Knockout Mouse Models for Peroxiredoxins


Department of Biochemistry, College of Medicine, Gachon University, Incheon 21999, Korea
Antioxidants 2020, 9(2), 182; https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9020182
Submission received: 31 January 2020
/
Revised: 16 February 2020
/
Accepted: 20 February 2020
/
Published: 22 February 2020
(This article belongs to the Special Issue Peroxiredoxin)
Abstract
:Peroxiredoxins (PRDXs) are members of a highly conserved peroxidase family and maintain intracellular reactive oxygen species (ROS) homeostasis. The family members are expressed in most organisms and involved in various biological processes, such as cellular protection against ROS, inflammation, carcinogenesis, atherosclerosis, heart diseases, and metabolism. In mammals, six PRDX members have been identified and are subdivided into three subfamilies: typical 2-Cys (PRDX1, PRDX2, PRDX3, and PRDX4), atypical 2-Cys (PRDX5), and 1-Cys (PRDX6) subfamilies. Knockout mouse models of PRDXs have been developed to investigate their in vivo roles. This review presents an overview of the knockout mouse models of PRDXs with emphases on the biological and physiological changes of these model mice.
1. Introduction
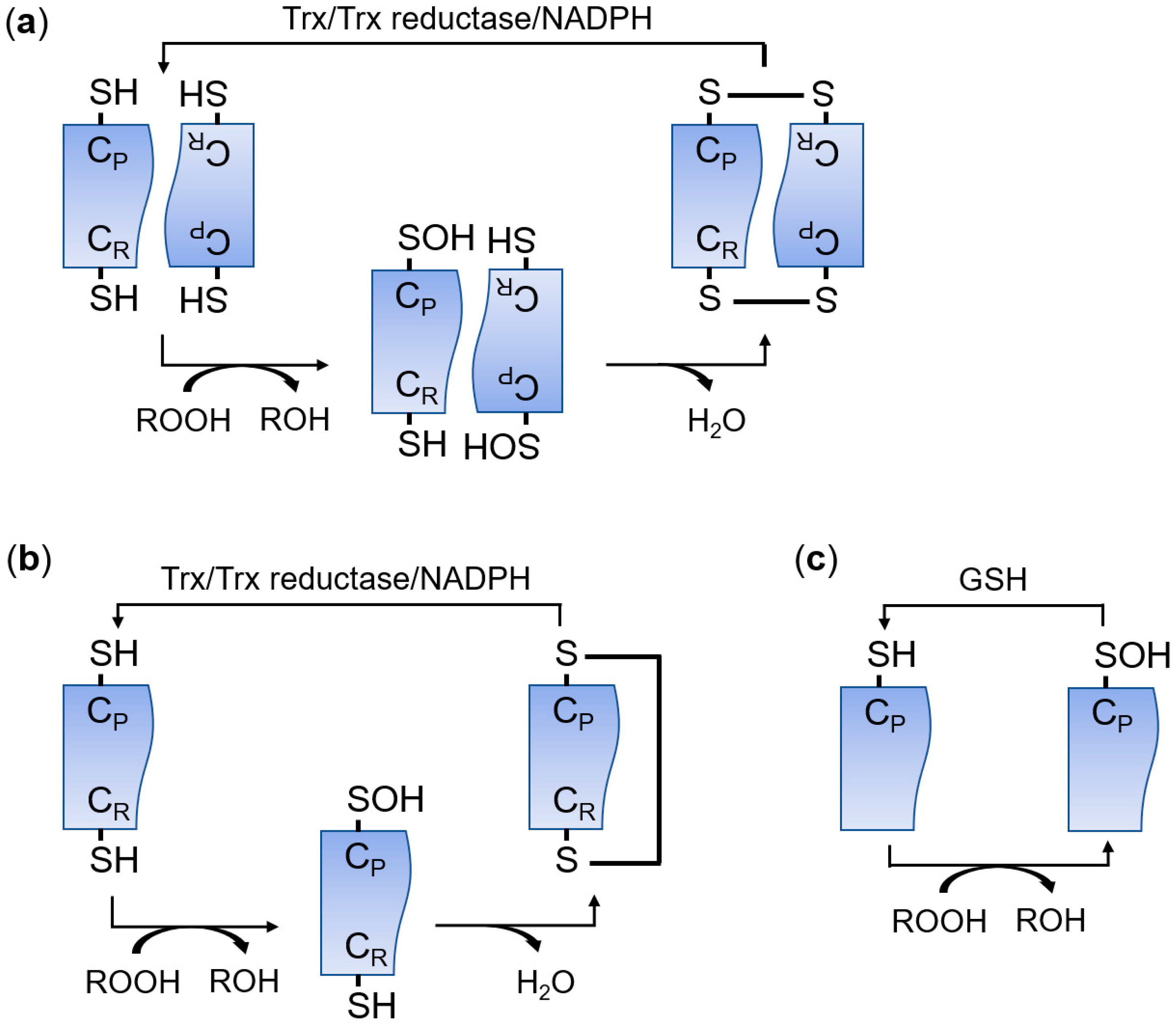
The peroxiredoxin (PRDX) family has peroxidase activity to remove peroxides, including hydrogen peroxide (H2O2), organic hydroperoxides, and peroxynitrite [1,2]. PRDXs are widely distributed in almost all organisms and there are more than 3500 members in this protein family [3]. PRDXs are classified into three subfamilies (typical 2-Cys, atypical 2-Cys, and 1-Cys) based on the number and location of the active cysteine residues and the type of disulfide bonds produced during the catalytic reaction (Figure 1) [4,5,6,7,8]. Typical 2-Cys PRDXs function as a homodimer [7]. Peroxides oxidize the conserved peroxidatic cysteine (CP) in typical 2-Cys PRDXs, and then the oxidized cysteine sulfenic acid residue in a subunit forms an intermolecular disulfide bond with the resolving cysteine (CR) in the other subunit [7]. Finally, the intermolecular disulfide bond is reduced by the thioredoxin (Trx)/Trx reductase/NADPH system [7]. Atypical 2-Cys PRDXs remove peroxides using the same mechanism as 2-Cys PRDXs except that atypical 2-Cys PRDXs form an intramolecular disulfide bond between CP and CR within a PRDX molecule [9]. 1-Cys PRDXs have only a CP residue and the oxidized CP is reduced by glutathione (GSH) instead of Trx [10]. According to the PeroxiRedoxin classification indEX (PREX) database that classifies PRDXs based on structural information around active sites, PRDXs are also divided into six subfamilies: AhpC-Prx1, BCP-PrxQ, Tpx, Prx5, Prx6, and AhpE [11,12]. In mammals, there are six PRDX members (PRDX1–PRDX6) [5]. PRDX1–PRDX4 are members of the typical 2-Cys or AhpC-Prx1 subfamily [5,12]. PRDX5 is a member of the atypical 2-Cys or Prx5 subfamily, and PRDX6 is classified into the 1-Cys or Prx6 subfamily [5,12]. Mammalian PRDXs are widely distributed in cells and perform various biological functions. PRDX1 is present in the nucleus and cytosol; PRDX2 and PRDX 6 are in the cytosol; PRDX3 is in the mitochondria; PRDX4 is in the endoplasmic reticulum (ER) and the cytosol; and PRDX5 is in the cytosol, peroxisomes, and mitochondria [8,13].
As genetically modified mouse (GEM) models, knockout mice are useful to investigate the roles of a gene. In the case of mouse Prdx genes, several knockout mouse strains targeting each Prdx gene have been generated by independent research groups [14]. These knockout mice provide useful information that is hard to obtain from other experiments. The present review summarizes the properties of Prdx-knockout mouse models and focuses on the biological and physiological changes of these model mouse strains.
2. PRDX1
2.1. Genetics and Knockout Mouse Strains
PRDX1 is a ubiquitously expressed nuclear and cytosolic peroxidase protein and is a member of the typical 2-Cys PRDX subfamily [7,13]. PRDX1 is involved in tumor suppression [15,16,17,18], inflammation [13,19,20,21,22,23,24,25,26], apoptosis [19,27,28], atherosclerosis [13,21], and molecular chaperoning [29,30]. According to the mouse Ensembl database, the Prdx1 gene is located on mouse chromosome 4 and there are five alternative forms of Prdx1 protein-coding transcripts [31]. Prdx1-knockout mouse strains have been generated by the homologous recombination [15,32] or gene trap [17,33] approaches. No Prdx1-knockout mouse strain with conditional potential has been reported, although the International Mouse Phenotyping Consortium (IMPC) has embryonic stem (ES) cell clones that possess the reporter and conditional allele [34].
2.2. Cancer
Neumann et al. have generated the first Prdx1-knockout mice (Prdx1−/−) [15]. The knockout mouse strain targets exon 3 of the Prdx1 gene by the insertion of a transposon containing a PGK-neomycin-resistant (neoR) cassette, which disrupts all protein-coding transcripts [15]. The knockout mice are viable and fertile but show severe hemolytic anemia and several malignant cancers, including lymphomas, sarcomas, and carcinomas, which cause a shortened life span [15]. Using the same knockout mice, Neumann’s group indicated that PRDX1 is involved in Pten-mediated tumor suppression in Ras-induced breast cancer [16] and inhibition of fibroblast transition into cancer-associated fibroblasts (CAFs) [18]. The tumor suppressor function of PRDX1 was also demonstrated in another study using a different Prdx1-knockout mouse strain, which was generated using the Lexicon gene trap ES cell clone, which has a gene trap vector in intron 3 of the Prdx1 gene [17]. Elevated nuclear ROS on primary tissues isolated from the Prdx1−/− leads to increased DNA damage and tumor susceptibility [17].
2.3. Erythrocytes
Severe hemolytic anemia with defects in blood parameters, which is observed in Neumann’s Prdx1-knockout mice, is not observed in other Prdx1-knockout mouse models [32,33], although Prdx1 deficiency aggravates hemolytic anemia symptoms in Prdx2-knockout mice [32]. These knockout mouse strains have been generated by replacing whole exons of the Prdx1 gene with a neoR cassette [32] or using the gene trap clone [33].
2.4. Inflammation
In different inflammation disease models, Prdx1−/− show conflicting results. PRDX1 enhances cerebral ischemia–reperfusion (I/R) injuries by activation of inflammation and apoptosis [19], and it initiates inflammation in the ozone-exposed lung [20]. Prdx1 deficiency, however, aggravates pulmonary inflammation and fibrosis in the bleomycin-treated model [26]. Atherosclerosis and chronic inflammation model mice (Prdx1−/−;ApoE−/−) show increased formation of atherosclerotic plaque compared with Prdx1+/+;ApoE−/− mice [13,21]. Prdx1-deficient macrophages present impaired lipophagic flux and cholesterol homeostasis [13]. Lipopolysaccharide (LPS)-induced lung injury, lethal shock, and neuroinflammation are increased in Prdx1−/− [22,23,24]. PRDX1 is a negative regulator of Th2-type allergic asthma that is induced by ovalbumin [25]. Inflammatory stimuli produce the intramolecular disulfide bond in HMGB1, which is mediated by PRDX1 or PRDX2 [35]. The formation of the disulfide bond is sufficient for HMGB1 secretion and secreted HMGB1 signals danger to surrounding cells. HMGB1 secretion induced by LPS is attenuated in macrophages isolated from Prdx1- or Prdx2-knockout mice [35].
2.5. Others
PRDX1 interacts with Gde2 and reduces the formation of an intramolecular disulfide bond between the N- and C-terminal regions of Gde2, which induces neuronal differentiation [36]. Prdx1 deficiency attenuates cisplatin-induced nephrotoxicity [37]. Studies using Prdx1-knockout mouse models also show that PRDX1 is involved in maintenance of stemness of mouse embryonic stem cells by suppression of ROS/JNK-induced neurogenesis [38], modulation of cellular senescence in mouse embryonic fibroblasts (MEFs) [39], host defenses against Mycobacterium tuberculosis and Staphylococcus aureus [40,41], and maintenance of progesterone production in the corpus luteum through regulating the unfolded protein response [42].
3. PRDX2
3.1. Genetics and Knockout Mouse Strains
PRDX2 is a cytosolic typical 2-Cys PRDX and has a similar structure to that of PRDX1 [7]. Mouse PRDX1 and PRDX2 proteins share 89% sequence similarity and 74% sequence identity and perform overlapping and distinct biological functions [31]. The Prdx2 gene is on mouse chromosome 8 and there are five alternative forms of Prdx2 protein-coding transcripts [31]. The first Prdx2-knockout mice (Prdx2−/−) have been generated by replacing the genomic DNA encoding exons 1-5 with a neoR cassette. They are viable and fertile [43] and most studies have used the same Prdx2-knockout mouse model. ES cell clones possessing the reporter and conditional allele of the Prdx2 gene have been produced [34], but a Prdx2-knockout mouse strain with conditional potential has not been reported.
3.2. Erythrocytes
Typical phenotypes of Prdx2−/− are hemolytic anemia, splenomegaly, Heinz body formation, and morphologically abnormal red blood cells [43]. PRDX2 is expressed in all cell types and is the third most abundant protein in erythrocytes [44]. Three PRDX isoforms (PRDX1, PRDX2, and PRDX6) are expressed in mature erythrocytes and PRDX2 is the most abundant protein among them [45]. The role of PRDX2 in protecting erythrocytes against oxidative stress has been verified by several studies using the same Prdx2-knockout mouse model [46,47,48,49,50,51,52,53,54,55]. PRDX2 has roles to protect erythrocytes from ROS-mediated DNA damage during erythropoiesis [48] and to protect hemoglobin from oxidative stress [49,50]. The decameric structure of PRDX2 binds to hemoglobin to stabilize and protect the protein [49]. Studies using Prdx2−/− show that PRDX2 is involved in the homeostasis of iron and membrane proteins of erythrocytes, as well as cellular senescence of erythrocytes and skin cells [51,53,56]. Erythrocytes lose PRDX2 protein gradually during the life span of erythrocytes [54]. PRDX2 is hyperoxidized by H2O2 and the hyperoxidized PRDX2 is degraded by the 20S proteasome [54].
3.3. Blood Vessels
PRDX2 is also involved in the homeostasis of blood vessels [57,58,59]. The redox-sensitive transcription factor Nrf2 activates PRDX2 expression to protect vascular smooth muscle cells from oxidative vascular injury [57]. In vascular endothelial cells, VEGFR2 no longer responds to VEGF stimulation by the oxidative-stress-induced formation of a cysteine disulfide bond in the C-terminal region of VEGFR2 [58]. PRDX2, not PRDX1, inhibits the formation of the intramolecular disulfide bond in VEGFR2 [58]. Furthermore, tumor angiogenesis is suppressed in Prdx2−/− [58]. The antioxidant activity of PRDX2 needs negative regulation of collagen-stimulated platelet activation and platelet-dependent thrombosis [60]. Among 2-Cys Prdxs, Prdx2 deficiency exacerbates the neointimal hyperplasia induced by the balloon injury of the carotid arteries [59].
3.4. Immune Responses
ROS are harmful byproducts but are also essential for immune responses [61] and their scavenger, PRDX2, inhibits immune cell responsiveness [62,63]. Increased exposure to ROS by Prdx2 deficiency activates the proliferation of T lymphocytes and the differentiation of dendritic cells [62,64]. Like Prdx1−/−, Prdx2−/− are sensitive to LPS-induced inflammatory responses, including lethal shock [65]. LPS activates inflammatory responses which are mediated by NADPH-oxidase-derived ROS generation in Prdx2-deficient macrophages [65]. Prdx2 deficiency increases immune cell accumulation in atherosclerotic lesions, which exacerbates atherosclerosis in ApoE−/− mice [66]. Hypoxia-induced oxidative stress in the lung of Prdx2−/− causes an amplified inflammatory response, vascular dysfunction, and autophagy activation, which lead to the development of pulmonary arterial hypertension [67]. Prdx2 deficiency ameliorates dextran sulfate sodium (DSS)-induced colitis by enhancing the development of Foxp3+ regulatory T cells [68].
3.5. Cancer
In the Apc+/Min colorectal cancer model, the depletion of Prdx2 inactivates the formation of intestinal adenomatous polyposis through Axin/β-catenin signaling [69,70]. Increased intracellular H2O2 level by the Apc mutation leads to the direct binding of PRDX2 to a poly(ADP-ribose) polymerase (PARP) tankyrase. This binding protects the tankyrase from its oxidative inactivation, and thus induces PARP-dependent Axin degradation [69,70].
3.6. Bone
Prdx2−/− have higher levels of bone mass than those of wild-type mice because PRDX2 is a negative regulator of BMP2-induced osteoblast differentiation [45]. PRDX2 also functions as a negative regulator of LPS-induced osteoclastogenesis and bone loss, which are induced by ROS-mediated JNK and STAT3 activation [71].
3.7. Others
PRDX2 is important for homeostasis of other tissues. PRDX2 protects hippocampal neurons from age-dependent mitochondrial decay [72] and maintains the stemness of mouse embryonic stem cells [38]. Oxidation of protein tyrosine phosphatases by ROS in Prdx2−/− fed a high-fat diet causes reduced body weight and increased glucose clearance [73]. PRDX2 controls corpus luteum regression that is induced by prostaglandin F2α-mediated ROS and protects against age-related ovarian failure [74,75].
4. Prdx3
4.1. Genetics and Knockout Mouse Strains
PRDX3 is a member of typical 2-Cys PRDXs and is mainly localized in mitochondria due to a mitochondrial targeting sequence at the N-terminal region. The Prdx3 gene is localized on mouse chromosome 19 and there is a Prdx2 protein-coding transcript [31]. Two knockout mouse lines have been analyzed to study the in vivo function of PRDX3. The first knockout mouse line was produced in 2007 using an ES cell clone generated by the gene trap approach [76]. In this ES clone, the VICTR20 gene trap vector [77] is inserted in intron 1 of the Prdx3 gene [76]. The second knockout mouse line was generated by the homologous recombination approach [78]. The genomic DNA region possessing exons 1-4 of the Prdx3 gene is replaced with the neoR cassette in the knockout mouse line [78]. ES cell clones possessing the reporter and conditional allele of the Prdx3 gene have been produced [34], but no Prdx3-knockout mouse strain with conditional potential has been reported.
4.2. Muscles
PRDX3 is important to protect mitochondria against oxidative stress. Prdx3 deficiency leads to reduced mitochondrial DNA content and ATP production and impaired mitochondrial fusion [79,80]. Mitochondrial homeostasis is necessary for the proper function of skeletal muscles. PRDX3 has roles in the prolonged contraction of skeletal muscles and physical strength [79,80].
4.3. Metabolism
PRDX3 is involved in metabolic homeostasis. Prdx3−/− show increased fat mass by adipocyte hypertrophy, impaired mitochondrial enzymes, and adipokine dysregulation, resulting in impaired glucose tolerance and insulin resistance [78].
4.4. Others
PRDX3 protects the lungs from LPS-induced damages, such as 8-hydroxy-2′-deoxyguanosine (8-OHdG) formation and protein carbonylation [76]. PRDX3 also protects macrophages and the liver against LPS-induced oxidative stress and pyrazole-induced oxidative damage, respectively [81,82]. Increased oxidative stress in Prdx3−/− shows placental defects, including focal necrosis and hyaline degeneration in trophoblast giant cells and vessel degeneration [83,84]. PRDX3 also has a protective role in UV-induced apoptosis of epidermal keratinocytes [85].
5. Prdx4
5.1. Genetics and Knockout Mouse Strains
PRDX4 is a member of typical 2-Cys PRDXs. Prdx4 is on the X chromosome and produces two forms of alternative transcripts [31]. Each of them uses a different exon 1 (exon 1A and exon 1B) [86]. All tissues, including the testis, express Prdx4 mRNAs transcribed from exon 1B, which encodes the cleavable N-terminal signal sequence, whereas the testis produces a testis-specific form of Prdx4 mRNAs transcribed from exon 1A [86]. PRDX4 is predominantly present in the ER and secreted to extracellular space [87,88]. A Prdx4-knockout mouse strain has been widely used to study in vivo functions of PRDX4 [89]. Originally, the knockout mouse line was generated with conditional potential by insertion of two loxP sequences flanking exon 1B of the Prdx4 gene [89]. However, most studies have used Prdx4-null mice, in which exon 1B is deleted. The testis-specific form of PRDX4 is not deleted in this knockout mouse strain [86].
5.2. Phenotypes
Prdx4-knockout male mice (Prdx4−/y) are fertile but show testicular atrophy [89]. Spermatogenic cells in Prdx4−/y are susceptible to cell death by oxidative stress [89]. In a DSS-induced colitis model, Prdx4−/y show loss of body weight and shortening of colon length, which may be caused by ER stress and oxidative damage in colonic epithelial cells [90]. Prdx4−/y show a higher incidence of hepatocellular carcinoma in the diethylnitrosamine injection model compared with that of wild-type mice [91]. Triple deletion of ER thiol oxidases, Ero1l and Ero1lb, and Prdx4 causes interfered procollagen maturation and thus forms defective connective tissues in the extracellular matrix [92]. Prdx4 and superoxide dismutase 1 (Sod1) double-knockout mice (Prdx4−/y;Sod−/−) show more severe liver phenotypes, such as aggravated liver steatosis and liver failure, at a relatively young age compared with those of wild-type, Prdx4−/y, and Sod−/− [93].
6. Prdx5
6.1. Genetics and Knockout Mouse Strains
PRDX5 is a unique member of the atypical 2-Cys subfamily in mammals and is ubiquitously expressed in tissues [5,12]. PRDX5 is present in the cytosol, peroxisomes, and mitochondria [8]. The Prdx5 gene is located on mouse chromosome 19 and there are four alternative forms of Prdx5 protein-coding transcripts [31]. A Prdx5-knockout mouse line has been generated by the homologous recombination approach [94]. Another Prdx5-knockout mouse line that is generated by the gene trap approach is commercially available [95]. In this knockout line, a gene trap vector is inserted in the 5′UTR region of the Prdx5 gene. ES cell clones possessing the reporter and conditional allele of the Prdx5 gene have been produced [34], but a Prdx5-knockout mouse strain with conditional potential has not been reported.
6.2. Metabolism
Prdx5 deficiency leads to increased susceptibility to high-fat-diet-induced obesity, and thus Prdx5-knockout mice fed a high-fat diet show several metabolic abnormalities, including increased body weight, adipocyte hypertrophy, fat accumulation in the liver, hepatic steatosis, and an increased triglyceride level in the serum [94,96].
7. Prdx6
7.1. Genetics and Knockout Mouse Strains
In mammals, PRDX6 is a unique member of the 1-Cys subfamily [5,12]. The Prdx6 gene is located on mouse chromosome 1 and produces two forms of alternatively spliced protein-coding transcripts [31]. PRDX6 is widely expressed in tissues and localized in the cytosol [8,97]. Two Prdx6-knockout mouse lines have been generated [97,98]. Exons 1 and 2 of the Prdx6 gene are replaced by a LacZ reporter and a neoR cassette in the first knockout mouse line [97], and a part of exon 3 is replaced by a neoR cassette in the second line [98]. Both knockout mouse lines are viable, fertile, and display no gross morphological defects [97,98]. IMPC produced a Prdx6-conditional knockout mouse line [34], although the mouse line has not been used for detailed phenotyping.
7.2. Tissue Protection
The protective roles of PRDX6 in the lung have been analyzed with Prdx6−/−. Administration of paraquat, an herbicide that produces damaging ROS within cells, causes tissue damage, decreased survival rate, and increased oxidation of lipids and proteins in the lungs of Prdx6−/− [97,99]. Exposure to 100% oxygen leads to similar defects in the lungs of Prdx6−/− [100]. Comparison between glutathione peroxidase 1 (Gpx1)-knockout mice and Prdx6−/− reveals that PRDX6 is the major enzyme for the reduction of phospholipid hydroperoxides in the lung [101]. In addition to the glutathione-dependent peroxidase activity, PRDX6 also has phospholipaseA2 (PLA2) and lysophospholipid:acyltransferase activities [102,103]. The deficiency of PLA2 activity alters phospholipid metabolism in the lungs of Prdx6−/− [102]. The lung and pulmonary microvascular endothelial cells (PMVECs) isolated from Prdx6−/− show increased sensitivity to peroxidative stress induced by exposure to 100% oxygen or tert-butyl hydroperoxide (t-BOOH) treatment [104,105]. These defects are partially rescued by the expression of mutant PRDX6 with either peroxidase activity alone or PLA2 activity alone [104,105]. However, coexpression of these mutant forms of PRDX6 rescues Prdx6-null PMVECs treated with t-BOOH as well as the expression of wild-type PRDX6 [104]. The glutathione-dependent peroxidase activity of PRDX6 can reduce both short-chain hydroperoxides such as H2O2 and phospholipid hydroperoxides [106]. The repair of peroxidized cell membranes of the lung or PMVECs is mostly dependent on the phospholipid hydroperoxidase activity rather than peroxidase activity toward H2O2 [106]. Protective effects of PRDX6 in the lung have been revealed using other lung injury models, including exposure to H2O2, LPS, or chronic cigarette smoke and cecal ligation and puncture (CLP)-induced acute lung injury [107,108,109,110]. Interestingly, the treatment of angiotensin II or phorbol ester increases the generation of superoxide and H2O2 in wild-type PMVECs but not in Prdx6-deficient PMVECs [111,112]. The authors explain that the PLA2 activity of PRDX6 is necessary for the activation of NADPH oxidase type 2 (NOX2), which produces superoxide [111,112]. PRDX6 also has protective roles in other tissues. PRDX6 protects the kidney from metabolic acidosis by contributing to the maintenance of anion exchanger 1 [113] and blood vessels in wounded skin [114].
7.3. Prion Disease
7.4. Inflammation and Metabolism
Oxidative stress contributes to the pathogenesis of various inflammatory and metabolic diseases. Hepatic I/R injury causes a significant increase of PRDX6 expression and PRDX6 transfer from the cytoplasm to the mitochondria [116]. Prdx6 deficiency in the I/R model increases the mitochondrial generation of H2O2 and mitochondrial dysfunction, thus leading to severe hepatocellular damage [116]. In Prdx6−/−, ethanol-induced lipid accumulation and peroxidation are observed in the liver [117]. An intensive study shows that Prdx6−/− develop insulin resistance, diabetic dyslipidemia, impaired insulin signaling, morphological changes in the pancreas and liver, and increased pro-inflammatory responses, suggesting that Prdx6 deficiency is a key mediator of hyperglycemia in type 2 diabetes [118]. A study suggests that PRDX6 is involved in the biosynthesis of fatty acid esters of hydroxy fatty acids that are lipid mediators with potent antidiabetic and anti-inflammatory activities [119]. In acute and chronic DSS-induced colitis models, however, Prdx6 deficiency attenuates the development of colitis [120]. The authors explain that Prdx6 deficiency is compensated by the upregulation of other PRDXs (PRDX3 and PRDX4) and antioxidant enzymes (Nrf2, Gss, and Gclm). A study tested the relationship between PRDX6 and atherosclerosis using Prdx6−/− with three different genetic backgrounds: atherosclerosis-resistant 129/SvJ (129), atherosclerosis-susceptible B6, and mixed B6;129 [121]. The effects of Prdx6 deficiency in atherosclerosis are minor and background dependent.
7.5. Aging
Prdx6−/− show age-related phenotypes [122,123,124]. Prdx6 deficiency decreases the fertility of male Prdx6−/− in an age-dependent manner [122]. PRDX6 protects spermatozoa against the oxidative stress that causes protein oxidation, lipid peroxidation, and DNA oxidation and fragmentation [122,123]. Lens epithelial cells (LECs) isolated from Prdx6−/− display elevated ROS expression and ER-stress-associated phenotypes [124,125]. Human LECs derived from aged men show a decreased level of PRDX6 and ER-stress-associated phenotypes [124]. LECs of Prdx6−/− are also more vulnerable to UV irradiation than those of wild-type mice [126].
7.6. Cancer
Prdx6 deficiency enhances susceptibility to tumorigenesis in the human-papillomavirus-8-induced skin cancer model [127]. The anti-tumorigenic effect of PRDX6 is achieved by the reduction of oxidative stress rather than altered proliferation, apoptosis, or the inflammatory response in keratinocytes [127].
8. Conclusions
PRDXs are typical peroxidases for the removal of cellular peroxides [1,2]. To investigate the biological roles of PRDXs, numerous approaches have been performed using the cell culture system. Although these experiments have provided valuable insights into PRDX biology, the approaches are not sufficient to reveal physiological functions in the human body. Knockout mouse models offer more reliable data to understand the in vivo functions of PRDXs (Table 1). More than one knockout mouse model of each PRDX has been generated and analyzed intensively. These knockout mouse models show that each PRDX functions essentially as a similar peroxidase and also performs specific functions depending on organs or intracellular organelles. In the past, the generation of knockout mouse models was time-consuming and labor-intensive work. However, recently developed gene-editing techniques using the CRISPR/Cas9 system have dramatically reduced these efforts [128]. Now, we can easily obtain GEM models that are more precisely modified than the previous complete knockout mouse models. Future studies using these new models, as well as complete knockout mouse models, will help us better understand the physiological roles of PRDXs and provide possible therapeutic targets for drugs against diseases, such as cancer and inflammatory and metabolic diseases.
Funding
This work was supported by the Gachon University research fund of 2019 (GCU-2019-0310).
Conflicts of Interest
The author declares no conflict of interest.
References
- Rhee, S.G. Overview on Peroxiredoxin. Mol. Cells 2016, 39, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Jang, H.H. The Role of Peroxiredoxin Family in Cancer Signaling. J. Cancer Prev. 2019, 24, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, K.J.; Knutson, S.T.; Soito, L.; Klomsiri, C.; Poole, L.B.; Fetrow, J.S. Analysis of the peroxiredoxin family: Using active-site structure and sequence information for global classification and residue analysis. Proteins 2011, 79, 947–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chae, H.Z.; Robison, K.; Poole, L.B.; Church, G.; Storz, G.; Rhee, S.G. Cloning and sequencing of thiol-specific antioxidant from mammalian brain: Alkyl hydroperoxide reductase and thiol-specific antioxidant define a large family of antioxidant enzymes. Proc. Natl. Acad. Sci. USA 1994, 91, 7017–7021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, S.G.; Kang, S.W.; Chang, T.S.; Jeong, W.; Kim, K. Peroxiredoxin, a novel family of peroxidases. IUBMB Life 2001, 52, 35–41. [Google Scholar] [CrossRef]
- Wood, Z.A.; Schroder, E.; Robin Harris, J.; Poole, L.B. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 2003, 28, 32–40. [Google Scholar] [CrossRef]
- Hall, A.; Karplus, P.A.; Poole, L.B. Typical 2-Cys peroxiredoxins–structures, mechanisms and functions. FEBS J. 2009, 276, 2469–2477. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.G.; Woo, H.A.; Kil, I.S.; Bae, S.H. Peroxiredoxin functions as a peroxidase and a regulator and sensor of local peroxides. J. Biol. Chem. 2012, 287, 4403–4410. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.S.; Kang, S.W.; Kim, K.; Baines, I.C.; Lee, T.H.; Rhee, S.G. Identification of a new type of mammalian peroxiredoxin that forms an intramolecular disulfide as a reaction intermediate. J. Biol. Chem. 2000, 275, 20346–20354. [Google Scholar] [CrossRef] [Green Version]
- Hall, A.; Nelson, K.; Poole, L.B.; Karplus, P.A. Structure-based insights into the catalytic power and conformational dexterity of peroxiredoxins. Antioxid. Redox Signal. 2011, 15, 795–815. [Google Scholar] [CrossRef] [Green Version]
- Soito, L.; Williamson, C.; Knutson, S.T.; Fetrow, J.S.; Poole, L.B.; Nelson, K.J. PREX: PeroxiRedoxin classification indEX, a database of subfamily assignments across the diverse peroxiredoxin family. Nucleic Acids Res. 2011, 39, D332–D337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- PeroxiRedoxin Classification IndEX. Available online: http://csb.wfu.edu/prex.test/ (accessed on 26 September 2019).
- Jeong, S.J.; Kim, S.; Park, J.G.; Jung, I.H.; Lee, M.N.; Jeon, S.; Kweon, H.Y.; Yu, D.Y.; Lee, S.H.; Jang, Y.; et al. Prdx1 (peroxiredoxin 1) deficiency reduces cholesterol efflux via impaired macrophage lipophagic flux. Autophagy 2018, 14, 120–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouse Genome Informatics. Available online: http://www.informatics.jax.org/ (accessed on 26 September 2019).
- Neumann, C.A.; Krause, D.S.; Carman, C.V.; Das, S.; Dubey, D.P.; Abraham, J.L.; Bronson, R.T.; Fujiwara, Y.; Orkin, S.H.; Van Etten, R.A. Essential role for the peroxiredoxin Prdx1 in erythrocyte antioxidant defence and tumour suppression. Nature 2003, 424, 561–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Schulte, J.; Knight, A.; Leslie, N.R.; Zagozdzon, A.; Bronson, R.; Manevich, Y.; Beeson, C.; Neumann, C.A. Prdx1 inhibits tumorigenesis via regulating PTEN/AKT activity. EMBO J. 2009, 28, 1505–1517. [Google Scholar] [CrossRef] [Green Version]
- Egler, R.A.; Fernandes, E.; Rothermund, K.; Sereika, S.; de Souza-Pinto, N.; Jaruga, P.; Dizdaroglu, M.; Prochownik, E.V. Regulation of reactive oxygen species, DNA damage, and c-Myc function by peroxiredoxin 1. Oncogene 2005, 24, 8038–8050. [Google Scholar] [CrossRef] [Green Version]
- Jezierska-Drutel, A.; Attaran, S.; Hopkins, B.L.; Skoko, J.J.; Rosenzweig, S.A.; Neumann, C.A. The peroxidase PRDX1 inhibits the activated phenotype in mammary fibroblasts through regulating c-Jun N-terminal kinases. BMC Cancer 2019, 19, 812. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, Y. PRDX1 enhances cerebral ischemia-reperfusion injury through activation of TLR4-regulated inflammation and apoptosis. Biochem. Biophys. Res. Commun. 2019, 519, 453–461. [Google Scholar] [CrossRef]
- Yanagisawa, R.; Warabi, E.; Inoue, K.; Yanagawa, T.; Koike, E.; Ichinose, T.; Takano, H.; Ishii, T. Peroxiredoxin I null mice exhibits reduced acute lung inflammation following ozone exposure. J. Biochem. 2012, 152, 595–601. [Google Scholar] [CrossRef]
- Kisucka, J.; Chauhan, A.K.; Patten, I.S.; Yesilaltay, A.; Neumann, C.; Van Etten, R.A.; Krieger, M.; Wagner, D.D. Peroxiredoxin1 prevents excessive endothelial activation and early atherosclerosis. Circ. Res. 2008, 103, 598–605. [Google Scholar] [CrossRef] [Green Version]
- Lv, W.P.; Li, M.X.; Wang, L. Peroxiredoxin 1 inhibits lipopolysaccharide-induced oxidative stress in lung tissue by regulating P38/JNK signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1876–1883. [Google Scholar]
- Sun, H.N.; Feng, L.; Wang, A.G.; Wang, J.Y.; Liu, L.; Jin, M.H.; Shen, G.N.; Jin, C.H.; Lee, D.S.; Kwon, T.H.; et al. Peroxiredoxin I deficiency increases LPSinduced lethal shock in mice. Mol. Med. Rep. 2018, 18, 2427–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.U.; Park, Y.H.; Min, J.S.; Sun, H.N.; Han, Y.H.; Hua, J.M.; Lee, T.H.; Lee, S.R.; Chang, K.T.; Kang, S.W.; et al. Peroxiredoxin I is a ROS/p38 MAPK-dependent inducible antioxidant that regulates NF-kappaB-mediated iNOS induction and microglial activation. J. Neuroimmunol. 2013, 259, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Takano, H.; Koike, E.; Warabi, E.; Yanagawa, T.; Yanagisawa, R.; Ishii, T. Peroxiredoxin I is a negative regulator of Th2-dominant allergic asthma. Int. Immunopharmacol. 2009, 9, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, N.; Ishii, Y.; Morishima, Y.; Yageta, Y.; Haraguchi, N.; Yamadori, T.; Masuko, H.; Sakamoto, T.; Yanagawa, T.; Warabi, E.; et al. Aggravation of bleomycin-induced pulmonary inflammation and fibrosis in mice lacking peroxiredoxin I. Am. J. Respir. Cell Mol. Biol. 2011, 45, 600–609. [Google Scholar] [CrossRef]
- Guo, W.; Liu, X.; Li, J.; Shen, Y.; Zhou, Z.; Wang, M.; Xie, Y.; Feng, X.; Wang, L.; Wu, X. Prdx1 alleviates cardiomyocyte apoptosis through ROS-activated MAPK pathway during myocardial ischemia/reperfusion injury. Int. J. Biol. Macromol. 2018, 112, 608–615. [Google Scholar] [CrossRef]
- Ito, T.; Kimura, S.; Seto, K.; Warabi, E.; Kawachi, Y.; Shoda, J.; Tabuchi, K.; Yamagata, K.; Hasegawa, S.; Bukawa, H.; et al. Peroxiredoxin I plays a protective role against UVA irradiation through reduction of oxidative stress. J. Dermatol. Sci. 2014, 74, 9–17. [Google Scholar] [CrossRef]
- Jang, H.H.; Kim, S.Y.; Park, S.K.; Jeon, H.S.; Lee, Y.M.; Jung, J.H.; Lee, S.Y.; Chae, H.B.; Jung, Y.J.; Lee, K.O.; et al. Phosphorylation and concomitant structural changes in human 2-Cys peroxiredoxin isotype I differentially regulate its peroxidase and molecular chaperone functions. FEBS Lett. 2006, 580, 351–355. [Google Scholar] [CrossRef] [Green Version]
- Nassour, H.; Wang, Z.; Saad, A.; Papaluca, A.; Brosseau, N.; Affar el, B.; Alaoui-Jamali, M.A.; Ramotar, D. Peroxiredoxin 1 interacts with and blocks the redox factor APE1 from activating interleukin-8 expression. Sci. Rep. 2016, 6, 29389. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, F.; Achuthan, P.; Akanni, W.; Allen, J.; Amode, M.R.; Armean, I.M.; Bennett, R.; Bhai, J.; Billis, K.; Boddu, S.; et al. Ensembl 2019. Nucleic Acids Res. 2019, 47, D745–D751. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.H.; Kwon, T.; Kim, S.U.; Ha, H.L.; Lee, T.H.; Kim, J.M.; Jo, E.K.; Kim, B.Y.; Yoon, D.Y.; Yu, D.Y. Peroxiredoxin I deficiency attenuates phagocytic capacity of macrophage in clearance of the red blood cells damaged by oxidative stress. BMB Rep. 2012, 45, 560–564. [Google Scholar] [CrossRef] [Green Version]
- Uwayama, J.; Hirayama, A.; Yanagawa, T.; Warabi, E.; Sugimoto, R.; Itoh, K.; Yamamoto, M.; Yoshida, H.; Koyama, A.; Ishii, T. Tissue Prx I in the protection against Fe-NTA and the reduction of nitroxyl radicals. Biochem. Biophys. Res. Commun. 2006, 339, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, M.E.; Flenniken, A.M.; Ji, X.; Teboul, L.; Wong, M.D.; White, J.K.; Meehan, T.F.; Weninger, W.J.; Westerberg, H.; Adissu, H.; et al. High-throughput discovery of novel developmental phenotypes. Nature 2016, 537, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.S.; Kim, H.S.; Lkhamsuren, K.; Kim, Y.H.; Han, M.G.; Shin, J.M.; Park, I.H.; Rhee, W.J.; Lee, S.K.; Rhee, S.G.; et al. Peroxiredoxin-mediated disulfide bond formation is required for nucleocytoplasmic translocation and secretion of HMGB1 in response to inflammatory stimuli. Redox Biol. 2019, 24, 101203. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Sabharwal, P.; Rao, M.; Sockanathan, S. The antioxidant enzyme Prdx1 controls neuronal differentiation by thiol-redox-dependent activation of GDE2. Cell 2009, 138, 1209–1221. [Google Scholar] [CrossRef] [Green Version]
- Okada, K.; Ma, D.; Warabi, E.; Morito, N.; Akiyama, K.; Murata, Y.; Yamagata, K.; Bukawa, H.; Shoda, J.; Ishii, T.; et al. Amelioration of cisplatin-induced nephrotoxicity in peroxiredoxin I-deficient mice. Cancer Chemother. Pharmacol. 2013, 71, 503–509. [Google Scholar] [CrossRef]
- Kim, S.U.; Park, Y.H.; Kim, J.M.; Sun, H.N.; Song, I.S.; Huang, S.M.; Lee, S.H.; Chae, J.I.; Hong, S.; Sik Choi, S.; et al. Dominant role of peroxiredoxin/JNK axis in stemness regulation during neurogenesis from embryonic stem cells. Stem Cells 2014, 32, 998–1011. [Google Scholar] [CrossRef]
- Park, Y.H.; Kim, H.S.; Lee, J.H.; Choi, S.A.; Kim, J.M.; Oh, G.T.; Kang, S.W.; Kim, S.U.; Yu, D.Y. Peroxiredoxin I participates in the protection of reactive oxygen species-mediated cellular senescence. BMB Rep. 2017, 50, 528–533. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, K.; Iwai, H.; Kato-Miyazawa, M.; Kirikae, F.; Zhao, J.; Yanagawa, T.; Ishii, T.; Miyoshi-Akiyama, T.; Funatogawa, K.; Kirikae, T. Peroxiredoxin 1 Contributes to Host Defenses against Mycobacterium tuberculosis. J. Immunol. 2016, 197, 3233–3244. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.N.; Liu, Y.; Wang, J.N.; Wang, C.; Liu, R.; Kong, L.Z.; Zhen, X.; Chandimali, N.; Cui, Y.D.; Kim, S.U.; et al. Protective Role of Peroxiredoxin I in Heat-Killed Staphylococcus Aureus-infected Mice. In Vivo 2019, 33, 749–755. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Lee, D.G.; Seong, J.B.; Lee, H.S.; Kwon, O.S.; Kang, B.S.; Park, J.W.; Lee, S.R.; Lee, D.S. Peroxiredoxin I maintains luteal function by regulating unfolded protein response. Reprod. Biol. Endocrinol. 2018, 16, 79. [Google Scholar] [CrossRef]
- Lee, T.H.; Kim, S.U.; Yu, S.L.; Kim, S.H.; Park, D.S.; Moon, H.B.; Dho, S.H.; Kwon, K.S.; Kwon, H.J.; Han, Y.H.; et al. Peroxiredoxin II is essential for sustaining life span of erythrocytes in mice. Blood 2003, 101, 5033–5038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, F.M.; Hampton, M.B.; Winterbourn, C.C. Peroxiredoxin 2 and peroxide metabolism in the erythrocyte. Antioxid. Redox Signal. 2008, 10, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Kim, D.Y.; Lee, D.S.; Kim, J.W.; Koh, J.T.; Kim, E.J.; Jang, W.G. Peroxiredoxin II negatively regulates BMP2-induced osteoblast differentiation and bone formation via PP2A Calpha-mediated Smad1/5/9 dephosphorylation. Exp. Mol. Med. 2019, 51, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.Y.; Jeong, D.K.; Kim, S.H.; Chung, K.J.; Cho, E.J.; Jin, C.H.; Yang, U.; Lee, S.R.; Lee, D.S.; Lee, T.H. Gene expression profiling related to the enhanced erythropoiesis in mouse bone marrow cells. J. Cell. Biochem. 2008, 104, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Kwon, J.; Choi, H.I.; Park, S.H.; Yang, U.; Park, H.R.; Ren, L.; Chung, K.J.; Kim, Y.U.; Park, B.J.; et al. In-depth analysis of cysteine oxidation by the RBC proteome: Advantage of peroxiredoxin II knockout mice. Proteomics 2012, 12, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.H.; Han, Y.H.; Hong, S.G.; Lee, D.J.; Ha, H.L.; Kang, S.W.; Li, W.; Yoon, D.Y.; Yu, D.Y. Reactive oxygen species mediated DNA damage is essential for abnormal erythropoiesis in peroxiredoxin II(-/-) mice. Biochem. Biophys. Res. Commun. 2012, 424, 189–195. [Google Scholar] [CrossRef]
- Han, Y.H.; Kim, S.U.; Kwon, T.H.; Lee, D.S.; Ha, H.L.; Park, D.S.; Woo, E.J.; Lee, S.H.; Kim, J.M.; Chae, H.B.; et al. Peroxiredoxin II is essential for preventing hemolytic anemia from oxidative stress through maintaining hemoglobin stability. Biochem. Biophys. Res. Commun. 2012, 426, 427–432. [Google Scholar] [CrossRef]
- Nagababu, E.; Mohanty, J.G.; Friedman, J.S.; Rifkind, J.M. Role of peroxiredoxin-2 in protecting RBCs from hydrogen peroxide-induced oxidative stress. Free Radic. Res. 2013, 47, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Matte, A.; Pantaleo, A.; Ferru, E.; Turrini, F.; Bertoldi, M.; Lupo, F.; Siciliano, A.; Ho Zoon, C.; De Franceschi, L. The novel role of peroxiredoxin-2 in red cell membrane protein homeostasis and senescence. Free Radic. Biol. Med. 2014, 76, 80–88. [Google Scholar] [CrossRef]
- Matte, A.; De Falco, L.; Iolascon, A.; Mohandas, N.; An, X.; Siciliano, A.; Leboeuf, C.; Janin, A.; Bruno, M.; Choi, S.Y.; et al. The Interplay Between Peroxiredoxin-2 and Nuclear Factor-Erythroid 2 Is Important in Limiting Oxidative Mediated Dysfunction in beta-Thalassemic Erythropoiesis. Antioxid. Redox Signal. 2015, 23, 1284–1297. [Google Scholar] [CrossRef] [Green Version]
- Matte, A.; De Falco, L.; Federti, E.; Cozzi, A.; Iolascon, A.; Levi, S.; Mohandas, N.; Zamo, A.; Bruno, M.; Lebouef, C.; et al. Peroxiredoxin-2: A Novel Regulator of Iron Homeostasis in Ineffective Erythropoiesis. Antioxid. Redox Signal. 2018, 28, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.S.; Yoon, H.J.; Kim, J.Y.; Woo, H.A.; Rhee, S.G. Circadian rhythm of hyperoxidized peroxiredoxin II is determined by hemoglobin autoxidation and the 20S proteasome in red blood cells. Proc. Natl. Acad. Sci. USA 2014, 111, 12043–12048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.M.; Ho, Y.S.; Yu, D.Y.; Kuypers, F.A.; Ravindranath, Y.; Goyette, G.W. The effects of disruption of genes for peroxiredoxin-2, glutathione peroxidase-1, and catalase on erythrocyte oxidative metabolism. Free Radic. Biol. Med. 2010, 48, 519–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.H.; Kim, H.S.; Kim, J.M.; Kim, S.K.; Yu, D.Y.; Moon, E.Y. Inhibitory role of peroxiredoxin II (Prx II) on cellular senescence. FEBS Lett. 2005, 579, 4897–4902. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Febbraio, M.; Reddy, S.P.; Yu, D.Y.; Yamamoto, M.; Silverstein, R.L. CD36 participates in a signaling pathway that regulates ROS formation in murine VSMCs. J. Clin. Investig. 2010, 120, 3996–4006. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.H.; Lee, D.J.; Lee, K.W.; Park, Y.S.; Lee, J.Y.; Lee, S.H.; Koh, Y.J.; Koh, G.Y.; Choi, C.; Yu, D.Y.; et al. Peroxiredoxin II is an essential antioxidant enzyme that prevents the oxidative inactivation of VEGF receptor-2 in vascular endothelial cells. Mol. Cell 2011, 44, 545–558. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.H.; Lee, D.J.; Kim, J.; Lee, J.Y.; Kim, H.W.; Kwon, K.; Taylor, W.R.; Jo, H.; Kang, S.W. Vascular injury involves the overoxidation of peroxiredoxin type II and is recovered by the peroxiredoxin activity mimetic that induces reendothelialization. Circulation 2013, 128, 834–844. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.Y.; Wang, S.B.; Min, J.H.; Chae, Y.H.; Baek, J.Y.; Yu, D.Y.; Chang, T.S. Peroxiredoxin II is an antioxidant enzyme that negatively regulates collagen-stimulated platelet function. J. Biol. Chem. 2015, 290, 11432–11442. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Bazhin, A.V.; Werner, J.; Karakhanova, S. Reactive oxygen species in the immune system. Int. Rev. Immunol. 2013, 32, 249–270. [Google Scholar] [CrossRef]
- Moon, E.Y.; Noh, Y.W.; Han, Y.H.; Kim, S.U.; Kim, J.M.; Yu, D.Y.; Lim, J.S. T lymphocytes and dendritic cells are activated by the deletion of peroxiredoxin II (Prx II) gene. Immunol. Lett. 2006, 102, 184–190. [Google Scholar] [CrossRef]
- Moon, E.Y.; Lee, J.H.; Oh, S.Y.; Ryu, S.K.; Kim, H.M.; Kwak, H.S.; Yoon, W.K. Reactive oxygen species augment B-cell-activating factor expression. Free Radic. Biol. Med. 2006, 40, 2103–2111. [Google Scholar] [CrossRef] [PubMed]
- Moon, E.Y.; Han, Y.H.; Lee, D.S.; Han, Y.M.; Yu, D.Y. Reactive oxygen species induced by the deletion of peroxiredoxin II (PrxII) increases the number of thymocytes resulting in the enlargement of PrxII-null thymus. Eur. J. Immunol. 2004, 34, 2119–2128. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Lee, D.S.; Song, C.H.; An, S.J.; Li, S.; Kim, J.M.; Kim, C.S.; Yoo, D.G.; Jeon, B.H.; Yang, H.Y.; et al. Roles of peroxiredoxin II in the regulation of proinflammatory responses to LPS and protection against endotoxin-induced lethal shock. J. Exp. Med. 2007, 204, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Park, J.G.; Yoo, J.Y.; Jeong, S.J.; Choi, J.H.; Lee, M.R.; Lee, M.N.; Hwa Lee, J.; Kim, H.C.; Jo, H.; Yu, D.Y.; et al. Peroxiredoxin 2 deficiency exacerbates atherosclerosis in apolipoprotein E-deficient mice. Circ. Res. 2011, 109, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Federti, E.; Matte, A.; Ghigo, A.; Andolfo, I.; James, C.; Siciliano, A.; Leboeuf, C.; Janin, A.; Manna, F.; Choi, S.Y.; et al. Peroxiredoxin-2 plays a pivotal role as multimodal cytoprotector in the early phase of pulmonary hypertension. Free Radic. Biol. Med. 2017, 112, 376–386. [Google Scholar] [CrossRef]
- Won, H.Y.; Jang, E.J.; Lee, K.; Oh, S.; Kim, H.K.; Woo, H.A.; Kang, S.W.; Yu, D.Y.; Rhee, S.G.; Hwang, E.S. Ablation of peroxiredoxin II attenuates experimental colitis by increasing FoxO1-induced Foxp3+ regulatory T cells. J. Immunol. 2013, 191, 4029–4037. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.H.; Lee, D.J.; Lee, S.; Lee, S.Y.; Jun, Y.; Kim, Y.; Kim, Y.; Lee, J.S.; Lee, D.K.; Lee, S.; et al. Interaction of tankyrase and peroxiredoxin II is indispensable for the survival of colorectal cancer cells. Nat. Commun. 2017, 8, 40. [Google Scholar] [CrossRef]
- Kang, D.H.; Lee, J.H.S.; Kang, S.W. Survival of APC-mutant colorectal cancer cells requires interaction between tankyrase and a thiol peroxidase, peroxiredoxin II. BMB Rep. 2017, 50, 391–392. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Noh, A.L.; Kang, J.H.; Sim, J.S.; Lee, D.S.; Yim, M. Peroxiredoxin II negatively regulates lipopolysaccharide-induced osteoclast formation and bone loss via JNK and STAT3. Antioxid. Redox Signal. 2015, 22, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.U.; Jin, M.H.; Kim, Y.S.; Lee, S.H.; Cho, Y.S.; Cho, K.J.; Lee, K.S.; Kim, Y.I.; Kim, G.W.; Kim, J.M.; et al. Peroxiredoxin II preserves cognitive function against age-linked hippocampal oxidative damage. Neurobiol. Aging 2011, 32, 1054–1068. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, S.J.; Chae, U.; Seong, J.; Lee, H.S.; Lee, S.R.; Lee, S.; Lee, D.S. Peroxiredoxin 2 mediates insulin sensitivity of skeletal muscles through regulation of protein tyrosine phosphatase oxidation. Int. J. Biochem. Cell Biol. 2018, 99, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, J.H.; Kim, T.S.; Lee, S.R.; Park, J.W.; Lee, S.; Kim, J.M.; Lee, D.S. Peroxiredoxin 2 regulates PGF2alpha-induced corpus luteum regression in mice by inhibiting ROS-dependent JNK activation. Free Radic. Biol. Med. 2017, 108, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, J.H.; Lee, D.G.; Kim, J.M.; Lee, D.S. Peroxiredoxin 2 deficiency accelerates age-related ovarian failure through the reactive oxygen species-mediated JNK pathway in mice. Free Radic. Biol. Med. 2018, 123, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Shoji, W.; Takano, H.; Nishimura, N.; Aoki, Y.; Takahashi, R.; Goto, S.; Kaifu, T.; Takai, T.; Obinata, M. Increased susceptibility of MER5 (peroxiredoxin III) knockout mice to LPS-induced oxidative stress. Biochem. Biophys. Res. Commun 2007, 355, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Zambrowicz, B.P.; Friedrich, G.A.; Buxton, E.C.; Lilleberg, S.L.; Person, C.; Sands, A.T. Disruption and sequence identification of 2,000 genes in mouse embryonic stem cells. Nature 1998, 392, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.Y.; Kim, Y.; Jeong, J.; Park, J.; Kim, I.; Huh, K.H.; Kim, Y.S.; Woo, H.A.; Rhee, S.G.; Lee, K.J.; et al. Peroxiredoxin 3 is a key molecule regulating adipocyte oxidative stress, mitochondrial biogenesis, and adipokine expression. Antioxid. Redox Signal. 2012, 16, 229–243. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.P.; Shin, Y.J.; Cho, S.C.; Lee, S.M.; Bahn, Y.J.; Kim, J.Y.; Kwon, E.S.; Jeong, D.Y.; Park, S.C.; Rhee, S.G.; et al. Peroxiredoxin 3 has a crucial role in the contractile function of skeletal muscle by regulating mitochondrial homeostasis. Free Radic. Biol. Med. 2014, 77, 298–306. [Google Scholar] [CrossRef]
- Zhang, Y.G.; Wang, L.; Kaifu, T.; Li, J.; Li, X.; Li, L. Featured Article: Accelerated decline of physical strength in peroxiredoxin-3 knockout mice. Exp. Biol. Med. 2016, 241, 1395–1400. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Kaifu, T.; Obinata, M.; Takai, T. Peroxiredoxin III-deficiency sensitizes macrophages to oxidative stress. J. Biochem. 2009, 145, 425–427. [Google Scholar] [CrossRef]
- Bae, S.H.; Sung, S.H.; Lee, H.E.; Kang, H.T.; Lee, S.K.; Oh, S.Y.; Woo, H.A.; Kil, I.S.; Rhee, S.G. Peroxiredoxin III and sulfiredoxin together protect mice from pyrazole-induced oxidative liver injury. Antioxid. Redox Signal. 2012, 17, 1351–1361. [Google Scholar] [CrossRef]
- Li, L.; Shoji, W.; Oshima, H.; Obinata, M.; Fukumoto, M.; Kanno, N. Crucial role of peroxiredoxin III in placental antioxidant defense of mice. FEBS Lett. 2008, 582, 2431–2434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Obinata, M.; Hori, K. Role of peroxiredoxin III in the pathogenesis of pre-eclampsia as evidenced in mice. Oxid. Med. Cell. Longev. 2010, 3, 71–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, J.Y.; Park, S.; Park, J.; Jang, J.Y.; Wang, S.B.; Kim, S.R.; Woo, H.A.; Lim, K.M.; Chang, T.S. Protective Role of Mitochondrial Peroxiredoxin III against UVB-Induced Apoptosis of Epidermal Keratinocytes. J. Investig. Dermatol. 2017, 137, 1333–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, S.H.; Kim, Y.J.; Oh, S.Y.; Fujii, J.; Zhang, Y.; Gladyshev, V.N.; Rhee, S.G. Identification and characterization of alternatively transcribed form of peroxiredoxin IV gene that is specifically expressed in spermatids of postpubertal mouse testis. J. Biol. Chem. 2011, 286, 39002–39012. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, A.; Okado, A.; Fujii, T.; Fujii, J.; Egashira, M.; Niikawa, N.; Taniguchi, N. Cloning of the peroxiredoxin gene family in rats and characterization of the fourth member. FEBS Lett. 1999, 443, 246–250. [Google Scholar] [CrossRef] [Green Version]
- Okado-Matsumoto, A.; Matsumoto, A.; Fujii, J.; Taniguchi, N. Peroxiredoxin IV is a secretable protein with heparin-binding properties under reduced conditions. J. Biochem. 2000, 127, 493–501. [Google Scholar] [CrossRef]
- Iuchi, Y.; Okada, F.; Tsunoda, S.; Kibe, N.; Shirasawa, N.; Ikawa, M.; Okabe, M.; Ikeda, Y.; Fujii, J. Peroxiredoxin 4 knockout results in elevated spermatogenic cell death via oxidative stress. Biochem. J. 2009, 419, 149–158. [Google Scholar] [CrossRef]
- Takagi, T.; Homma, T.; Fujii, J.; Shirasawa, N.; Yoriki, H.; Hotta, Y.; Higashimura, Y.; Mizushima, K.; Hirai, Y.; Katada, K.; et al. Elevated ER stress exacerbates dextran sulfate sodium-induced colitis in PRDX4-knockout mice. Free Radic. Biol. Med. 2019, 134, 153–164. [Google Scholar] [CrossRef]
- Guo, X.; Noguchi, H.; Ishii, N.; Homma, T.; Hamada, T.; Hiraki, T.; Zhang, J.; Matsuo, K.; Yokoyama, S.; Ishibashi, H.; et al. The Association of Peroxiredoxin 4 with the Initiation and Progression of Hepatocellular Carcinoma. Antioxid. Redox Signal. 2019, 30, 1271–1284. [Google Scholar] [CrossRef]
- Zito, E.; Hansen, H.G.; Yeo, G.S.; Fujii, J.; Ron, D. Endoplasmic reticulum thiol oxidase deficiency leads to ascorbic acid depletion and noncanonical scurvy in mice. Mol. Cell 2012, 48, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Homma, T.; Kurahashi, T.; Lee, J.; Nabeshima, A.; Yamada, S.; Fujii, J. Double Knockout of Peroxiredoxin 4 (Prdx4) and Superoxide Dismutase 1 (Sod1) in Mice Results in Severe Liver Failure. Oxid. Med. Cell. Longev. 2018, 2018, 2812904. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Park, S.J.; Kim, J.H.; Seong, J.B.; Kim, K.M.; Woo, H.A.; Lee, D.S. Peroxiredoxin 5 regulates adipogenesis-attenuating oxidative stress in obese mouse models induced by a high-fat diet. Free Radic. Biol. Med. 2018, 123, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.T.; Motenko, H.; Richardson, J.E.; Richards-Smith, B.; Smith, C.L. The International Mouse Strain Resource (IMSR): Cataloging worldwide mouse and ES cell line resources. Mamm. Genome 2015, 26, 448–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.H.; Seong, J.B.; Huh, J.W.; Bae, Y.C.; Lee, H.S.; Lee, D.S. Peroxiredoxin 5 ameliorates obesity-induced non-alcoholic fatty liver disease through the regulation of oxidative stress and AMP-activated protein kinase signaling. Redox Biol. 2020, 28, 101315. [Google Scholar] [CrossRef]
- Wang, X.; Phelan, S.A.; Forsman-Semb, K.; Taylor, E.F.; Petros, C.; Brown, A.; Lerner, C.P.; Paigen, B. Mice with targeted mutation of peroxiredoxin 6 develop normally but are susceptible to oxidative stress. J. Biol. Chem. 2003, 278, 25179–25190. [Google Scholar] [CrossRef] [Green Version]
- Mo, Y.; Feinstein, S.I.; Manevich, Y.; Zhang, Q.; Lu, L.; Ho, Y.S.; Fisher, A.B. 1-Cys peroxiredoxin knock-out mice express mRNA but not protein for a highly related intronless gene. FEBS Lett. 2003, 555, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Feinstein, S.I.; Manevich, Y.; Ho, Y.S.; Fisher, A.B. Peroxiredoxin 6 gene-targeted mice show increased lung injury with paraquat-induced oxidative stress. Antioxid. Redox Signal. 2006, 8, 229–237. [Google Scholar] [CrossRef]
- Wang, Y.; Feinstein, S.I.; Manevich, Y.; Ho, Y.S.; Fisher, A.B. Lung injury and mortality with hyperoxia are increased in peroxiredoxin 6 gene-targeted mice. Free Radic. Biol. Med. 2004, 37, 1736–1743. [Google Scholar] [CrossRef]
- Liu, G.; Feinstein, S.I.; Wang, Y.; Dodia, C.; Fisher, D.; Yu, K.; Ho, Y.S.; Fisher, A.B. Comparison of glutathione peroxidase 1 and peroxiredoxin 6 in protection against oxidative stress in the mouse lung. Free Radic. Biol. Med. 2010, 49, 1172–1181. [Google Scholar] [CrossRef] [Green Version]
- Fisher, A.B.; Dodia, C.; Feinstein, S.I.; Ho, Y.S. Altered lung phospholipid metabolism in mice with targeted deletion of lysosomal-type phospholipase A2. J. Lipid Res. 2005, 46, 1248–1256. [Google Scholar] [CrossRef] [Green Version]
- Fisher, A.B.; Dodia, C.; Sorokina, E.M.; Li, H.; Zhou, S.; Raabe, T.; Feinstein, S.I. A novel lysophosphatidylcholine acyl transferase activity is expressed by peroxiredoxin 6. J. Lipid Res. 2016, 57, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lien, Y.C.; Feinstein, S.I.; Dodia, C.; Fisher, A.B. The roles of peroxidase and phospholipase A2 activities of peroxiredoxin 6 in protecting pulmonary microvascular endothelial cells against peroxidative stress. Antioxid. Redox Signal. 2012, 16, 440–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Benipal, B.; Zhou, S.; Dodia, C.; Chatterjee, S.; Tao, J.Q.; Sorokina, E.M.; Raabe, T.; Feinstein, S.I.; Fisher, A.B. Critical role of peroxiredoxin 6 in the repair of peroxidized cell membranes following oxidative stress. Free Radic. Biol. Med. 2015, 87, 356–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, A.B.; Vasquez-Medina, J.P.; Dodia, C.; Sorokina, E.M.; Tao, J.Q.; Feinstein, S.I. Peroxiredoxin 6 phospholipid hydroperoxidase activity in the repair of peroxidized cell membranes. Redox Biol. 2018, 14, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Feinstein, S.I.; Fisher, A.B. Peroxiredoxin 6 as an antioxidant enzyme: Protection of lung alveolar epithelial type II cells from H2O2-induced oxidative stress. J. Cell. Biochem. 2008, 104, 1274–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundar, I.K.; Chung, S.; Hwang, J.W.; Arunachalam, G.; Cook, S.; Yao, H.; Mazur, W.; Kinnula, V.L.; Fisher, A.B.; Rahman, I. Peroxiredoxin 6 differentially regulates acute and chronic cigarette smoke-mediated lung inflammatory response and injury. Exp. Lung Res. 2010, 36, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; An, X.; Wang, X.; Hu, X.; Bi, J.; Tong, L.; Yang, D.; Song, Y.; Bai, C. Peroxiredoxin 6 knockout aggravates cecal ligation and puncture-induced acute lung injury. Int. Immunopharmacol. 2019, 68, 252–258. [Google Scholar] [CrossRef]
- Yang, D.; Jin, M.; Bai, C.; Zhou, J.; Shen, Y. Peroxiredoxin 6 suppresses Muc5ac overproduction in LPS-induced airway inflammation through H2O2-EGFR-MAPK signaling pathway. Respir. Physiol. Neurobiol. 2017, 236, 84–90. [Google Scholar] [CrossRef]
- Vazquez-Medina, J.P.; Dodia, C.; Weng, L.; Mesaros, C.; Blair, I.A.; Feinstein, S.I.; Chatterjee, S.; Fisher, A.B. The phospholipase A2 activity of peroxiredoxin 6 modulates NADPH oxidase 2 activation via lysophosphatidic acid receptor signaling in the pulmonary endothelium and alveolar macrophages. FASEB J. 2016, 30, 2885–2898. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Feinstein, S.I.; Dodia, C.; Sorokina, E.; Lien, Y.C.; Nguyen, S.; Debolt, K.; Speicher, D.; Fisher, A.B. Peroxiredoxin 6 phosphorylation and subsequent phospholipase A2 activity are required for agonist-mediated activation of NADPH oxidase in mouse pulmonary microvascular endothelium and alveolar macrophages. J. Biol. Chem. 2011, 286, 11696–11706. [Google Scholar] [CrossRef] [Green Version]
- Sorrell, S.L.; Golder, Z.J.; Johnstone, D.B.; Frankl, F.E.K. Renal peroxiredoxin 6 interacts with anion exchanger 1 and plays a novel role in pH homeostasis. Kidney Int. 2016, 89, 105–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumin, A.; Schafer, M.; Epp, N.; Bugnon, P.; Born-Berclaz, C.; Oxenius, A.; Klippel, A.; Bloch, W.; Werner, S. Peroxiredoxin 6 is required for blood vessel integrity in wounded skin. J. Cell. Biol. 2007, 179, 747–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asuni, A.A.; Guridi, M.; Sanchez, S.; Sadowski, M.J. Antioxidant peroxiredoxin 6 protein rescues toxicity due to oxidative stress and cellular hypoxia in vitro, and attenuates prion-related pathology in vivo. Neurochem. Int. 2015, 90, 152–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eismann, T.; Huber, N.; Shin, T.; Kuboki, S.; Galloway, E.; Wyder, M.; Edwards, M.J.; Greis, K.D.; Shertzer, H.G.; Fisher, A.B.; et al. Peroxiredoxin-6 protects against mitochondrial dysfunction and liver injury during ischemia-reperfusion in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G266–G274. [Google Scholar] [CrossRef] [Green Version]
- Roede, J.R.; Orlicky, D.J.; Fisher, A.B.; Petersen, D.R. Overexpression of peroxiredoxin 6 does not prevent ethanol-mediated oxidative stress and may play a role in hepatic lipid accumulation. J. Pharmacol. Exp. Ther. 2009, 330, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Pacifici, F.; Arriga, R.; Sorice, G.P.; Capuani, B.; Scioli, M.G.; Pastore, D.; Donadel, G.; Bellia, A.; Caratelli, S.; Coppola, A.; et al. Peroxiredoxin 6, a novel player in the pathogenesis of diabetes. Diabetes 2014, 63, 3210–3220. [Google Scholar] [CrossRef] [Green Version]
- Kuda, O.; Brezinova, M.; Silhavy, J.; Landa, V.; Zidek, V.; Dodia, C.; Kreuchwig, F.; Vrbacky, M.; Balas, L.; Durand, T.; et al. Nrf2-Mediated Antioxidant Defense and Peroxiredoxin 6 Are Linked to Biosynthesis of Palmitic Acid Ester of 9-Hydroxystearic Acid. Diabetes 2018, 67, 1190–1199. [Google Scholar] [CrossRef] [Green Version]
- Melhem, H.; Spalinger, M.R.; Cosin-Roger, J.; Atrott, K.; Lang, S.; Wojtal, K.A.; Vavricka, S.R.; Rogler, G.; Frey-Wagner, I. Prdx6 Deficiency Ameliorates DSS Colitis: Relevance of Compensatory Antioxidant Mechanisms. J. Crohns Colitis 2017, 11, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Phelan, S.A.; Petros, C.; Taylor, E.F.; Ledinski, G.; Jurgens, G.; Forsman-Semb, K.; Paigen, B. Peroxiredoxin 6 deficiency and atherosclerosis susceptibility in mice: Significance of genetic background for assessing atherosclerosis. Atherosclerosis 2004, 177, 61–70. [Google Scholar] [CrossRef]
- Ozkosem, B.; Feinstein, S.I.; Fisher, A.B.; O’Flaherty, C. Advancing age increases sperm chromatin damage and impairs fertility in peroxiredoxin 6 null mice. Redox Biol. 2015, 5, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Ozkosem, B.; Feinstein, S.I.; Fisher, A.B.; O’Flaherty, C. Absence of Peroxiredoxin 6 Amplifies the Effect of Oxidant Stress on Mobility and SCSA/CMA3 Defined Chromatin Quality and Impairs Fertilizing Ability of Mouse Spermatozoa. Biol. Reprod. 2016, 94, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatma, N.; Singh, P.; Chhunchha, B.; Kubo, E.; Shinohara, T.; Bhargavan, B.; Singh, D.P. Deficiency of Prdx6 in lens epithelial cells induces ER stress response-mediated impaired homeostasis and apoptosis. Am. J. Physiol. 2011, 301, C954–C967. [Google Scholar] [CrossRef] [PubMed]
- Fatma, N.; Kubo, E.; Sharma, P.; Beier, D.R.; Singh, D.P. Impaired homeostasis and phenotypic abnormalities in Prdx6-/-mice lens epithelial cells by reactive oxygen species: Increased expression and activation of TGFbeta. Cell Death Differ. 2005, 12, 734–750. [Google Scholar] [CrossRef] [PubMed]
- Kubo, E.; Hasanova, N.; Tanaka, Y.; Fatma, N.; Takamura, Y.; Singh, D.P.; Akagi, Y. Protein expression profiling of lens epithelial cells from Prdx6-depleted mice and their vulnerability to UV radiation exposure. Am. J. Physiol. 2010, 298, C342–C354. [Google Scholar] [CrossRef] [PubMed]
- Rolfs, F.; Huber, M.; Gruber, F.; Bohm, F.; Pfister, H.J.; Bochkov, V.N.; Tschachler, E.; Dummer, R.; Hohl, D.; Schafer, M.; et al. Dual role of the antioxidant enzyme peroxiredoxin 6 in skin carcinogenesis. Cancer Res. 2013, 73, 3460–3469. [Google Scholar] [CrossRef] [Green Version]
- Burgio, G. Redefining mouse transgenesis with CRISPR/Cas9 genome editing technology. Genome Biol. 2018, 19, 27. [Google Scholar] [CrossRef]
Figure 1.
Catalytic cycle of typical 2-Cys (a), atypical 2-Cys (b), and 1-Cys (c) peroxiredoxins (PRDXs). CP, peroxidatic cysteine; CR, resolving cysteine; GSH, glutathione; ROOH, peroxide; Trx, thioredoxin.
Figure 1.
Catalytic cycle of typical 2-Cys (a), atypical 2-Cys (b), and 1-Cys (c) peroxiredoxins (PRDXs). CP, peroxidatic cysteine; CR, resolving cysteine; GSH, glutathione; ROOH, peroxide; Trx, thioredoxin.


{kind=link}
Table 1.
Knockout mouse models of Prdxs.
| Gene | Models | Phenotypes | Challenges 1 | Affected Organs/Cells |
|---|---|---|---|---|
| Prdx1 | KO1 [15] (Homologous recombination, Insertion, exon 3) 2 | Tumorigenesis [15,16,18] | None | Various |
| Hemolytic anemia [15] | None | Red blood cells (RBCs) | ||
| Atherosclerosis [21] | Normal diet, ApoE−/− | Aorta | ||
| Neuronal defects [36] | None | Embryonic neurons | ||
| KO2 [32] (Homologous recombination, Replacement, exons 1–6) | Pro-inflammation [23] | Lipopolysaccharide (LPS) | Liver | |
| Defective RBC clearance [32] | None | Macrophages | ||
| Cellular senescence [39] | None | Mouse embryonic fibroblasts (MEFs) | ||
| Defective host defense [41] | Staphylococcus aureus | Liver, lung | ||
| KO3 [17] (Gene Trap, Insertion, Intron 3) | Tumorigenesis [17] | None | Brain, liver, spleen, MEFs | |
| KO4 [33] (Gene Trap, Insertion, Intron 3) | Anti-inflammation [20] | Ozone | Lung | |
| Pro-inflammation [26] | Bleomycin | Lung | ||
| Fibrosis [26] | Bleomycin | Lung | ||
| Asthma [25] | Cisplatin | Lung | ||
| Defective host defense [40] | Mycobacterium tuberculosis | Lung | ||
| Kidney defects [37] | Ovalbumin | Kidney | ||
| Pro-apoptosis [28] | UVA | MEFs | ||
| Uncertain | Anti-inflammation [19] | Ischemia–reperfusion (I/R) injury | Brain | |
| Pro-inflammation [24] | LPS | Microglia | ||
| Anti-apoptosis [19] | I/R injury | Brain | ||
| Pro-apoptosis [42] | Tunicamycin | Corpus luteum | ||
| Atherosclerosis [13] | Normal or high-fat diet, ApoE−/− | Aorta | ||
| Lung defects [22] | LPS | Lung, macrophages | ||
| Loss of stemness [38] | Differentiation medium | Embryonic stem (ES) cells | ||
| Reduced progesterone [42] | Tunicamycin | Corpus luteum | ||
| Defective HMGB1 secretion [35] | LPS | Macrophages | ||
| Prdx2 | KO1 [43] (Homologous recombination, Replacement, exons 1–5) | Hemolytic anemia [32,43,46,47,48,49,50,51,52,53,54,55] | None | RBCs, spleen, bone marrow (BM) |
| Defective iron homeostasis [53] | Carbonyl-iron, LPS | RBCs, liver, BM | ||
| Blood vessel defects [58,59] | None | Endothelial cells | ||
| Balloon injury | Carotid arteries | |||
| Platelet defects [60] | Collagen | Platelet | ||
| Pro-inflammation [62,63,64,65] | None | Spleen, BM, thymus | ||
| LPS | Macrophages | |||
| Anti-inflammation [68] | Dextran sulfate sodium (DSS) | Colon | ||
| Atherosclerosis [66] | Atherogenic cholate-containing diet, ApoE−/− | Aorta | ||
| Anti-tumorigenesis [69,70] | Apc+/Min | Intestine, colon | ||
| Metabolic defects [73] | High-fat diet | MEFs, serum, muscle | ||
| Ovary defects [74,75] | None, PGF2α, 4-vinylcyclohexene diepoxide | Ovary | ||
| Bone defects [45,71] | LPS | Bone | ||
| Neuronal defects [72] | None | Neurons | ||
| Pulmonary hypertension [67] | Hypoxia | Lung | ||
| Cellular senescence [56] | None | MEFs | ||
| Loss of stemness [38] | Differentiation medium | ES cells | ||
| Defective HMGB1 secretion [35] | LPS | Macrophages | ||
| Uncertain | Blood vessel defects [57] | FeCl3 | Carotid arteries | |
| Prdx3 | KO1 [78] (Homologous recombination, Replacement, exons 1–4) | Pro-apoptosis [85] | UVB | Keratinocytes |
| Muscle defects [79] | None | Muscle | ||
| Liver defects [82] | Pyrazole | Liver | ||
| Metabolic defects [78] | None | Fat, adipocytes | ||
| KO2 [76] (Gene Trap, Insertion, Intron 1) | Pro-apoptosis [80] | None | Brain | |
| Muscle defects [80] | None | Muscle | ||
| Lung defects [76] | LPS | Lung | ||
| Macrophage defects [81] | LPS | Macrophages | ||
| Placental defects [83,84] | None | Placenta | ||
| Prdx4 | KO1 [89] (Homologous recombination, Cre-loxP deletion, exon 1) | Defective spermatogenesis [89] | None | Testis |
| Tumorigenesis [91] | Diethylnitrosamine | Liver | ||
| Colon defects [90] | Dextran sulfate sodium | Colon | ||
| Liver defects [93] | Sod1−/− | Liver | ||
| Defective connective tissues [92] | Ero1l−/−, Ero1bl−/− | Connective tissues | ||
| Prdx5 | KO1 [94] (Homologous recombination, Replacement, not specified) | Metabolic defects [94,96] | High-fat diet | Fat, liver |
| Prdx6 | KO1 [97] (Homologous recombination, Replacement, exons 1 and 2) | Tissue defects [97] | Paraquat | Lung, kidney, liver, macrophages |
| Anti-inflammation [120] | DSS | Colon | ||
| Tumorigenesis [127] | Human papillomavirus 8 | Skin | ||
| Atherosclerosis (mild) [121] | Atherogenic high fat diet | Aorta, plasma | ||
| Metabolic defects [118] | None | Various | ||
| Lens defects [124,125] | None, hypoxia, CoCl2, tunicamycin, H2O2, | Lens epithelial cells (LECs) | ||
| Lens defects and pro-apoptosis [126] | UVB | LECs | ||
| Liver defects [117] | Ethanol | Liver | ||
| Prion disease [115] | ME7 | Brain | ||
| Vascular defects [114] | UV | Skin, blood vessels | ||
| KO2 [98] (Homologous recombination, Replacement, exon 3) | Lung defects [99,100,101,107,108,109,110] | Paraquat | Lung | |
| Hyperoxia | Lung | |||
| Hyperoxia, tert-butylhydroperoxide (t-BOOH), Paraquat | Lung, pulmonary microvascular endothelial cells (PMVECs) | |||
| H2O2 | Type II alveolar epithelial cells | |||
| Cigarette smoke | Lung | |||
| Cecal ligation and puncture | Lung | |||
| LPS | Lung | |||
| Lung defects (phospholipid metabolism) [102] | None | Lung | ||
| PMVEC defects [104] | t-BOOH | PMVECs | ||
| Lung and PMVEC defects [105,106] | t-BOOH, hyperoxia | PMVECs, lung | ||
| Attenuated production of superoxide and H2O2 [111,112] | Angiotensin II, Phorbol ester | Lung, PMVECs, alveolar macrophages | ||
| Defective spermatogenesis [122,123] | Aging, t-BOOH | Sperm | ||
| Liver defects [116] | I/R injury | Liver | ||
| Kidney defects [113] | NH4Cl | Kidney |
1 Most phenotypes are induced by various challenges. 2 Methods to generate knockout mouse models.
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, Y.J. Knockout Mouse Models for Peroxiredoxins. Antioxidants 2020, 9, 182. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9020182
AMA Style
Lee YJ. Knockout Mouse Models for Peroxiredoxins. Antioxidants. 2020; 9(2):182. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9020182
Chicago/Turabian StyleLee, Young Jae. 2020. "Knockout Mouse Models for Peroxiredoxins" Antioxidants 9, no. 2: 182. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9020182
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.