Tominaga, K.; Sekiyama, A.; Teshima-Kondo, S. NADPH Oxidases in the Gastrointestinal Tract: A Potential Role of Nox1 in Innate Immune Response and Carcinogenesis. Antioxid. Redox Signal. 2006, 8, 1573–1582. [Google Scholar] [CrossRef]
- Segal, B.H.; Grimm, M.J.; Khan, A.N.H.; Han, W.; Blackwell, T.S. Regulation of innate immunity by NADPH oxidase. Free Radic. Biol. Med. 2012, 53, 72–80. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, S.; Brault, J.; Stasia, M.-J.; Knaus, U.G. Genetic disorders coupled to ROS deficiency. Redox Biol. 2015, 6, 135–156. [Google Scholar] [CrossRef] [Green Version]
- Matute, J.D.; Arias, A.A.; Wright, N.A.M.; Wrobel, I.; Waterhouse, C.C.M.; Li, X.J.; Marchal, C.C.; Stull, N.D.; Lewis, D.B.; Steele, M.; et al. A new genetic subgroup of chronic granulomatous disease with autosomal recessive mutations in p40phox and selective defects in neutrophil NADPH oxidase activity. Blood 2009, 114, 3309–3315. [Google Scholar] [CrossRef]
- Bagaitkar, J.; Matute, J.D.; Austin, A.; Arias, A.A.; Dinauer, M.C. Activation of neutrophil respiratory burst by fungal particles requires phosphatidylinositol 3-phosphate binding to p40phox in humans but not in mice. Blood 2012, 120, 3385–3387. [Google Scholar] [CrossRef] [Green Version]
- Dinauer, M.C. Disorders of Neutrophil Function: An Overview. Recent Results Cancer Res. 2014, 1124, 501–515. [Google Scholar] [CrossRef]
- Casas, A.I.; Dao, V.T.-V.; Daiber, A.; Maghzal, G.J.; Di Lisa, F.; Kaludercic, N.; Leach, S.; Cuadrado, A.; Jaquet, V.; Seredenina, T.; et al. Reactive Oxygen-Related Diseases: Therapeutic Targets and Emerging Clinical Indications. Antioxid. Redox Signal. 2015, 23, 1171–1185. [Google Scholar] [CrossRef] [Green Version]
- Crotzer, V.L.; Matute, J.D.; Arias, A.A.; Zhao, H.; Quilliam, L.A.; Dinauer, M.C.; Blum, J.S. Cutting edge: NADPH oxidase modulates MHC class II antigen presentation by B cells. J. Immunol. 2012, 189, 3800–3804. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.H.; Devadas, S.; Kwon, J.; Pinto, L.A.; Williams, M.S. T cells express a phagocyte-type NADPH oxidase that is activated after T cell receptor stimulation. Nat. Immunol. 2004, 5, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Padgett, L.E.; Tse, H.M. NADPH Oxidase-Derived Superoxide Provides a Third Signal for CD4 T Cell Effector Responses. J. Immunol. 2016, 197, 1733–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarosz, E.L.; Chang, C.-H. The Role of Reactive Oxygen Species in Regulating T Cell-mediated Immunity and Disease. Immune Netw. 2018, 18, e14. [Google Scholar] [CrossRef]
- Yi, Y.; Hahm, S.H.; Lee, K.H. Retroviral Gene Therapy: Safety Issues and Possible Solutions. Curr. Gene Ther. 2005, 5, 25–35. [Google Scholar] [CrossRef]
- Ranzani, M.; Annunziato, S.; Adams, D.J.; Montini, E. Cancer gene discovery: Exploiting insertional mutagenesis. Mol. Cancer Res. 2013, 11, 1141–1158. [Google Scholar] [CrossRef] [Green Version]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Brüne, B.; Dehne, N.; Grossmann, N.; Jung, M.; Namgaladze, D.; Schmid, T.; Von Knethen, A.; Weigert, A. Redox Control of Inflammation in Macrophages. Antioxid. Redox Signal. 2013, 19, 595–637. [Google Scholar] [CrossRef] [Green Version]
- Elbatreek, M.H.; Mucke, H.; Schmidt, H.H.H.W. NOX Inhibitors: From Bench to Naxibs to Bedside. Handb. Exp. Pharmacol. 2020, 1–24. [Google Scholar] [CrossRef]
- Mitchell, J.P.; Carmody, R.J. Nf-κb and the transcriptional control of inflammation. Int. Rev. Cell Mol. Biol. 2018, 335, 41–84. [Google Scholar]
- Taniguchi, K.; Karin, M. NF-κB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Adelaja, A.; Hoffmann, A. Signaling Crosstalk Mechanisms That May Fine-Tune Pathogen-Responsive NFκB. Front. Immunol. 2019, 10, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS Crosstalk in Inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef]
- Lepetsos, P.; Papavassiliou, K.A.; Papavassiliou, A.G. Redox and nf-κb signaling in osteoarthritis. Free Radic. Biol. Med. 2019, 132, 90–100. [Google Scholar] [CrossRef]
- Kaur, U.; Banerjee, P.; Bir, A.; Sinha, M.; Biswas, A.; Chakrabarti, S. Reactive oxygen species, redox signaling and neuroinflammation in alzheimer’s disease: The nf-κb connection. Curr. Top Med. Chem. 2015, 15, 446–457. [Google Scholar] [CrossRef]
- Loukili, N.; Rosenblatt-Velin, N.; Rolli, J.; Levrand, S.; Feihl, F.; Waeber, B.; Pacher, P.; Liaudet, L. Oxidants positively or negatively regulate nuclear factor kappab in a context-dependent manner. J. Biol. Chem. 2010, 285, 15746–15752. [Google Scholar] [CrossRef] [Green Version]
- Frazão, J.B.; Thain, A.; Zhu, Z.; Luengo, M.; Condino-Neto, A.; Newburger, P.E. Regulation ofCYBBGene Expression in Human Phagocytes by a Distant Upstream NF-κB Binding Site. J. Cell. Biochem. 2015, 116, 2008–2017. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide | Sequence |
|---|---|
| com | 5-AAGAGAAACTCCTCTGCTGTGAA-3 |
| wt | 5-CGCACTGGAACCCCTGAGAAAGG-3 |
| mut | 5-GTTCTAATTCCATCAGAAGCTTATCG-3 |
| Oligonucleotide | Sequence |
|---|---|
| NOX2wt | |
| NOX2wt_f | 5-GAGGCAGAACCAACACTTAACC-3 |
| NOX2wt_r | 5-TGAAGAGATGTGCAATTGTGTG-3 |
| NOX2ko | |
| NOX2ko_f | 5-TGTCATTCTGGTGTGGTTGG-3 |
| NOX2ko_r | 5-TCTGGATTCATCGACTGTGG-3 |
| Oligonucleotide. | Sequence |
|---|---|
| NOX2 | |
| qRT-PCR_5P2 | 5-CCAACTGGGATAACGAGTTCA-3 |
| qRT-PCR_3P2 | 5-GAGAGTTTCAGCCAAGGCTTC-3 |
| GAPDH | |
| qRT-PCR_5P | 5-TTCACCACCATGGAGAAGGC-3 |
| qRT-PCR_3P | 5-GGCATGGACTGTGGTCATGA-3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Göllner, M.; Ihrig-Biedert, I.; Petermann, V.; Saurin, S.; Oelze, M.; Kröller-Schön, S.; Vujacic-Mirski, K.; Kuntic, M.; Pautz, A.; Daiber, A.; et al. NOX2ko Mice Show Largely Increased Expression of a Mutated NOX2 mRNA Encoding an Inactive NOX2 Protein. Antioxidants 2020, 9, 1043. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9111043
Göllner M, Ihrig-Biedert I, Petermann V, Saurin S, Oelze M, Kröller-Schön S, Vujacic-Mirski K, Kuntic M, Pautz A, Daiber A, et al. NOX2ko Mice Show Largely Increased Expression of a Mutated NOX2 mRNA Encoding an Inactive NOX2 Protein. Antioxidants. 2020; 9(11):1043. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9111043
Chicago/Turabian StyleGöllner, Monika, Irmgard Ihrig-Biedert, Victoria Petermann, Sabrina Saurin, Matthias Oelze, Swenja Kröller-Schön, Ksenija Vujacic-Mirski, Marin Kuntic, Andrea Pautz, Andreas Daiber, and et al. 2020. "NOX2ko Mice Show Largely Increased Expression of a Mutated NOX2 mRNA Encoding an Inactive NOX2 Protein" Antioxidants 9, no. 11: 1043. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9111043