Analysis of the Status of the Cutaneous Endogenous and Exogenous Antioxidative System of Smokers and the Short-Term Effect of Defined Smoking Thereon


 , and
, and
Abstract
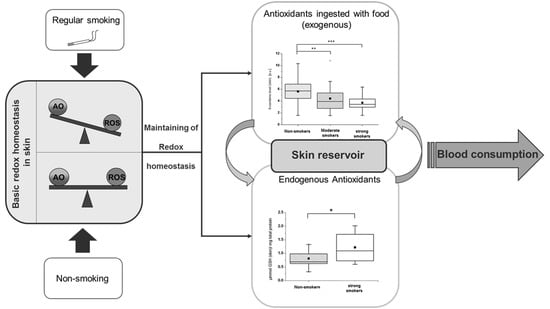
:
1. Introduction
2. Materials and Methods
2.1. Subjects and Study Design
2.2. Determination of the Cutaneous Radical Scavenging Capacity by Electron Paramagnetic Resonance (EPR) Spectroscopy
2.3. Blood Sample Preparation
2.4. Determination of Carotenoids (β-Carotene and Lycopene) in Skin and Blood by Resonance Raman Spectroscopy (RRS)
2.5. Determination of Glutathione (GSH) in Blood and Skin
2.6. Messenger RNA Analysis in Peripheral Blood Mononuclear Cells
2.7. Quantification of Interleukin-22 in Blood Serum
2.8. Data Analysis and Statistical Evaluation
3. Results
3.1. Basic AO Status (0 h)
3.2. AO Status in Response to Defined Smoking
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Huang, M.F.; Lin, W.L.; Ma, Y.C. A study of reactive oxygen species in mainstream of cigarette. Indoor Air 2005, 15, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Pryor, W.A. Biological Effects of Cigarette-Smoke, Wood Smoke, and the Smoke from Plastics-the Use of Electron-Spin-Resonance. Free Radic. Biol. Med. 1992, 13, 659–676. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive Oxygen Species in Metabolic and Inflammatory Signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef] [PubMed]
- Kopa, P.N.; Pawliczak, R. Effect of smoking on gene expression profile-overall mechanism, impact on respiratory system function, and reference to electronic cigarettes. Toxicol. Mech. Methods 2018, 28, 397–409. [Google Scholar] [CrossRef]
- Institute of Medicine. Public Health Implications of Raising the Minimum Age of Legal Access to Tobacco Products; The National Academies Press: Washington, DC, USA, 2015. [Google Scholar] [CrossRef]
- Lephart, E.D. Skin aging and oxidative stress: Equol’s anti-aging effects via biochemical and molecular mechanisms. Ageing Res. Rev. 2016, 31, 36–54. [Google Scholar] [CrossRef]
- Rinnerthaler, M.; Bischof, J.; Streubel, M.K.; Trost, A.; Richter, K. Oxidative stress in aging human skin. Biomolecules 2015, 5, 545–589. [Google Scholar] [CrossRef] [Green Version]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Krinsky, N.I. Mechanism of Action of Biological Antioxidants. Proc. Soc. Exp. Biol. Med. 1992, 200, 248–254. [Google Scholar] [CrossRef]
- Bruno, R.S.; Leonard, S.W.; Atkinson, J.; Montine, T.J.; Ramakrishnan, R.; Bray, T.M.; Traber, M.G. Faster plasma vitamin E disappearance in smokers is normalized by vitamin C supplementation. Free Radic. Biol. Med. 2006, 40, 689–697. [Google Scholar] [CrossRef]
- Bruno, R.S.; Ramakrishnan, R.; Montine, T.J.; Bray, T.M.; Traber, M.G. α-Tocopherol disappearance is faster in cigarette smokers and is inversely related to their ascorbic acid status. Am. J. Clin. Nutr. 2005, 81, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Goralczyk, R. Beta-carotene and lung cancer in smokers: Review of hypotheses and status of research. Nutr. Cancer 2009, 61, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Meinke, M.C.; Lauer, A.; Taskoparan, B.; Gersonde, I.; Lademann, J.; Darvin, M.E. Influence on the Carotenoid Levels of Skin Arising from Age, Gender, Body Mass Index in Smoking/Non-Smoking Individuals. Free Radic. Antioxid. 2011, 1, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L., Jr.; Valanis, B.; Williams, J.H., Jr.; et al. Risk factors for lung cancer and for intervention effects in CARET, the Beta-Carotene and Retinol Efficacy Trial. J. Natl. Cancer Inst. 1996, 88, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Alpha-Tocopherol Beta Carotene Cancer Prevention Study Group. The effect of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers. N. Engl. J. Med. 1994, 330, 1029–1035. [Google Scholar] [CrossRef]
- Anderson, M.E. Glutathione: An overview of biosynthesis and modulation. Chem. Biol. Interact. 1998, 111, 1–14. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Bhatnagar, A.; Tripathi, Y.; Kumar, A. Change in Oxidative Stress of Normotensive Elderly Subjects Following Lifestyle Modifications. J. Clin. Diagn. Res. 2016, 10, CC9–CC13. [Google Scholar] [CrossRef]
- Neurohr, C.; Lenz, A.G.; Ding, I.; Leuchte, H.; Kolbe, T.; Behr, J. Glutamate-cysteine ligase modulatory subunit in BAL alveolar macrophages of healthy smokers. Eur. Respir. J. 2003, 22, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Altraja, S.; Mahlapuu, R.; Soomets, U.; Altraja, A. Cigarette smoke-induced differential regulation of glutathione metabolism in bronchial epithelial cells is balanced by an antioxidant tetrapeptide UPF1. Exp. Toxicol. Pathol. 2013, 65, 711–717. [Google Scholar] [CrossRef]
- Chen, P.X.; Moldoveanu, S.C. Mainstream Smoke Chemical Analyses for 2R4F Kentucky Reference Cigarette. Constributions Tob. Res. 2014, 20, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, J.; Groth, N.; Herrling, T.; Zimmer, G. Electron paramagnetic resonance studies on nitroxide radical 2,2,5,5-tetramethyl-4-piperidin-1-oxyl (TEMPO) redox reactions in human skin. Free Radic. Biol. Med. 1997, 22, 967–976. [Google Scholar] [CrossRef]
- Lauer, A.C.; Groth, N.; Haag, S.F.; Darvin, M.E.; Lademann, J.; Meinke, M.C. Radical scavenging capacity in human skin before and after vitamin C uptake: An in vivo feasibility study using electron paramagnetic resonance spectroscopy. J. Invest. Derm. 2013, 133, 1102–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolk, K.; Mitsui, H.; Witte, K.; Gellrich, S.; Gulati, N.; Humme, D.; Witte, E.; Gonsior, M.; Beyer, M.; Kadin, M.E.; et al. Deficient cutaneous antibacterial competence in cutaneous T-cell lymphomas: Role of Th2-mediated biased Th17 function. Clin. Cancer Res. 2014, 20, 5507–5516. [Google Scholar] [CrossRef] [Green Version]
- Darvin, M.E.; Gersonde, I.; Meinke, M.; Sterry, W.; Lademann, J. Non-invasive in vivo determination of the carotenoids beta-carotene and lycopene concentrations in the human skin using the Raman spectroscopic method. J. Phys. D Appl. Phys. 2005, 38, 2696–2700. [Google Scholar] [CrossRef]
- Meinke, M.C.; Schanzer, S.; Lohan, S.B.; Shchatsinin, I.; Darvin, M.E.; Vollert, H.; Magnussen, B.; Kocher, W.; Helfmann, J.; Lademann, J. Comparison of different cutaneous carotenoid sensors and influence of age, skin type, and kinetic changes subsequent to intake of a vegetable extract. J. Biomed. Opt. 2016, 21, 107002. [Google Scholar] [CrossRef] [PubMed]
- Ermakov, I.V.; Ermakova, M.R.; Gellermann, W.; Lademann, J. Noninvasive selective detection of lycopene and beta-carotene in human skin using Raman spectroscopy. J. Biomed. Opt. 2004, 9, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Hesterberg, K.; Schanzer, S.; Patzelt, A.; Sterry, W.; Fluhr, J.W.; Meinke, M.C.; Lademann, J.; Darvin, M.E. Raman spectroscopic analysis of the carotenoid concentration in egg yolks depending on the feeding and housing conditions of the laying hens. J. Biophotonics 2012, 5, 33–39. [Google Scholar] [CrossRef]
- Cohn, V.H.; Lyle, J. A fluorometric assay for glutathione. Anal. Biochem. 1966, 14, 434–440. [Google Scholar] [CrossRef]
- Hissin, P.J.; Hilf, R. A fluorometric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef]
- Ikeno, H.; Tochio, T.; Tanaka, H.; Nakata, S. Decrease in glutathione may be involved in pathogenesis of acne vulgaris. J. Cosmet. Dermatol. 2011, 10, 240–244. [Google Scholar] [CrossRef]
- Wolk, K.; Frambach, Y.; Jacobi, A.; Wilsmann-Theis, D.; Phillipp, S.; Witte-Handel, E.; Wenzel, J.; Mossner, R.; Sabat, R. Increased levels of lipocalin 2 in palmoplantar pustular psoriasis. J. Dermatol. Sci. 2018, 90, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Cheng, Z.; Liu, W.; Wu, K. Expression of interleukin (IL)-10, IL-17A and IL-22 in serum and sputum of stable chronic obstructive pulmonary disease patients. COPD 2013, 10, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Sabat, R.; Ouyang, W.; Wolk, K. Therapeutic opportunities of the IL-22-IL-22R1 system. Nat. Rev. Drug Discov. 2014, 13, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Lademann, J.; Kocher, W.; Yu, R.; Meinke, M.C.; Na Lee, B.; Jung, S.; Sterry, W.; Darvin, M.E. Cutaneous carotenoids: The mirror of lifestyle? Skin Pharmacol. Physiol. 2014, 27, 201. [Google Scholar] [CrossRef] [PubMed]
- Koster, M.I. Making an epidermis. Ann. N. Y. Acad. Sci. 2009, 1170, 7–10. [Google Scholar] [CrossRef]
- Thiele, J.J.; Schroeter, C.; Hsieh, S.N.; Podda, M.; Packer, L. The antioxidant network of the stratum corneum. Curr. Probl. Dermatol. 2001, 29, 26–42. [Google Scholar] [CrossRef]
- Ighodaro, O.M. Molecular pathways associated with oxidative stress in diabetes mellitus. Biomed. Pharm. 2018, 108, 656–662. [Google Scholar] [CrossRef]
- Laaksonen, D.E.; Atalay, M.; Niskanen, L.; Uusitupa, M.; Hanninen, O.; Sen, C.K. Blood glutathione homeostasis as a determinant of resting and exercise-induced oxidative stress in young men. Redox Rep. 1999, 4, 53–59. [Google Scholar] [CrossRef]
- Elokda, A.S.; Nielsen, D.H. Effects of exercise training on the glutathione antioxidant system. Eur. J. Cardiovasc. Prev. Rehabil. 2007, 14, 630–637. [Google Scholar] [CrossRef]
- Cantin, A.M. Cellular response to cigarette smoke and oxidants: Adapting to survive. Proc. Am. Thorac. Soc. 2010, 7, 368–375. [Google Scholar] [CrossRef]
- Carolan, B.J.; Harvey, B.G.; Hackett, N.R.; O’Connor, T.P.; Cassano, P.A.; Crystal, R.G. Disparate oxidant gene expression of airway epithelium compared to alveolar macrophages in smokers. Respir. Res. 2009, 10, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohan, S.B.; Vitt, K.; Scholz, P.; Keck, C.M.; Meinke, M.C. ROS production and glutathione response in keratinocytes after application of beta-carotene and VIS/NIR irradiation. Chem. Biol. Interact. 2018, 280, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bacic, G.; Pavicevic, A.; Peyrot, F. In vivo evaluation of different alterations of redox status by studying pharmacokinetics of nitroxides using magnetic resonance techniques. Redox Biol. 2016, 8, 226–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlberg, S.; Rancan, F.; Epple, M.; Loza, K.; Hoppe, D.; Lademann, J.; Vogt, A.; Kleuser, B.; Gerecke, C.; Meinke, M.C. Comparison of different methods to study effects of silver nanoparticles on the pro- and antioxidant status of human keratinocytes and fibroblasts. Methods 2016, 109, 55–63. [Google Scholar] [CrossRef]
- Elpelt, A.; Ivanov, D.; Novackova, A.; Kovacik, A.; Sochorova, M.; Saeidpour, S.; Teutloff, C.; Lohan, S.B.; Lademann, J.; Vavrova, K.; et al. Investigation of TEMPO partitioning in different skin models as measured by EPR spectroscopy-Insight into the stratum corneum. J. Magn. Reson. 2019, 310, 106637. [Google Scholar] [CrossRef]
- Fuchs, J.; Freisleben, H.J.; Podda, M.; Zimmer, G.; Milbradt, R.; Packer, L. Nitroxide Radical Biostability in Skin. Free Radic. Biol. Med. 1993, 15, 415–423. [Google Scholar] [CrossRef]
- Bizon, A.; Milnerowicz, H. Effect of tobacco smoking on glutathione concentration in the blood. Prz. Lek. 2012, 69, 809–811. [Google Scholar]
- Garg, N.; Singh, R.; Dixit, J.; Jain, A.; Tewari, V. Levels of lipid peroxides and antioxidants in smokers and nonsmokers. J. Periodontal Res. 2006, 41, 405–410. [Google Scholar] [CrossRef]
- Teichmann, A.; Jacobi, U.; Weigmann, H.J.; Sterry, W.; Lademann, J. Reservoir function of the stratum corneum: Development of an in vivo method to quantitatively determine the stratum corneum reservoir for topically applied substances. Skin Pharm. Physiol. 2005, 18, 75–80. [Google Scholar] [CrossRef]
- Farris, P.K.; Krol, Y. Under Persistent Assault: Understanding the Factors that Deteriorate Human Skin and Clinical Efficacy of Topical Antioxidants in Treating Aging Skin. Cosmetics 2015, 2, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the Most Efficient Biological Carotenoid Singlet Oxygen Quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharm. 2013, 75, 588–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vierkotter, A.; Krutmann, J. Environmental influences on skin aging and ethnic-specific manifestations. Dermatoendocrinol 2012, 4, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Morita, A.; Torii, K.; Maeda, A.; Yamaguchi, Y. Molecular basis of tobacco smoke-induced premature skin aging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 53–55. [Google Scholar] [CrossRef]
- Malila, N.; Taylor, P.R.; Virtanen, M.J.; Korhonen, P.; Huttunen, J.K.; Albanes, D.; Virtamo, J. Effects of alpha-tocopherol and beta-carotene supplementation on gastric cancer incidence in male smokers (ATBC Study, Finland). Cancer Causes Control 2002, 13, 617–623. [Google Scholar] [CrossRef]
- Albanes, D.; Heinonen, O.P.; Taylor, P.R.; Virtamo, J.; Edwards, B.K.; Rautalahti, M.; Hartman, A.M.; Palmgren, J.; Freedman, L.S.; Haapakoski, J.; et al. Alpha-Tocopherol and beta-carotene supplements and lung cancer incidence in the alpha-tocopherol, beta-carotene cancer prevention study: Effects of base-line characteristics and study compliance. J. Natl. Cancer Inst. 1996, 88, 1560–1570. [Google Scholar] [CrossRef]
- Bardia, A.; Tleyjeh, I.M.; Cerhan, J.R.; Sood, A.K.; Limburg, P.J.; Erwin, P.J.; Montori, V.M. Efficacy of antioxidant supplementation in reducing primary cancer incidence and mortality: Systematic review and meta-analysis. Mayo Clin. Proc. 2008, 83, 23–34. [Google Scholar] [CrossRef]
- Penning, T.M. Aldo-Keto Reductase Regulation by the Nrf2 System: Implications for Stress Response, Chemotherapy Drug Resistance, and Carcinogenesis. Chem. Res. Toxicol. 2017, 30, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Hackett, N.R.; Heguy, A.; Harvey, B.G.; O’Connor, T.P.; Luettich, K.; Flieder, D.B.; Kaplan, R.; Crystal, R.G. Variability of antioxidant-related gene expression in the airway epithelium of cigarette smokers. Am. J. Respir. Cell Mol. Biol. 2003, 29, 331–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolk, K.; Kunz, S.; Witte, E.; Friedrich, M.; Asadullah, K.; Sabat, R. IL-22 increases the innate immunity of tissues. Immunity 2004, 21, 241–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radaeva, S.; Sun, R.; Pan, H.N.; Hong, F.; Gao, B. Interleukin 22 (IL-22) plays a protective role in T cell-mediated murine hepatitis: IL-22 is a survival factor for hepatocytes via STAT3 activation. Hepatology 2004, 39, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Ki, S.H.; Park, O.; Zheng, M.; Morales-Ibanez, O.; Kolls, J.K.; Bataller, R.; Gao, B. Interleukin-22 treatment ameliorates alcoholic liver injury in a murine model of chronic-binge ethanol feeding: Role of signal transducer and activator of transcription 3. Hepatology 2010, 52, 1291–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sub-Study | Smoking Status | Number of Participants | Gender: Female/Male | Analyzed Parameters |
|---|---|---|---|---|
| A | nSm | 57 | 33/24 | Basic (0 h) AO status in skin: radical scavenging capacity, carotenoids |
| mSm | 18 | 7/11 | ||
| sSm | 23 | 9/14 | ||
| B | nSm | 13 | 9/4 | Basic GSH (0 h) status in skin |
| mSm | 0 | n.a. | ||
| sSm | 13 | 5/8 | ||
| C | nSm | 10 | 5/5 | AO status 4 h after defined cigarette consumption compared to basis (0 h) status in skin and blood plasma of sSm: scavenging capacity, carotenoids, GSH and IL-22 (blood only) |
| mSm | 0 | n.a. | ||
| sSm | 10 | 5/5 | ||
| D | nSm # | 5 | 4/1 | Basic status of expression of oxidative stress-associated genes in blood cells and its comparison to the status 4 h after defined cigarette consumption in sSm |
| mSm | 0 | n.a. | ||
| sSm # | 5 | 3/2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lohan, S.B.; Bühring, K.; Lauer, A.-C.; Friedrich, A.; Lademann, J.; Buss, A.; Sabat, R.; Wolk, K.; Meinke, M.C. Analysis of the Status of the Cutaneous Endogenous and Exogenous Antioxidative System of Smokers and the Short-Term Effect of Defined Smoking Thereon. Antioxidants 2020, 9, 537. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9060537
Lohan SB, Bühring K, Lauer A-C, Friedrich A, Lademann J, Buss A, Sabat R, Wolk K, Meinke MC. Analysis of the Status of the Cutaneous Endogenous and Exogenous Antioxidative System of Smokers and the Short-Term Effect of Defined Smoking Thereon. Antioxidants. 2020; 9(6):537. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9060537
Chicago/Turabian StyleLohan, Silke B., Karl Bühring, Anna-Christina Lauer, Annette Friedrich, Jürgen Lademann, Annette Buss, Robert Sabat, Kerstin Wolk, and Martina C. Meinke. 2020. "Analysis of the Status of the Cutaneous Endogenous and Exogenous Antioxidative System of Smokers and the Short-Term Effect of Defined Smoking Thereon" Antioxidants 9, no. 6: 537. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9060537