Insights into Arbutin Effects on Bone Cells: Towards the Development of Antioxidant Titanium Implants

 , ,
, ,  , , and
, , and 
Abstract
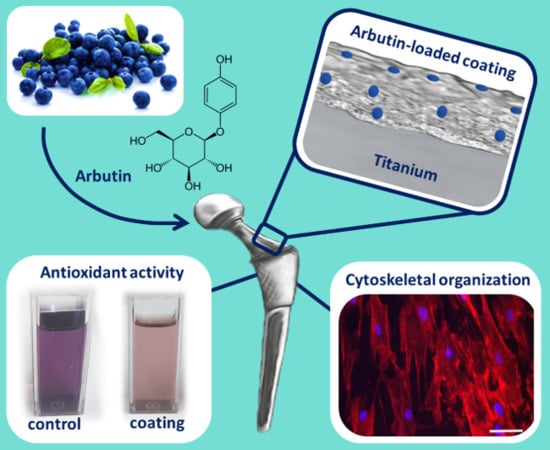
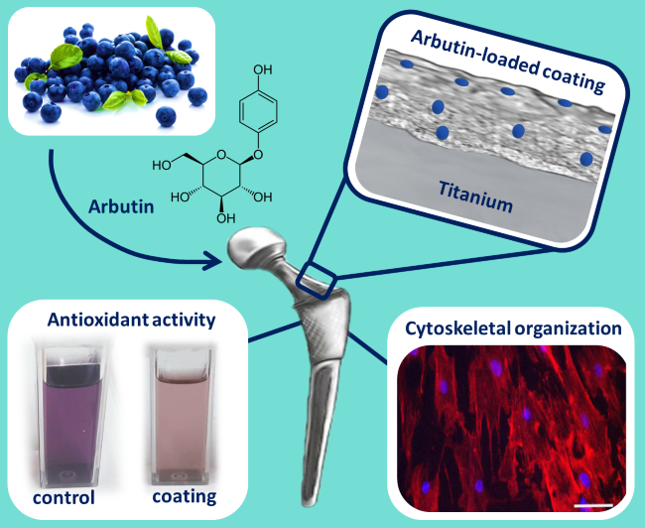
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Coating Preparation
2.3. Arbutin Loading Procedures
2.4. X-ray Photoelectron Spectroscopy (XPS)
2.5. Scanning Electron Microscopy (SEM) Morphological Analysis of the Coatings
2.6. High-Performance Liquid Chromatography (HPLC)
2.7. DPPH Assay
2.8. Cell Culture, Cytocompatibility and Antioxidant Activity Assessment
2.8.1. Cell Cultures
2.8.2. Arbutin Treatment
2.8.3. Material Seeding
2.8.4. MTT (3-Dimethylthiazol-2,5-diphenyltetrazolium bromide) Colorimetric Assay
2.8.5. qRT-PCR
2.8.6. Western Blot Analysis
2.8.7. Fluorescence Microscopy
2.9. Statistical Analyses
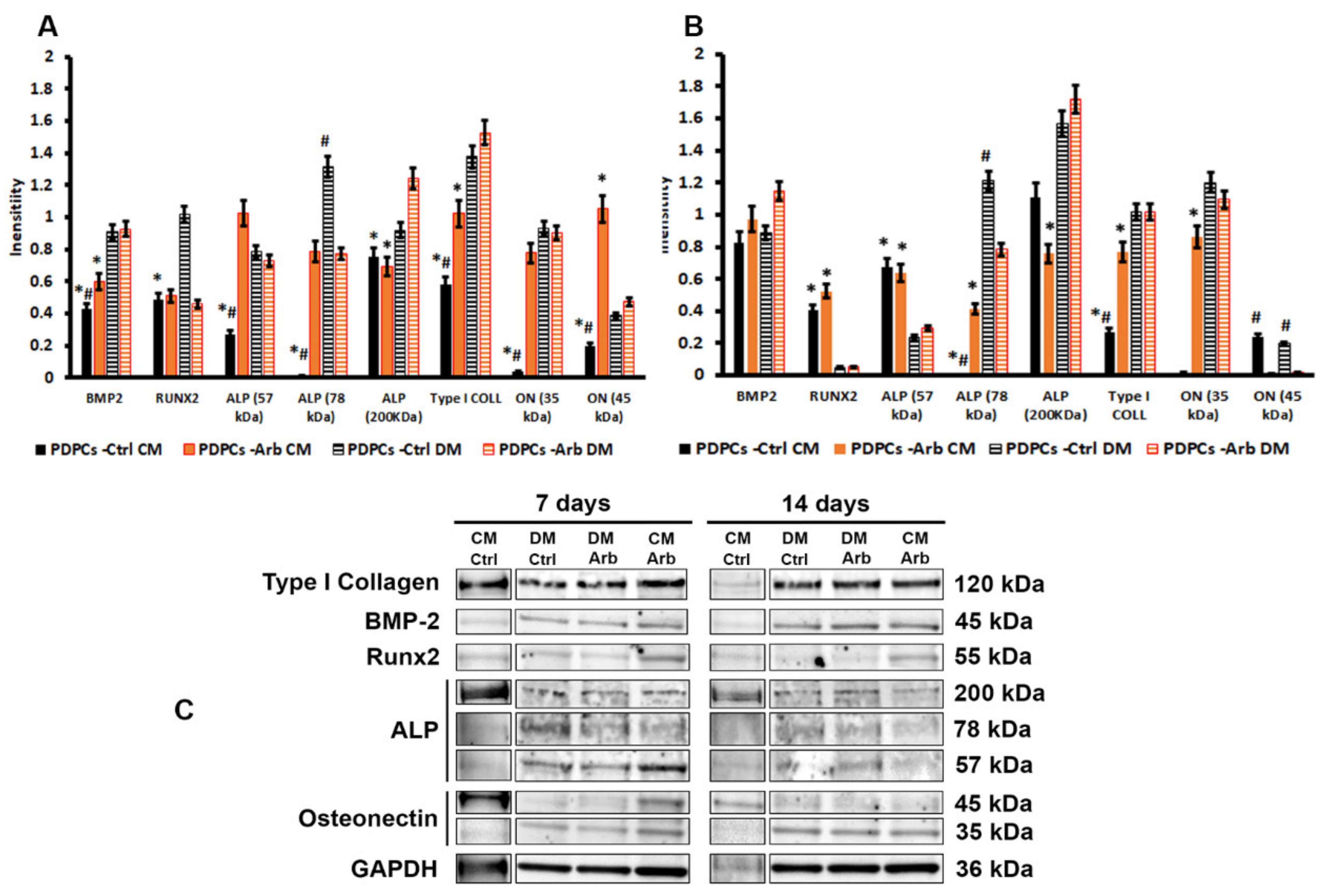
3. Results and Discussion
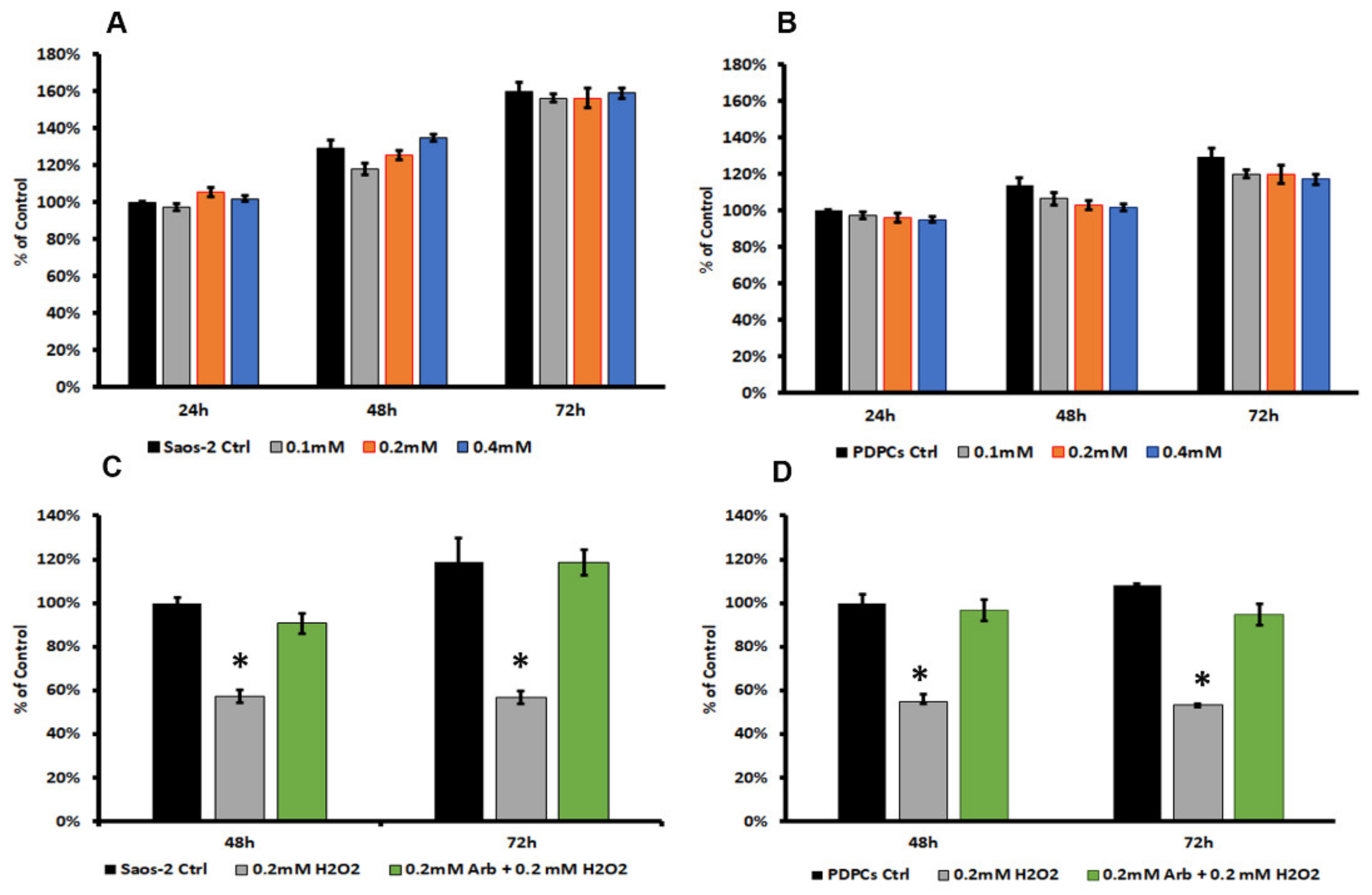
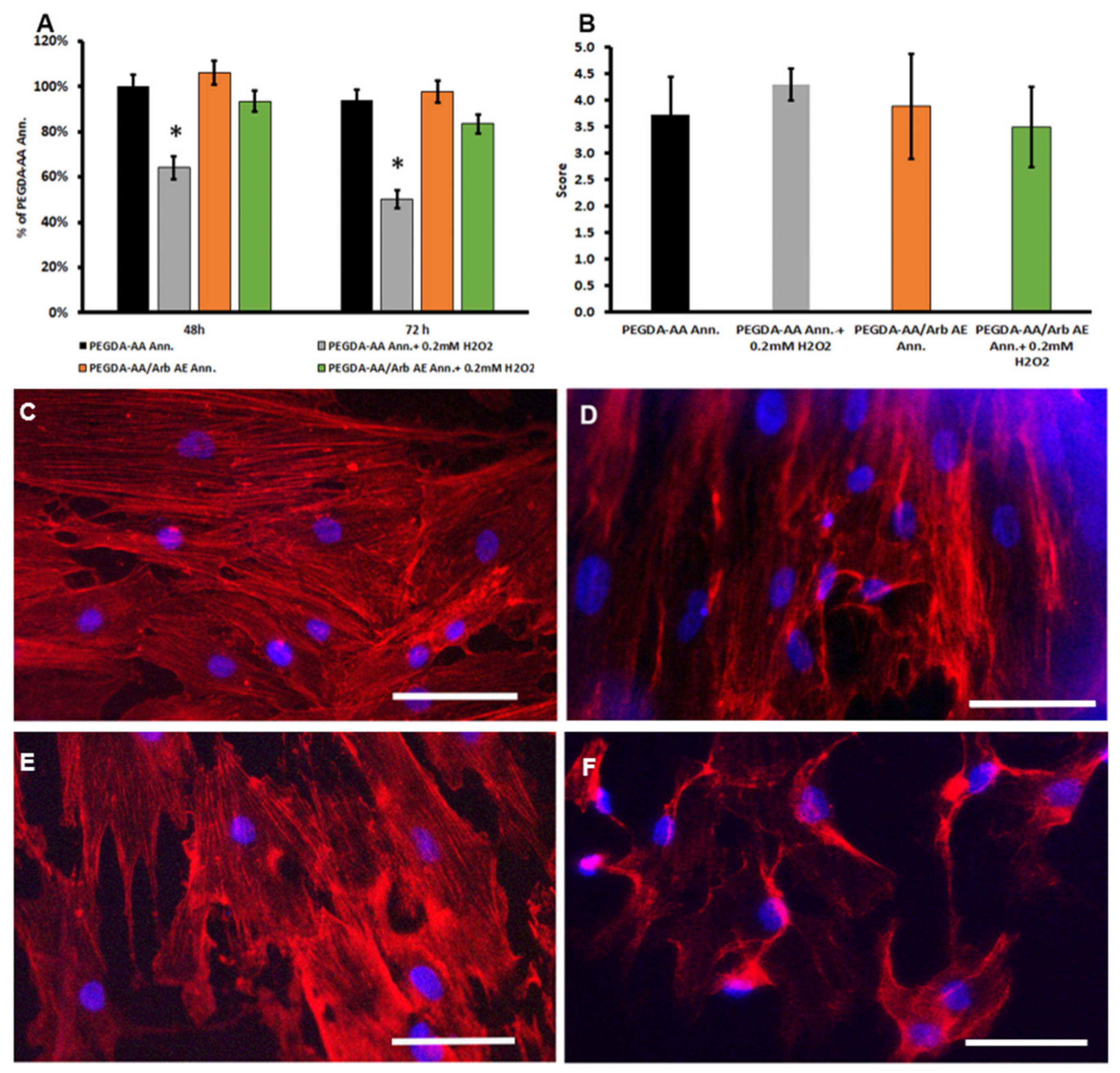
3.1. Arbutin Cytocompatibility and Antioxidant Activity
3.2. Coating Preparation, Morphological and Physicochemical Characterization
3.2.1. Electrochemical Preparation of PEGDA-AA/Arb Coatings on Titanium
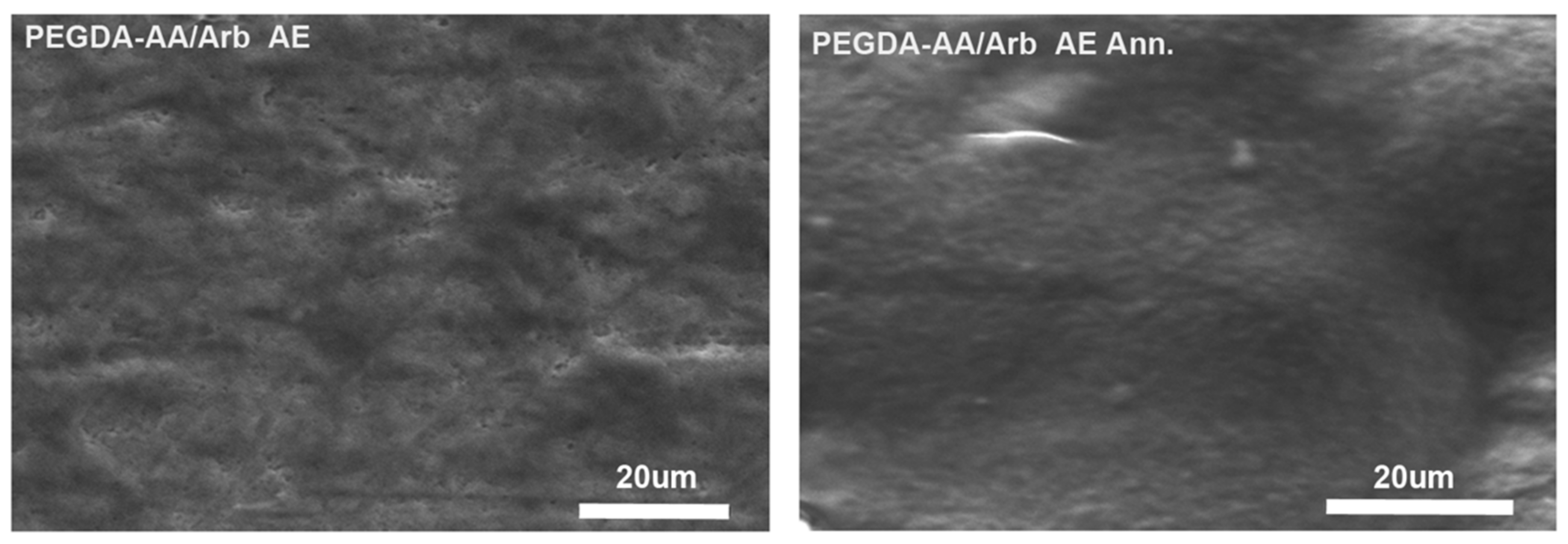
3.2.2. Scanning Electron Microscopy of the Coatings
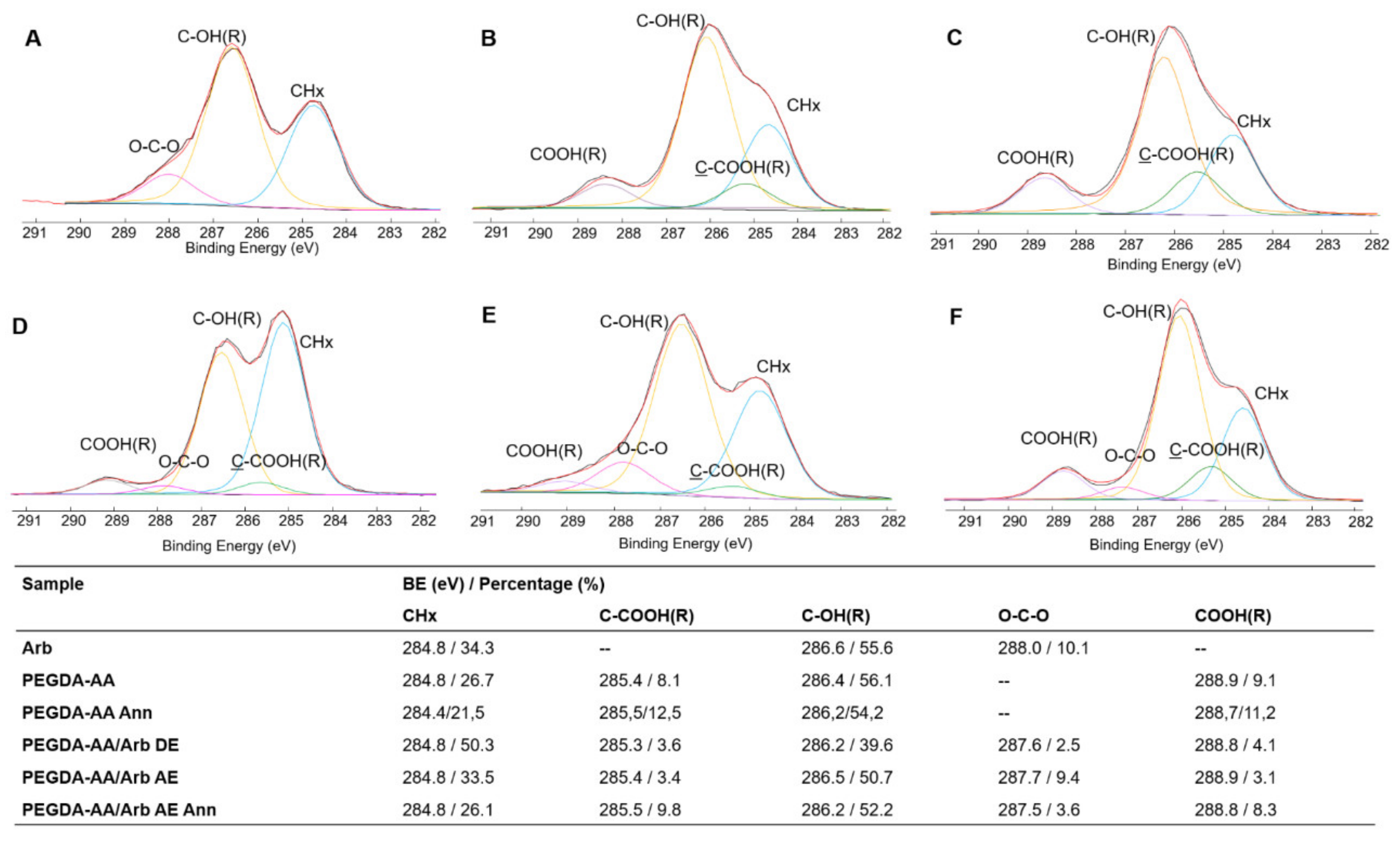
3.2.3. XPS Analysis of the Coatings
3.2.4. Arbutin Quantification by High-Performance Liquid Chromatography (HPLC)
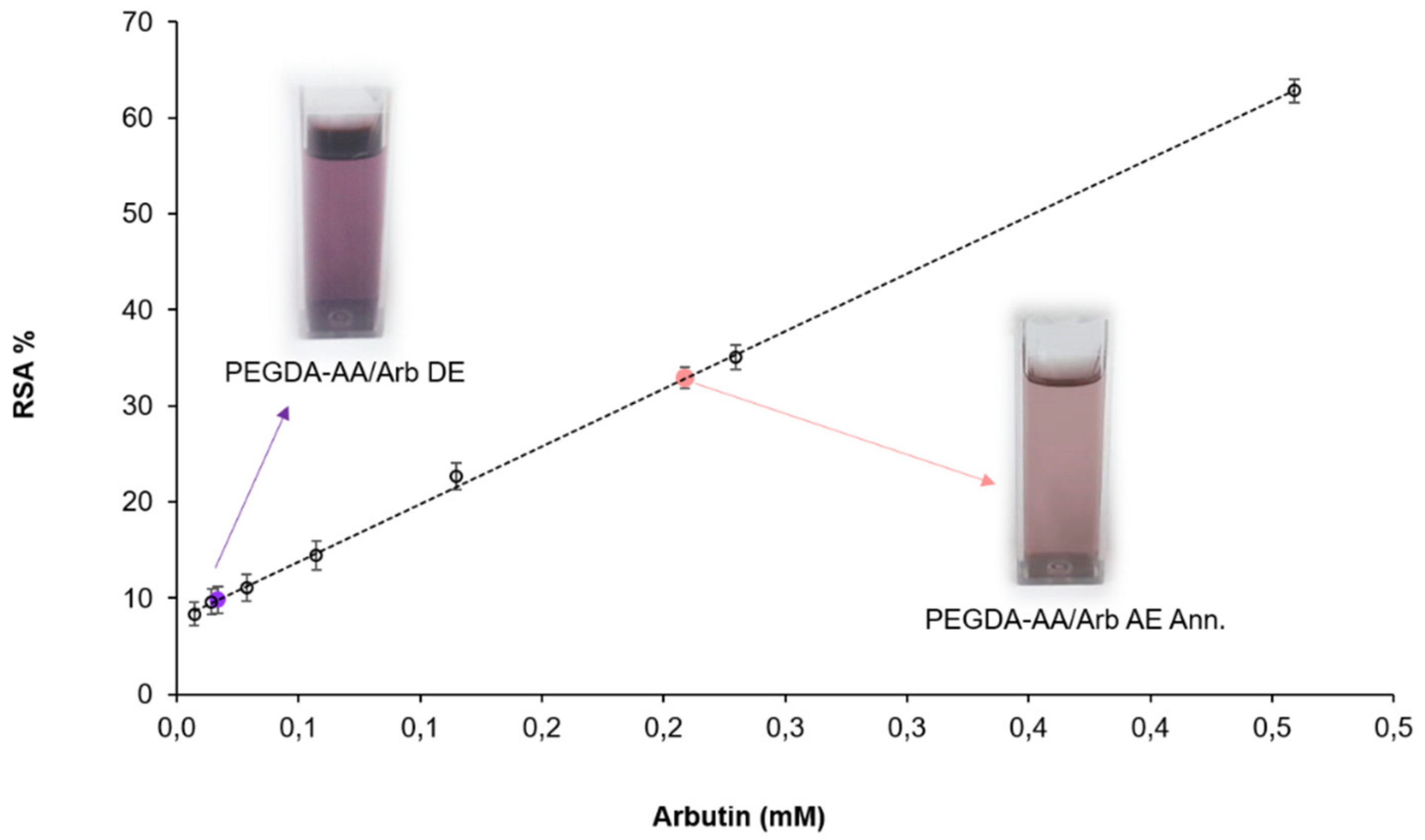
3.3. Antioxidant Activity Evaluations by DPPH Assay
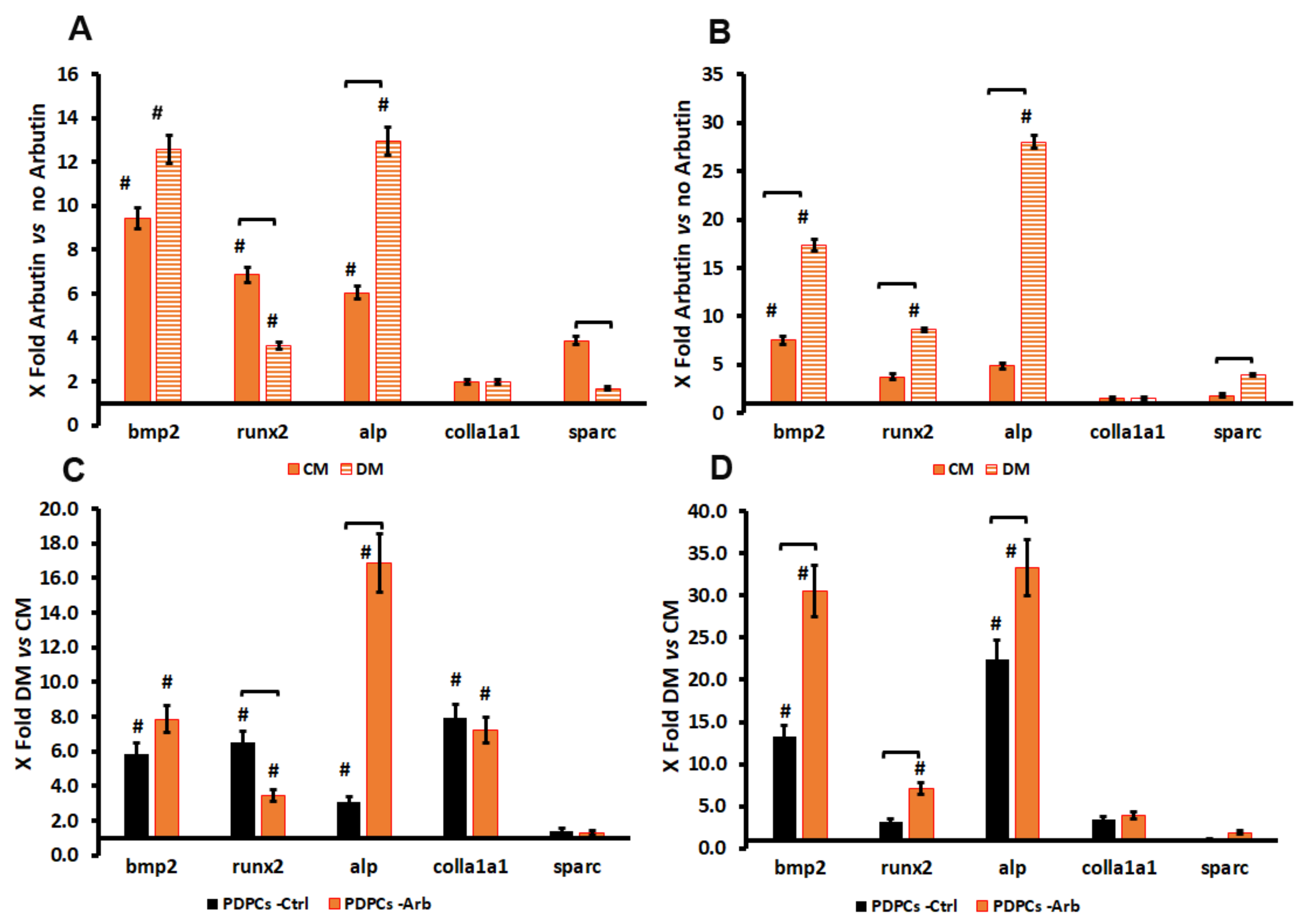
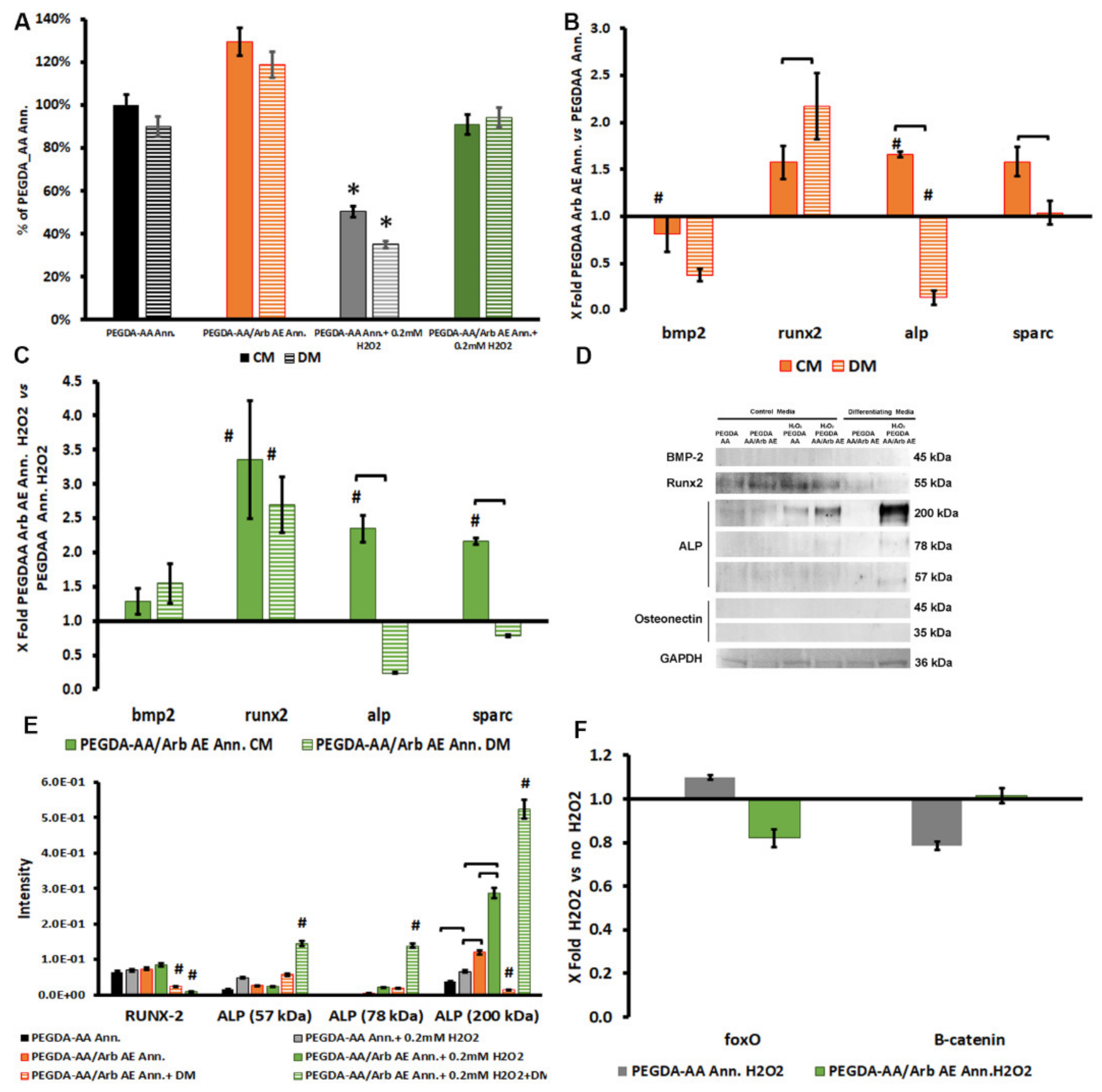
3.4. Arbutin-Loaded Coating Cytocompatibility and Anti-Oxidant Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Acrylic acid |
| ALP | Alkaline phosphatase |
| BMP2 | Bone morphogenetic protein 2 |
| Coll1 | Collagen type I |
| CM | Complete medium, i.e., DMEM with 10% FBS |
| DM | Differentiating medium, i.e., DMEM supplemented with the osteogenic factors: dexamethasone, ascorbic acid and β-glycerophosphate |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DPPH assay | 2,2-difenil-1-picrylhydrazyl assay |
| FBS | Fetal bovine serum |
| FoxO | Forkhead box O |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GUSB | Beta-glucuronidase |
| HPLC | High-performance liquid chromatography |
| MSCs | Mesenchymal stem cells |
| MTT | 3-dimethylthiazol-2,5-diphenyltetrazolium bromide |
| OB | Osteoblasts |
| ON | Osteonectin–protein |
| PDPCs | Periosteal-derived precursor cells |
| PEGDA | Poly(ethylene-glycol diacrylate) |
| PEGDA-AA | Poly(acrylic acid)–poly(ethylene-glycol diacrylate) coating |
| PEGDA-AA/Arb AE | Poly(acrylic acid)–poly(ethylene-glycol diacrylate) coating with arbutin embedded after electrosynthesis |
| PEGDA-AA/Arb DE | Poly(acrylic acid)–poly(ethylene-glycol diacrylate) coating with arbutin embedded during electrosynthesis |
| RANK-L | Receptor activator of nuclear factor kappa-Β ligand |
| RSA % | Radical scavenging activity percentage |
| ROS | Reactive oxygen species |
| RUNX-2 | Runt-related transcription factor 2 |
| Saos-2 | Human osteosarcoma cell line |
| SEM | Scanning electron microscopy |
| Sparc | Secreted protein, acidic, cysteine-rich–osteonectin gene |
| TCF | T-cell factor |
| TGA | Thermogravimetric analysis |
| XPS | X-ray photoelectron spectroscopy |
| Wnt | Wingless-related integration site |
References
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Miner. Bone Metab. 2017, 14, 209–216. [Google Scholar] [CrossRef]
- Zukowski, P.; Maciejczyk, M.; Waszkiel, D. Sources of free radicals and oxidative stress in the oral cavity. Arch. Oral Biol. 2018, 92, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Storz, P. Forkhead homeobox type O transcription factors in the responses to oxidative stress. Antioxid. Redox Signal. 2011, 14, 593–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasote, D.M.; Katyare, S.S.; Hedge, M.V.; Bae, H. Significance of antioxidant potential of plants and its relevance to therapeutic applications. Int. J. Biol. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustarichie, R.; Gozali, D. Formulation and evaluation of alpha arbutin skin lightening cream using polyacrylate base by cold process. Int. J. App. Pharm. 2019, 11, 100–105. [Google Scholar] [CrossRef]
- Jurica, K.; Gobin, I.; Kremer, D.; Čepo, D.V.; Grubešić, R.J.; Karačonji, I.B.; Kosalec, I. Arbutin and its metabolite hydroquinone as the main factors in the antimicrobial effect of strawberry tree (Arbutus unedo L.) leaves. J. Herb. Med. 2017, 8, 17–23. [Google Scholar] [CrossRef]
- Dadgar, M.; Pouramir, M.; Dastan, Z.; Ghasemi-Kasman, M.; Ashrafpour, M.; Moghadamnia, A.A.; Khafri, S.; Pourghasem, M. Arbutin attenuates behavioral impairment and oxidative stress in an animal model of Parkinson’s disease. Avicenna J. Phytomed. 2018, 8, 533–542. [Google Scholar]
- Zhao, W.; Wang, S.; Qin, T.; Wang, W. Arbutin attenuates hydrogen peroxide-induced oxidative injury through regulation of microRNA-29a in retinal ganglion cells. Biomed. Pharmacother. 2019, 112, 108729. [Google Scholar] [CrossRef]
- Torre, E. Molecular signalling mechanisms behind polyphenol-induced bone anabolism. Phytochem. Rev. 2017, 16, 1183–1226. [Google Scholar] [CrossRef]
- Omori, A.; Yoshimura, Y.; Deyama, Y.; Suzuki, K. Rosmarinic acid and arbutin suppress osteoclast differentiation by inhibiting superoxide and NFATc1 downregulation in RAW 264.7 cells. Biomed. Rep. 2015, 3, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, X.; Yang, L.; Liu, S.; Yang, L.; Li, M.; Fu, Q. Arbutin promotes MC3T3-E1 mouse osteoblast precursor cell proliferation and differentiation via the Wnt/β-catenin signaling pathway. Mol. Med. Rep. 2019, 19, 4637–4644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindler, G.; Patzak, U.; Brinkhaus, B.; von Niecieck, A.; Wittig, J.; Kramer, N.; Glockl, I.; Veit, M. Urinary excretion and metabolism of arbutin after oral administration of arctostaphylos uvae ursi extract as film-coated tablets and aqueous solution in healthy humans. J. Clin. Pharmacol. 2002, 42, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Córdoba, A.; Satué, M.; Gómez-Florit, M.; Monjo, M.; Ramis, J.M. Flavonoid coated titanium surfaces for bioactive bone implants. Stem Cell Transl. Investig. 2015, 2, e520. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Shen, X.; Hu, Y.; Xu, K.; Ran, Q.; Yu, Y.; Dai, L.; Yuan, Z.; Huang, L.; Shen, T.; et al. Surface functionalization of Titanium implants with chitosan-catechol conjugate for suppression of ROS-induced cells damage and improvement of osteogenesis. Biomaterials 2017, 114, 82–96. [Google Scholar] [CrossRef]
- De Giglio, E.; Cometa, S.; Satriano, C.; Sabbatini, L.; Zambonin, P.G. Electrosynthesis of hydrogel films on metal substrates for the development of coatings with tunable drug delivery performances. J. Biomed. Mater. Res. A 2008, 88, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- De Giglio, E.; Cometa, S.; Ricci, M.A.; Cafagna, D.; Savino, A.M.; Sabbatini, L.; Orciani, M.; Ceci, E.; Novello, L.; Tantillo, G.M.; et al. Ciprofloxacin-modified electrosynthesized hydrogel coatings to prevent Titanium-implant associated infections. Acta Biomater. 2011, 7, 882–891. [Google Scholar] [CrossRef] [PubMed]
- De Giglio, E.; Cometa, S.; Cioffi, N.; Torsi, L.; Sabbatini, L. Analytical investigations of poly (acrylic acid) coatings electrodeposited on Titanium-based implants: A versatile approach to biocompatibility enhancement. Anal. Bioanal. Chem. 2007, 389, 2055–2063. [Google Scholar] [CrossRef] [PubMed]
- Muchtaridi, M.; Musfiroh, I.; Fauzi, A. Method development and validation for analysis of deoxyarbutinin anhydrous emulsion system using high-performance liquid chromatography. Int. J. Appl. Pharm. 2019, 11, 172–175. [Google Scholar] [CrossRef]
- Prakash, O.; Revathy, B.; Kudachikar, V.B. Characterization, quantification of free, esterified and bound phenolics in Kainth (Pyrus pashia Buch.-Ham. Ex, D. Don) fruit pulp by UPLC-ESI-HRMS/MS and evaluation of their antioxidant activity. Food Chem. 2019, 299, 125114. [Google Scholar] [CrossRef]
- Shrikanta, A.; Kumar, A.; Govindaswamy, V. Resveratrol content and antioxidant properties of underutilized fruits. J. Food Sci. Technol. 2015, 52, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, C.; Borsari, V.; Falconi, M.; Gigante, A.; Lazzarini, R.; Fini, M.; Di Primio, R.; Mattioli-Belmonte, M. Human periosteum-derived stem cells for tissue engineering applications: The role of VEGF. Stem Cell Rev. Rep. 2012, 8, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for defining multipotent mesenchymal stem cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Ciuffreda, M.C.; Malpasso, G.; Musarò, P.; Turco, V.; Gnecchi, M. Protocols for in vitro differentiation of human mesenchymal stem cells into Osteogenic, Chondrogenic and Adipogenic lineages. In Mesenchymal Stem Cells; Humana Press: New York, NY, USA, 2016; Volume 1416, pp. 149–158. [Google Scholar] [CrossRef]
- Ragni, E.; Viganò, M.; Rebulla, P.; Giordano, R.; Lazzari, L. What is beyond a qRT-PCR study on mesenchymal stem cell differentiation properties: How to choose the most reliable housekeeping genes. J. Cell. Mol. Med. 2013, 17, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hempel, U.; Hefti, T.; Kalbacova, M.; Wolf-Brandstetter, C.; Dieter, P.; Schlottig, F. Response of osteoblast-like SAOS-2 cells to zirconia ceramics with different surface topographies. Clin. Oral Implant. Res. 2010, 21, 174–181. [Google Scholar] [CrossRef]
- Collins, F.L.; Rios-Arce, N.D.; Schepper, J.D.; Parameswaran, N.; McCabe, L.R. The potential of probiotics as a therapy for osteoporosis. Microbiol Spectr. 2017, 5, 213–233. [Google Scholar] [CrossRef]
- Freeman, F.E.; Stevens, H.Y.; Owens, P.; Guldberg, R.E.; McNamara, L.M. Osteogenic differentiation of mesenchymal stem cells by mimicking the cellular niche of the endochondral template. Tissue Eng. Part A 2016, 22, 1176–1190. [Google Scholar] [CrossRef]
- Nakamura, T.; Nakamura, K.; Stinson, R.A. Release of alkaline phosphatase from human osteosarcoma cells by phosphatidylinositol phospholipase C: Effect of tunicamycin. Arch. Biochem. Biophys. 1988, 265, 190–196. [Google Scholar] [CrossRef]
- Numa-Kinjoh, N.; Komaru, K.; Ishida, Y.; Sohda, M.; Oda, K. Molecular phenotype of tissue-nonspecific alkaline phosphatase with a proline (108) to leucine substitution associated with dominant odontohypophosphatasia. Mol. Genet. Metab. 2015, 115, 180–185. [Google Scholar] [CrossRef]
- Al-Shawafi, H.A.; Komaru, K.; Oda, K. Molecular defect of tissue-nonspecific alkaline phosphatase bearing a substitution at position 426 associated with hypophosphatasia. Mol. Cell Biochem. 2017, 427, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Romberg, R.W.; Werness, P.G.; Lollar, P.; Riggs, B.L.; Mann, K.G. Isolation and characterization of native adult osteonectin. J. Biol. Chem. 1985, 260, 2728–2736. [Google Scholar]
- Cometa, S.; Mattioli-Belmonte, M.; Cafagna, D.; Iatta, R.; Ceci, E.; De Giglio, E. Antibiotic-modified hydrogel coatings on titanium dental implants. J. Biol. Regul. Homeost. Agents 2012, 26, 65–71. [Google Scholar] [PubMed]
- Catauro, M.; Bollino, F.; Papale, F.; Piccolella, S.; Pacifico, S. Sol–gel synthesis and characterization of SiO2/PCL hybrid materials containing quercetin as new materials for antioxidant implants. Mater. Sci. Eng. C 2016, 58, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Mattila, P.K.; Lappalainen, P. Filopodia: Molecular architecture and cellular functions. Nat. Rev. Mol. Cell Biol. 2008, 9, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Bilezikian, J.P.; Raisz, L.G.; Rodan, G.A. Principles of Bone Biology, 2nd ed.; Academic Press: San Diego, CA, USA, 2002; Volume 1. [Google Scholar]
- Balcerzak, M.; Hamade, E.; Zhang, L.; Pikula, S.; Azzar, G.; Radisson, J.; Bandorowicz-Pikula, J.; Buchet, R. The roles of annexins and alkaline phosphatase in mineralization process. Acta Biochim. Pol. 2003, 50, 1019–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaikov, G.E.; Valente, A.J.M.; Iordanskii, A.L. Advances in Kinetics and Mechanism of Chemical Reactions; CRC Press: Toronto, ON, Canada, 2013. [Google Scholar]
- Wang, P.Y.; Clements, L.R.; Thissen, H.; Tsai, W.B.; Voelcker, N.H. Screening rat mesenchymal stem cell attachment and differentiation on surface chemistries using plasma polymer gradients. Acta Biomater. 2015, 11, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.; Ambrogini, E.; Bartell, S.M.; Han, L.; Roberson, P.K.; de Cabo, R.; Almeida, M. FOXOs attenuate bone formation by suppressing Wnt signaling. J. Clin. Investig. 2013, 123, 3409–3419. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Han, L.; Martin-Millan, M.; O’Brien, C.A.; Manolagas, S.C. Oxidative stress antagonizes Wnt signaling in osteoblast precursors by diverting β-catenin from T cell factor-to forkhead box O-mediated transcription. J. Biol. Chem. 2007, 282, 27298–27305. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Denu, R.A.; Hematti, P. Effects of oxidative stress on mesenchymal stem cell biology. Oxidative Med. Cell. Longev. 2016, 314, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferretti, C.; Vozzi, G.; Falconi, M.; Orciani, M.; Gesi, M.; Di Primio, R.; Mattioli-Belmonte, M. Role of IGF1 and IGF1/VEGF on human mesenchymal stromal cells in bone healing: Two sources and two fates. Tissue Eng. Part A 2014, 20, 2473–2482. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Lu, D.; Bai, J.; Zheng, H.; Ke, Z.; Li, X.; Luo, S. Oxidative stress inhibits osteoblastic differentiation of bone cells by ERK and NF-κB. Biochem. Biophys. Res. Commun. 2004, 314, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Gong, Y.; Xu, L.; Zhou, M.; Li, J.; Song, J. Bidirectional regulation of osteogenic differentiation by the FOXO subfamily of Forkhead transcription factors in mammalian MSCs. Cell Prolif. 2019, 52, e12540. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Atomic Percentages (%) | |
|---|---|---|
| C1s | O1s | |
| Arb | 63.4 | 36.6 |
| PEGDA-AA | 67.6 | 32.4 |
| PEGDA-AA Ann. | 66.1 | 33.9 |
| PEGDA-AA/Arb DE | 72.8 | 23.6 |
| PEGDA-AA/Arb AE | 65.8 | 34.2 |
| PEGDA-AA/Arb AE Ann. | 69.9 | 30.1 |
| Sample | Loaded Arbutin (mM) |
|---|---|
| PEGDA-AA/Arb DE | 0.019 ± 0.002 |
| PEGDA-AA/Arb AE 1 h | 0.220 ± 0.004 |
| PEGDA-AA/Arb AE 1 h Ann. | 0.206 ± 0.003 |
| PEGDA-AA/Arb AE 3 h | 0.41 ± 0.09 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonifacio, M.A.; Cerqueni, G.; Cometa, S.; Licini, C.; Sabbatini, L.; Mattioli-Belmonte, M.; De Giglio, E. Insights into Arbutin Effects on Bone Cells: Towards the Development of Antioxidant Titanium Implants. Antioxidants 2020, 9, 579. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9070579
Bonifacio MA, Cerqueni G, Cometa S, Licini C, Sabbatini L, Mattioli-Belmonte M, De Giglio E. Insights into Arbutin Effects on Bone Cells: Towards the Development of Antioxidant Titanium Implants. Antioxidants. 2020; 9(7):579. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9070579
Chicago/Turabian StyleBonifacio, Maria A., Giorgia Cerqueni, Stefania Cometa, Caterina Licini, Luigia Sabbatini, Monica Mattioli-Belmonte, and Elvira De Giglio. 2020. "Insights into Arbutin Effects on Bone Cells: Towards the Development of Antioxidant Titanium Implants" Antioxidants 9, no. 7: 579. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9070579