
Host Responses Following Infection with Canadian-Origin Wildtype and Vaccine Revertant Infectious Laryngotracheitis Virus

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Eggs
2.2. Virus
2.3. Experimental Design
2.4. Techniques
2.4.1. DNA Extraction
2.4.2. RNA Extraction and Reverse Transcription
2.4.3. ILTV Genome Load Quantification and Cytokine mRNA Expression by Quantitative (q)PCR Assays
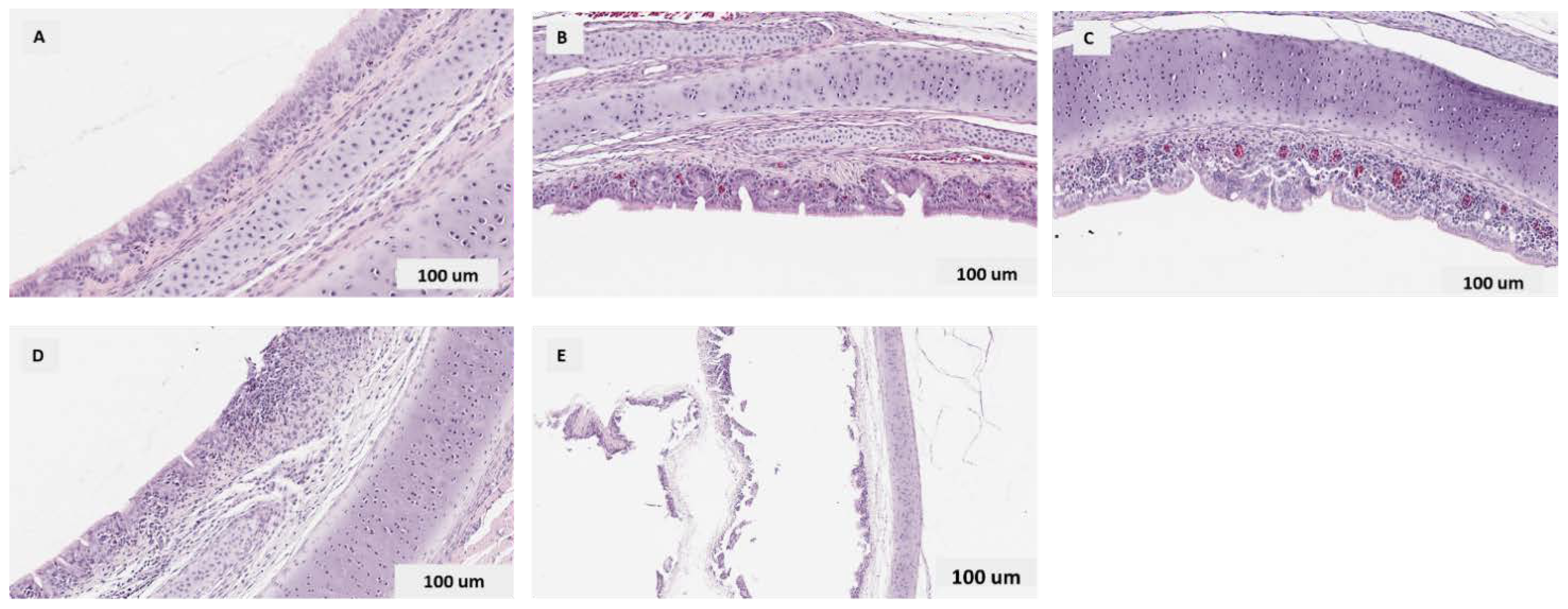
2.4.4. Histopathology and Image Analysis
2.4.5. Immunofluorescent Assay
2.5. Data and Statistical Analyses
3. Results
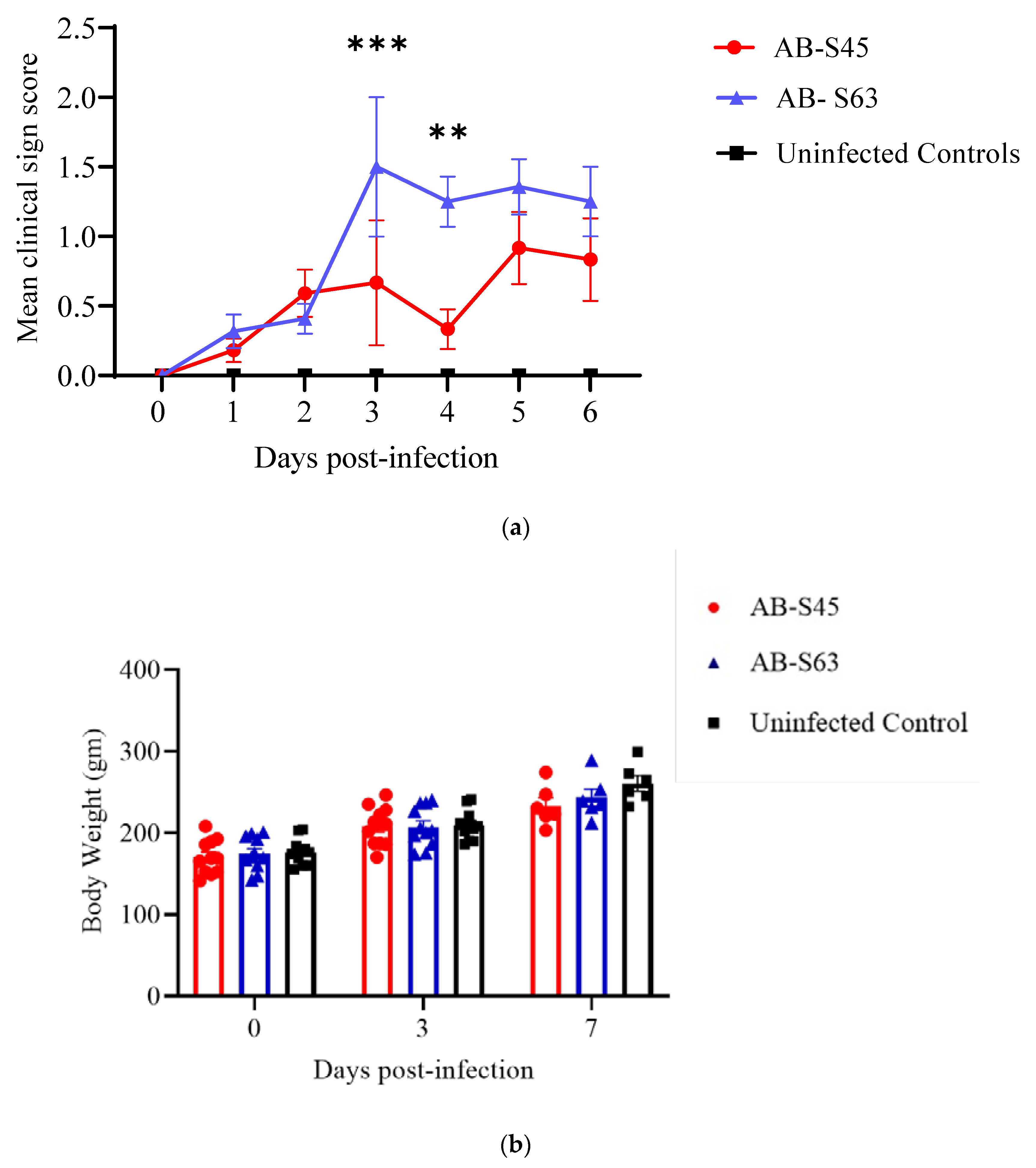
3.1. Clinical Signs
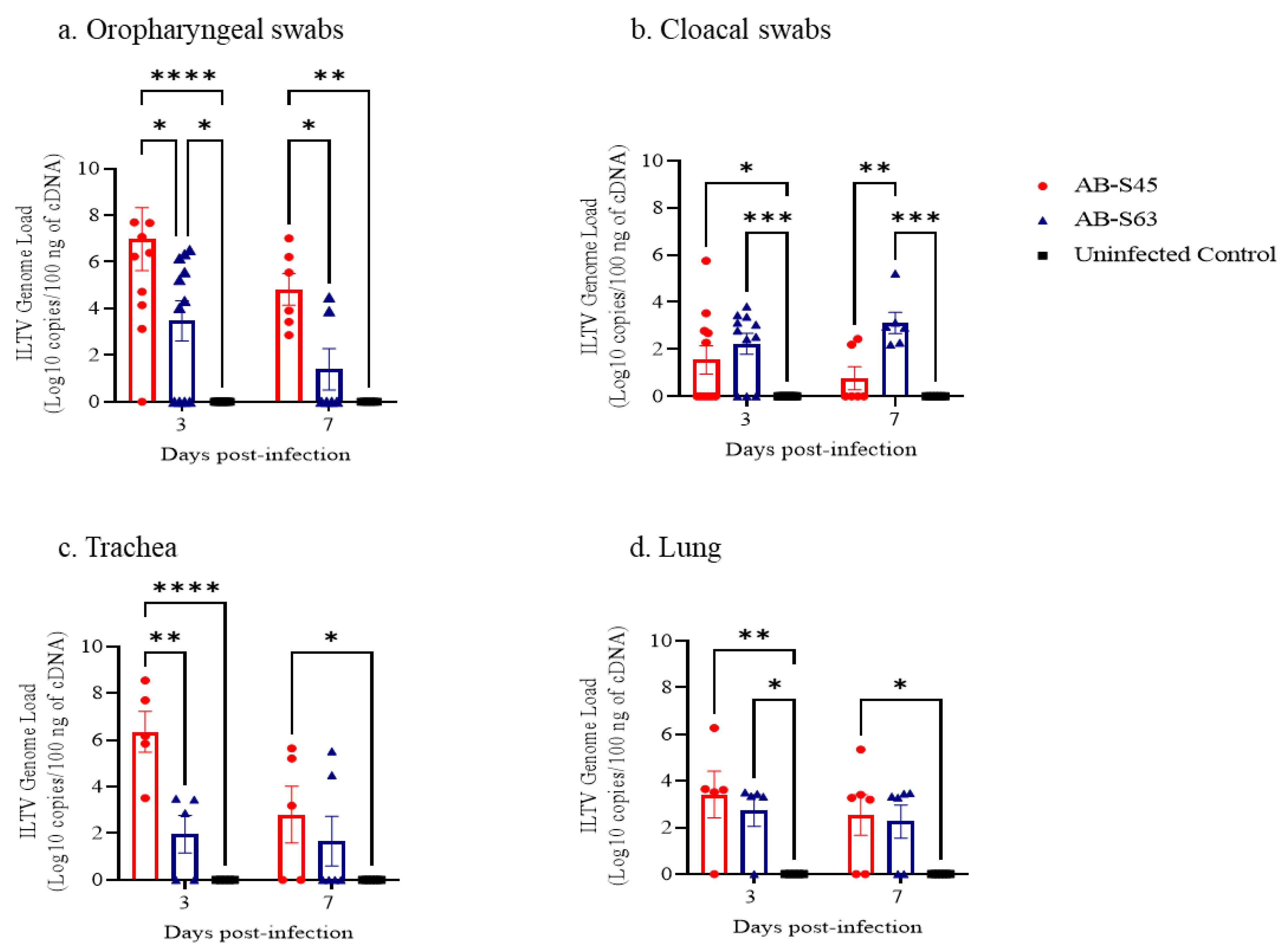
3.2. ILTV Genome Loads
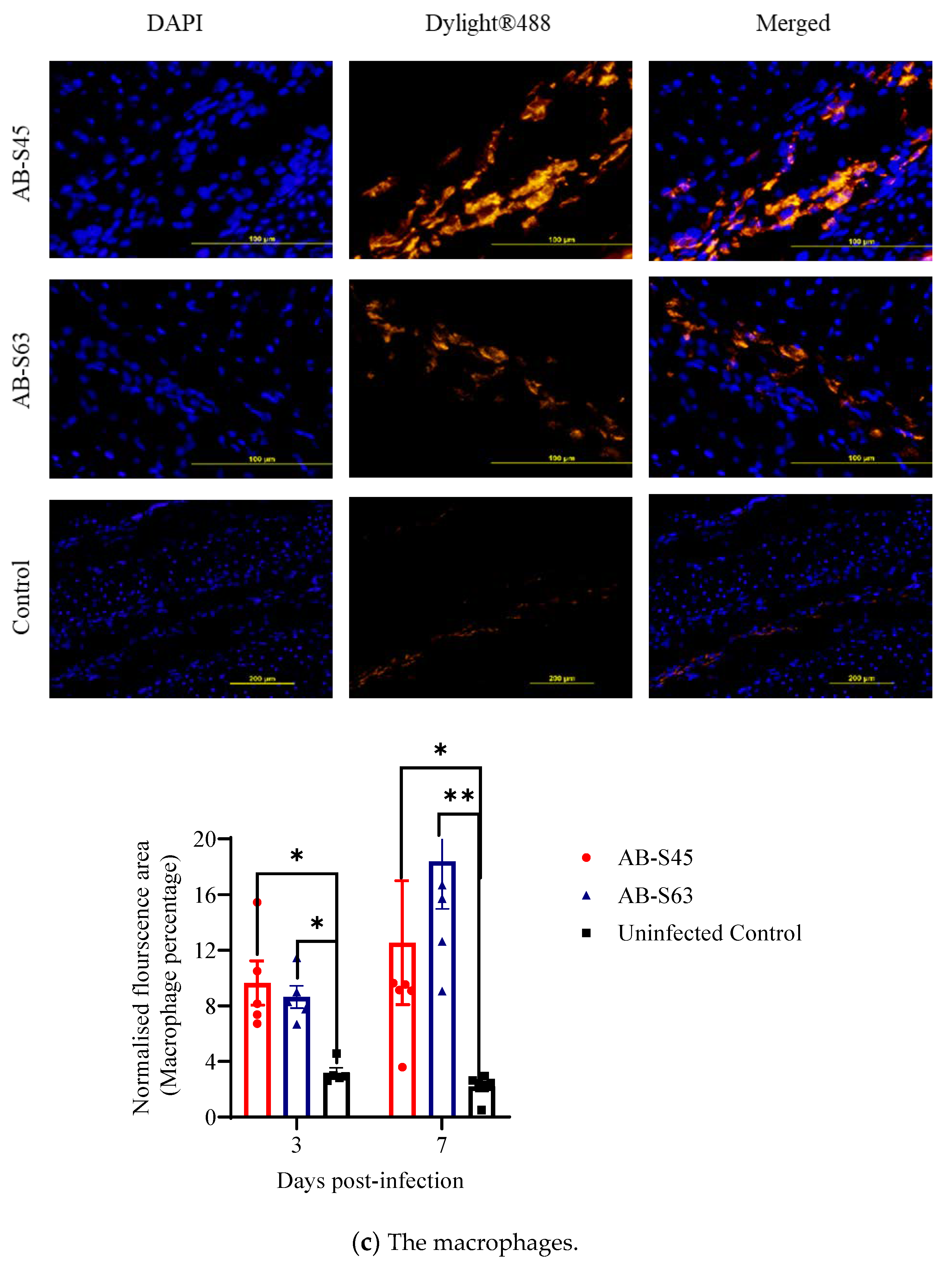
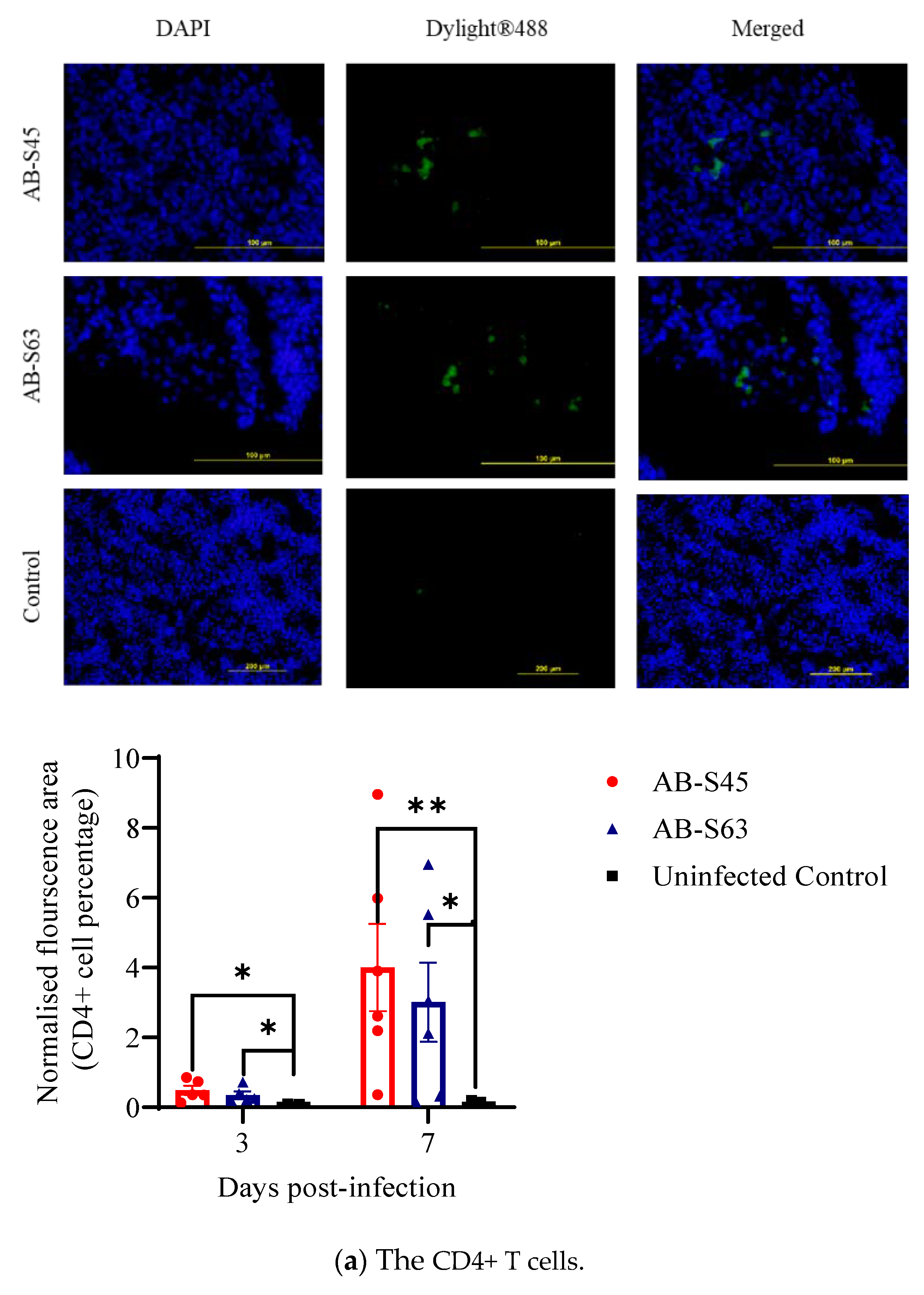
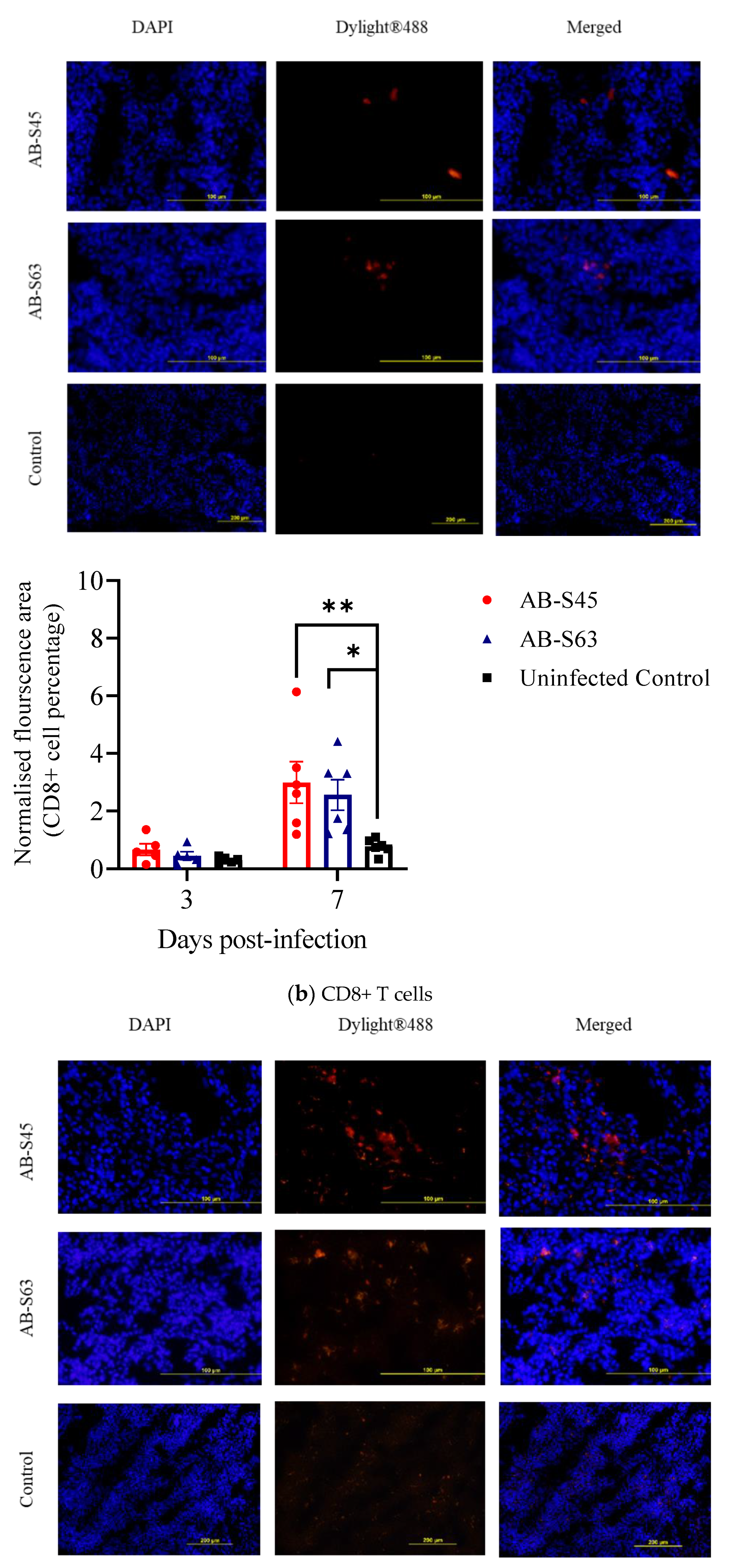
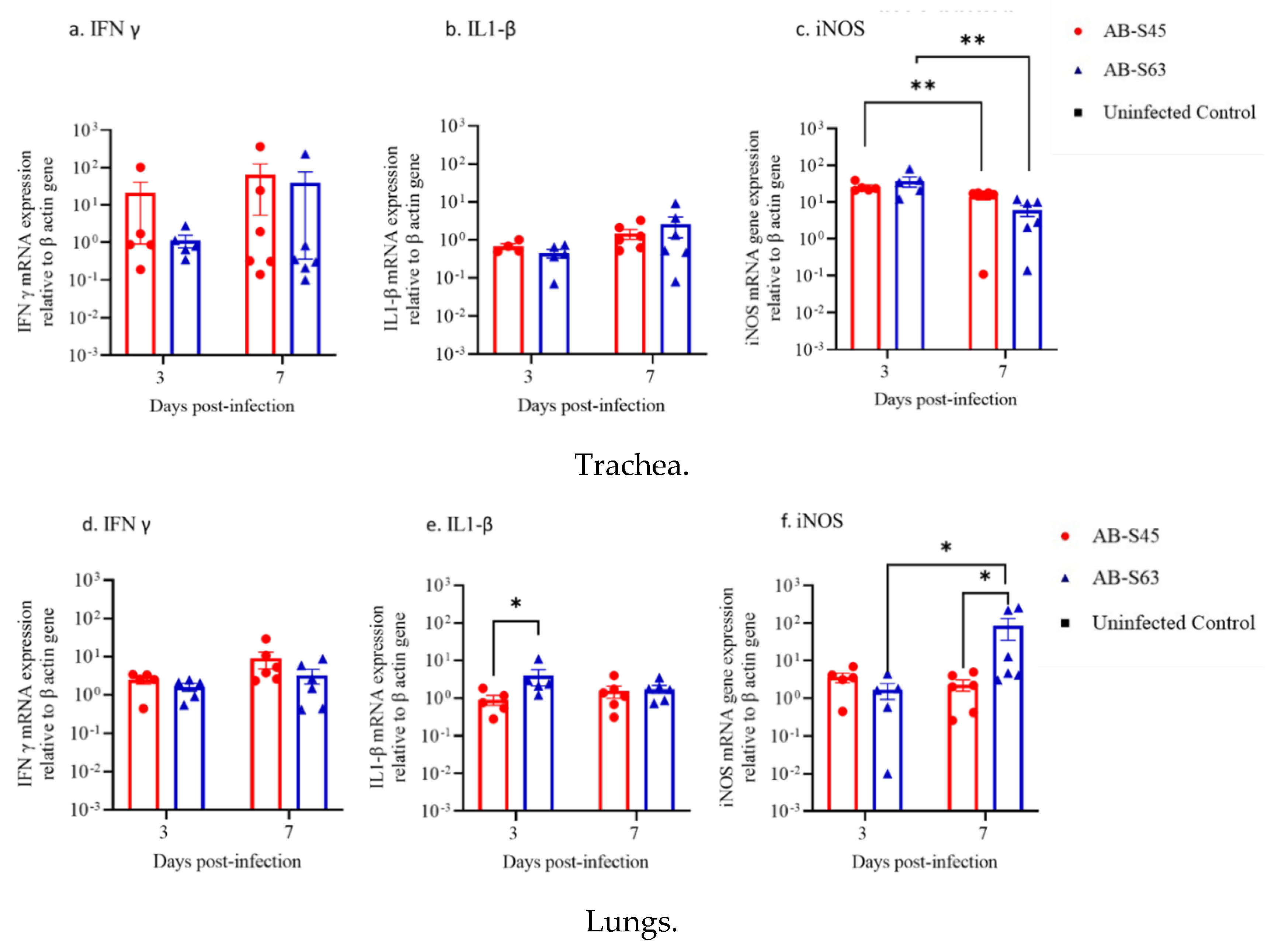
3.3. Host Responses
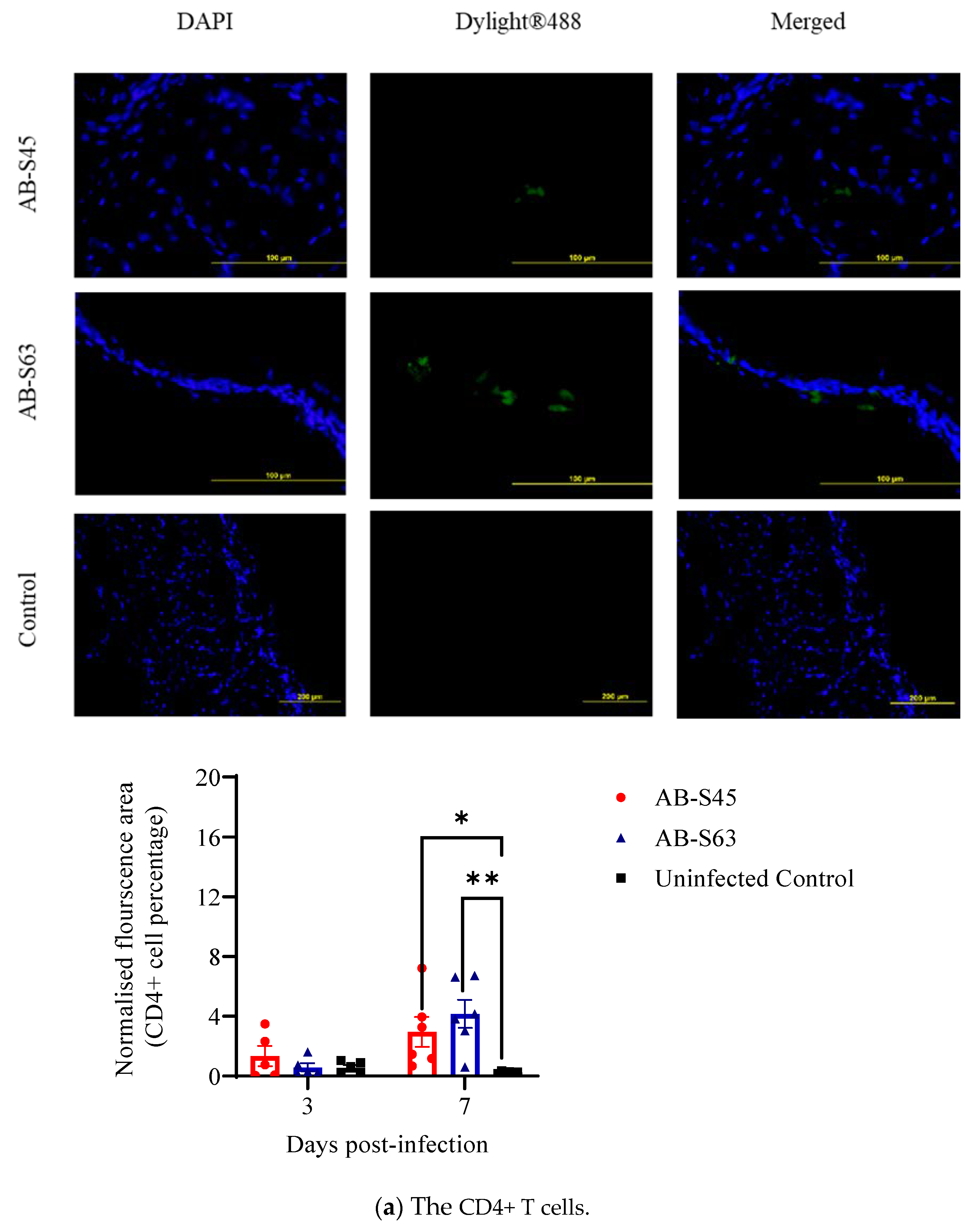
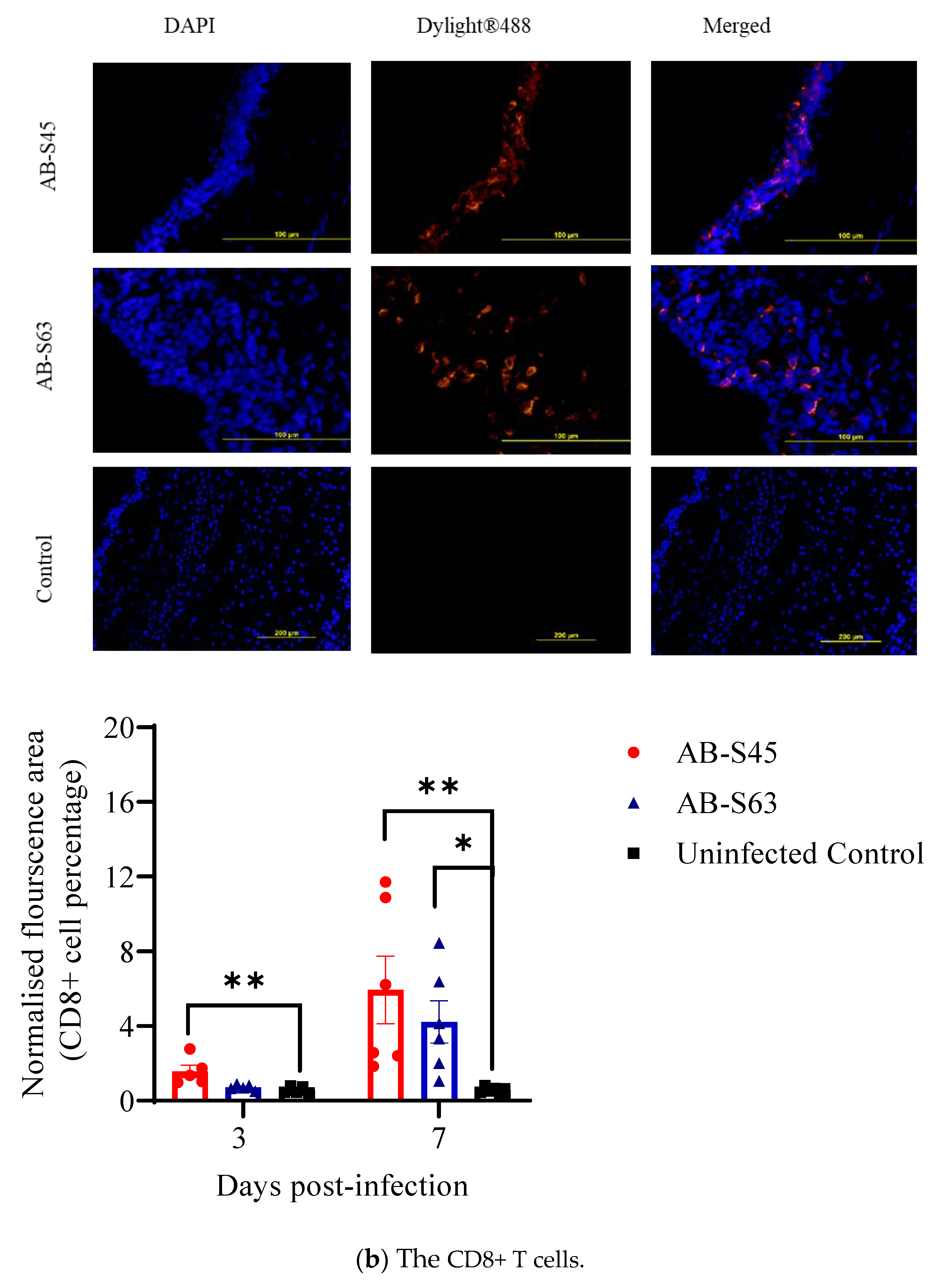
3.3.1. Recruitment of T Cells (CD4+ T Cells and CD8+ T Cells) and Macrophages
3.3.2. IFN-γ, IL-1β, and iNOS mRNA Expressions in the Trachea and Lungs
3.3.3. Histopathological Findings in the Trachea
Quantitative Histopathological Scoring of Tracheal Lesions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bagust, T.J.; Jones, R.C.; Guy, J.S. Avian infectious laryngotracheitis. Rev. Sci. Technol. 2000, 19, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Bagust, T.J. Laryngotracheitis (Gallid-1) herpesvirus infection in the chicken. 4. Latency establishment by wild and vaccine strains of ILT virus. Avian Pathol. 1986, 15, 581–595. [Google Scholar] [CrossRef] [PubMed]
- Cover, M.S. The early history of infectious laryngotracheitis. Avian Dis. 1996, 40, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Crawshaw, G.J.; Boycott, B.R. Infectious laryngotracheitis in peafowl and pheasants. Avian Dis. 1982, 26, 397–401. [Google Scholar] [CrossRef]
- Ou, S.C.; Giambrone, J.J. Infectious laryngotracheitis virus in chickens. World J. Virol. 2012, 1, 142–149. [Google Scholar] [CrossRef]
- Ojkic, D.; Swinton, J.; Vallieres, M.; Martin, E.; Shapiro, J.; Sanei, B.; Binnington, B. Characterization of field isolates of infectious laryngotracheitis virus from Ontario. Avian Pathol. 2006, 35, 286–292. [Google Scholar] [CrossRef]
- Herpesvirales. In Fenner’s Veterinary Virology; MacLachlan, N.J.; Dubovi, E.J. (Eds.) Academic Press: San Diego, CA, USA, 2011; Volume 4, pp. 179–201. [Google Scholar] [CrossRef]
- Garcia, M.; Spatz, S.J.; Guy, J.S. Infectious Laryngotracheitis; Blackwell Publishing: Ames, IA, USA, 2013. [Google Scholar]
- Guy, J.S.; Barnes, H.J.; Morgan, L.M. Virulence of infectious laryngotracheitis viruses: Comparison of modified-live vaccine viruses and North Carolina field isolates. Avian Dis. 1990, 34, 106–113. [Google Scholar] [CrossRef]
- Hayashi, S.; Odagiri, Y.; Kotani, T.; Horiuchi, T. Pathological changes of tracheal mucosa in chickens infected with infectious laryngotracheitis virus. Avian Dis. 1985, 29, 943–950. [Google Scholar] [CrossRef]
- Purcell, D.A. The ultrastructural changes produced by infectious laryngotracheitis virus in tracheal epithelium of the fowl. Res. Vet. Sci. 1971, 12, 455–458. [Google Scholar] [CrossRef]
- Kirkpatrick, N.C.; Mahmoudian, A.; Colson, C.A.; Devlin, J.M.; Noormohammadi, A.H. Relationship between mortality, clinical signs and tracheal pathology in infectious laryngotracheitis. Avian Pathol. 2006, 35, 449–453. [Google Scholar] [CrossRef] [Green Version]
- Oldoni, I.; Rodríguez-Avila, A.; Riblet, S.M.; Zavala, G.; García, M. Pathogenicity and growth characteristics of selected infectious laryngotracheitis virus strains from the United States. Avian Pathol. 2009, 38, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Bagust, T.J.; Johnson, M.A. Avian infectious laryngotracheitis: Virus-host interactions in relation to prospects for eradication. Avian Pathol. 1995, 24, 373–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, H. Infectious laryngotracheitis: A review. Braz. J. Poult. Sci. 2003, 5, 157–168. [Google Scholar] [CrossRef]
- Johnson, Y.J.; Colby, M.M.; Tablante, N.L.; Hegngi, F.N.; Salem, M.; Gedamu, N.; Pope, C. Application of Commercial and Backyard Poultry Geographic Information System Databases for the Identification of Risk Factors for Clinical Infectious Laryngotracheitis in a Cluster of Cases on the Delmarva Peninsula. Int. J. Poult. Sci. 2004, 3, 201–205. [Google Scholar]
- Vagnozzi, A.; Zavala, G.; Riblet, S.M.; Mundt, A.; García, M. Protection induced by commercially available live-attenuated and recombinant viral vector vaccines against infectious laryngotracheitis virus in broiler chickens. Avian Pathol. 2012, 41, 21–31. [Google Scholar] [CrossRef]
- Barboza-Solis, C.; Najimudeen, S.M.; Perez-Contreras, A.; Ali, A.; Joseph, T.; King, R.; Ravi, M.; Peters, D.; Fonseca, K.; Gagnon, C.A.; et al. Evaluation of Recombinant Herpesvirus of Turkey Laryngotracheitis (rHVT-LT) Vaccine against Genotype VI Canadian Wild-Type Infectious Laryngotracheitis Virus (ILTV) Infection. Vaccines 2021, 9, 1425. [Google Scholar] [CrossRef]
- García, M. Current and future vaccines and vaccination strategies against infectious laryngotracheitis (ILT) respiratory disease of poultry. Vet. Microbiol. 2017, 206, 157–162. [Google Scholar] [CrossRef]
- Johnson, D.I.; Vagnozzi, A.; Dorea, F.; Riblet, S.M.; Mundt, A.; Zavala, G.; García, M. Protection against infectious laryngotracheitis by in ovo vaccination with commercially available viral vector recombinant vaccines. Avian Dis. 2010, 54, 1251–1259. [Google Scholar] [CrossRef]
- Palomino-Tapia, V.A.; Zavala, G.; Cheng, S.; García, M. Long-term protection against a virulent field isolate of infectious laryngotracheitis virus induced by inactivated, recombinant, and modified live virus vaccines in commercial layers. Avian Pathol. 2019, 48, 209–220. [Google Scholar] [CrossRef]
- Dufour-Zavala, L. Epizootiology of infectious laryngotracheitis and presentation of an industry control program. Avian Dis. 2008, 52, 1–7. [Google Scholar] [CrossRef]
- Agnew-Crumpton, R.; Vaz, P.K.; Devlin, J.M.; O’Rourke, D.; Blacker-Smith, H.P.; Konsak-Ilievski, B.; Hartley, C.A.; Noormohammadi, A.H. Spread of the newly emerging infectious laryngotracheitis viruses in Australia. Infect. Genet. Evol. 2016, 43, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Markham, P.F.; Coppo, M.J.; Legione, A.R.; Markham, J.F.; Noormohammadi, A.H.; Browning, G.F.; Ficorilli, N.; Hartley, C.A.; Devlin, J.M. Attenuated vaccines can recombine to form virulent field viruses. Science 2012, 337, 188. [Google Scholar] [CrossRef] [PubMed]
- Perez Contreras, A.; van der Meer, F.; Checkley, S.; Joseph, T.; King, R.; Ravi, M.; Peters, D.; Fonseca, K.; Gagnon, C.A.; Provost, C.; et al. Analysis of Whole-Genome Sequences of Infectious laryngotracheitis Virus Isolates from Poultry Flocks in Canada: Evidence of Recombination. Viruses 2020, 12, 1302. [Google Scholar] [CrossRef] [PubMed]
- Bayoumi, M.; El-Saied, M.; Amer, H.; Bastami, M.; Sakr, E.E.; El-Mahdy, M. Molecular characterization and genetic diversity of the infectious laryngotracheitis virus strains circulating in Egypt during the outbreaks of 2018 and 2019. Arch. Virol. 2020, 165, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Oldoni, I.; García, M. Characterization of infectious laryngotracheitis virus isolates from the US by polymerase chain reaction and restriction fragment length polymorphism of multiple genome regions. Avian Pathol. 2007, 36, 167–176. [Google Scholar] [CrossRef] [Green Version]
- York, J.J.; Fahey, K.J. Humoral and cell-mediated immune responses to the glycoproteins of infectious laryngotracheitis herpesvirus. Arch. Virol. 1990, 115, 289–297. [Google Scholar] [CrossRef]
- Fahey, K.J.; Bagust, T.J.; York, J.J. Laryngotracheitis herpesvirus infection in the chicken: The role of humoral antibody in immunity to a graded challenge infection. Avian Pathol. 1983, 12, 505–514. [Google Scholar] [CrossRef]
- Honda, T.; Okamura, H.; Taneno, A.; Yamada, S.; Takahashi, E. The role of cell-mediated immunity in chickens inoculated with the cell-associated vaccine of attenuated infectious laryngotracheitis virus. J. Vet. Med. Sci. 1994, 56, 1051–1055. [Google Scholar] [CrossRef] [Green Version]
- Fahey, K.J.; York, J.J.; Bagust, T.J. Laryngotracheitis herpesvirus infection in the chicken. II. The adoptive transfer of resistance with immune spleen cells. Avian Pathol. 1984, 13, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Vagnozzi, A.E.; Beltrán, G.; Zavala, G.; Read, L.; Sharif, S.; García, M. Cytokine gene transcription in the trachea, Harderian gland, and trigeminal ganglia of chickens inoculated with virulent infectious laryngotracheitis virus (ILTV) strain. Avian Pathol. 2018, 47, 497–508. [Google Scholar] [CrossRef] [Green Version]
- Barboza-Solis, C.; Contreras, A.P.; Palomino-Tapia, V.A.; Joseph, T.; King, R.; Ravi, M.; Peters, D.; Fonseca, K.; Gagnon, C.A.; van der Meer, F.; et al. Genotyping of Infectious Laryngotracheitis Virus (ILTV) Isolates from Western Canadian Provinces of Alberta and British Columbia Based on Partial Open Reading Frame (ORF) a and b. Animals 2020, 10, 1634. [Google Scholar] [CrossRef] [PubMed]
- Perez-Contreras, A.; Barboza-Solis, C.; Najimudeen, S.M.; Checkley, S.; Meer, F.V.; Joseph, T.; King, R.; Ravi, M.; Peters, D.; Fonseca, K.; et al. Pathogenic and Transmission Potential of Wildtype and Chicken Embryo Origin (CEO) Vaccine Revertant Infectious Laryngotracheitis Virus. Viruses 2021, 13, 541. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.A.; Halami, M.Y.; Sultan, H.H.; Abd El-Razik, A.G.; Vahlenkamp, T.W. Chicken embryo origin-like strains are responsible for Infectious laryngotracheitis virus outbreaks in Egyptian cross-bred broiler chickens. Virus Genes 2013, 46, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- De Silva Senapathi, U.; Abdul-Cader, M.S.; Amarasinghe, A.; van Marle, G.; Czub, M.; Gomis, S.; Abdul-Careem, M.F. The In Ovo Delivery of CpG Oligonucleotides Protects against Infectious Bronchitis with the Recruitment of Immune Cells into the Respiratory Tract of Chickens. Viruses 2018, 10, 635. [Google Scholar] [CrossRef] [Green Version]
- Kameka, A.M.; Haddadi, S.; Kim, D.S.; Cork, S.C.; Abdul-Careem, M.F. Induction of innate immune response following infectious bronchitis corona virus infection in the respiratory tract of chickens. Virology 2014, 450–451, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Najimudeen, S.M.; Barboza-Solis, C.; Ali, A.; Buharideen, S.M.; Isham, I.M.; Hassan, M.S.H.; Ojkic, D.; Van Marle, G.; Cork, S.C.; van der Meer, F.; et al. Pathogenesis and host responses in lungs and kidneys following Canadian 4/91 infectious bronchitis virus (IBV) infection in chickens. Virology 2022, 566, 75–88. [Google Scholar] [CrossRef]
- Thapa, S.; Nagy, E.; Abdul-Careem, M.F. In ovo delivery of Toll-like receptor 2 ligand, lipoteichoic acid induces pro-inflammatory mediators reducing post-hatch infectious laryngotracheitis virus infection. Vet. Immunol. Immunopathol. 2015, 164, 170–178. [Google Scholar] [CrossRef]
- Wilson, F.D.; Banda, A.; Hoerr, F.J.; Alvarado, I.; Orozco, E.; Mackey, R. Histopathologic Lesion Scoring and Histomorphometric Methods for Measuring Vaccine Reactions in the Trachea of Broiler Chickens. Avian Dis. 2021, 65, 18–25. [Google Scholar] [CrossRef]
- Najimudeen, S.M.; Hassan, M.S.H.; Goldsmith, D.; Ojkic, D.; Cork, S.C.; Boulianne, M.; Abdul-Careem, M.F. Molecular Characterization of 4/91 Infectious Bronchitis Virus Leading to Studies of Pathogenesis and Host Responses in Laying Hens. Pathogens 2021, 10, 624. [Google Scholar] [CrossRef]
- Neff, C.; Sudler, C.; Hoop, R.K. Characterization of western European field isolates and vaccine strains of avian infectious laryngotracheitis virus by restriction fragment length polymorphism and sequence analysis. Avian Dis. 2008, 52, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Li, S.; Xie, Q.; Chen, F.; Bi, Y. Characterization of field strains of infectious laryngotracheitis virus in China by restriction fragment length polymorphism and sequence analysis. J. Vet. Diagn. Investig. 2016, 28, 46–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chacón, J.L.; Núñez, L.F.; Vejarano, M.P.; Parra, S.H.; Astolfi-Ferreira, C.S.; Ferreira, A.J. Persistence and spreading of field and vaccine strains of infectious laryngotracheitis virus (ILTV) in vaccinated and unvaccinated geographic regions, in Brazil. Trop. Anim. Health Prod. 2015, 47, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- García, M.; Zavala, G. Commercial Vaccines and Vaccination Strategies Against Infectious Laryngotracheitis: What We Have Learned and Knowledge Gaps That Remain. Avian Dis. 2019, 63, 325–334. [Google Scholar] [CrossRef]
- Coppo, M.J.; Hartley, C.A.; Devlin, J.M. Immune responses to infectious laryngotracheitis virus. Dev. Comp. Immunol. 2013, 41, 454–462. [Google Scholar] [CrossRef]
- Guy, J.S.; Barnes, H.J.; Smith, L. Increased virulence of modified-live infectious laryngotracheitis vaccine virus following bird-to-bird passage. Avian Dis. 1991, 35, 348–355. [Google Scholar] [CrossRef]
- Maekawa, D.; Whang, P.; Riblet, S.M.; Hurley, D.J.; Guy, J.S.; García, M. Assessing the infiltration of immune cells in the upper trachea mucosa after infectious laryngotracheitis virus (ILTV) vaccination and challenge. Avian Pathol. 2021, 50, 540–556. [Google Scholar] [CrossRef]
- Luo, J.; Carrillo, J.A.; Menendez, K.R.; Tablante, N.L.; Song, J. Transcriptome analysis reveals an activation of major histocompatibility complex 1 and 2 pathways in chicken trachea immunized with infectious laryngotracheitis virus vaccine. Poult. Sci. 2014, 93, 848–855. [Google Scholar] [CrossRef]
- Haq, K.; Wootton, S.K.; Barjesteh, N.; Golovan, S.; Bendall, A.; Sharif, S. Effects of interferon-γ knockdown on vaccine-induced immunity against Marek’s disease in chickens. Can. J. Vet. Res. 2015, 79, 1–7. [Google Scholar]
- Qureshi, M.A. Avian macrophage and immune response: An overview. Poult. Sci. 2003, 82, 691–698. [Google Scholar] [CrossRef]
- Wu, Z.; Kaiser, P. Antigen presenting cells in a non-mammalian model system, the chicken. Immunobiology 2011, 216, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Amarasinghe, A.; Abdul-Cader, M.S.; Nazir, S.; De Silva Senapathi, U.; van der Meer, F.; Cork, S.C.; Gomis, S.; Abdul-Careem, M.F. Infectious bronchitis corona virus establishes productive infection in avian macrophages interfering with selected antimicrobial functions. PLoS ONE 2017, 12, e0181801. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Cader, M.S.; Ehiremen, G.; Nagy, E.; Abdul-Careem, M.F. Low pathogenic avian influenza virus infection increases the staining intensity of KUL01+ cells including macrophages yet decrease of the staining intensity of KUL01+ cells using clodronate liposomes did not affect the viral genome loads in chickens. Vet. Immunol. Immunopathol. 2018, 198, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Song, J.J.; Wooming, A.; Li, X.; Zhou, H.; Bottje, W.G.; Kong, B.-W. Transcriptional profiling of host gene expression in chicken embryo lung cells infected with laryngotracheitis virus. BMC Genom. 2010, 11, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Wang, Z.; Shao, C.; Yu, J.; Liu, H.; Chen, H.; Li, L.; Wang, X.; Ren, Y.; Huang, X.; et al. Analysis of chicken macrophage functions and gene expressions following infectious bronchitis virus M41 infection. Vet. Res. 2021, 52, 14. [Google Scholar] [CrossRef] [PubMed]
- Haddadi, S.; Thapa, S.; Kameka, A.; Hui, J.; Czub, M.; Nagy, E.; Muench, G.; Abdul-Careem, M. Toll-like receptor 2 ligand, lipoteichoic acid is inhibitory against infectious laryngotracheitis virus infection in vitro and in vivo. Dev. Comp. Immunol. 2015, 48, 22–32. [Google Scholar] [CrossRef]
- Thapa, S.; Cader, M.S.A.; Murugananthan, K.; Nagy, E.; Sharif, S.; Czub, M.; Abdul-Careem, M.F. In ovo delivery of CpG DNA reduces avian infectious laryngotracheitis virus induced mortality and morbidity. Viruses 2015, 7, 1832–1852. [Google Scholar] [CrossRef]
- Barjesteh, N.; Behboudi, S.; Brisbin, J.T.; Villanueva, A.I.; Nagy, E.; Sharif, S. TLR ligands induce antiviral responses in chicken macrophages. PLoS ONE 2014, 9, e105713. [Google Scholar] [CrossRef]
- Burggraaf, S.; Bingham, J.; Payne, J.; Kimpton, W.G.; Lowenthal, J.W.; Bean, A.G. Increased inducible nitric oxide synthase expression in organs is associated with a higher severity of H5N1 influenza virus infection. PLoS ONE 2011, 6, e14561. [Google Scholar] [CrossRef] [Green Version]
- Von Kortzfleisch, V.T.; Karp, N.A.; Palme, R.; Kaiser, S.; Sachser, N.; Richter, S.H. Improving reproducibility in animal research by splitting the study population into several ‘mini-experiments’. Sci. Rep. 2020, 10, 16579. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bird # | 3 Days Post-Infection | 7 Days Post-Infection | ||||
|---|---|---|---|---|---|---|
| Control | AB-S45 ILTV | AB-63 ILTV | Control | AB-S45 ILTV | AB-S63 ILTV | |
| 1 | 1 | 3 | 4 | 0 | 4 | 1 |
| 2 | 0 | 1 | 2 | 1 | 3 | 1 |
| 3 | 0 | 2 | 1 | 1 | 1 | 1 |
| 4 | 1 | 3 | 2 | 1 | 1 | 5 |
| 5 | 1 | 1 | 1 | 0 | 1 | 0 |
| 6 | 1 | - | - | 1 | n/a * | 3 |
| Mean ± SEM ** | 0.6 ± 0.25 | 2 ± 0.45 | 2 ± 0.55 | 0.7 ± 0.2 | 2 ± 0.7 | 1.8 ± 0.8 |
| Bird # | 3 Days Post-Infection | 7 Days Post-Infection | ||||
|---|---|---|---|---|---|---|
| Control | AB-S45 ILTV | AB-63 ILTV | Control | AB-S45 ILTV | AB-S63 ILTV | |
| 1 | 56.85 | 60.51 | 21.74 | 29.60 | 44.20 | 33.13 |
| 2 | 60.91 | 28.13 | 34.48 | 23.42 | 66.86 | 29.34 |
| 3 | 42.71 | 27.46 | 35.35 | 30.85 | 37.34 | 25.84 |
| 4 | 58.25 | 36.20 | 21.38 | 34.21 | 41.25 | 24.04 |
| 5 | 25.97 | 67.06 | 19.63 | 36.52 | 39.68 | 52.97 |
| 6 | 26.63 | - | - | 20.76 | n/a * | 48.32 |
| Mean± SEM ** | 45.2 ± 6.5 | 43.87 ± 8.3 | 26.51 ± 3.4 | 29.23 ± 2.5 | 45.87 ± 5.4 | 35.61 ± 4.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elshafiee, E.A.; Isham, I.M.; Najimudeen, S.M.; Perez-Contreras, A.; Barboza-Solis, C.; Ravi, M.; Abdul-Careem, M.F. Host Responses Following Infection with Canadian-Origin Wildtype and Vaccine Revertant Infectious Laryngotracheitis Virus. Vaccines 2022, 10, 782. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10050782
Elshafiee EA, Isham IM, Najimudeen SM, Perez-Contreras A, Barboza-Solis C, Ravi M, Abdul-Careem MF. Host Responses Following Infection with Canadian-Origin Wildtype and Vaccine Revertant Infectious Laryngotracheitis Virus. Vaccines. 2022; 10(5):782. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10050782
Chicago/Turabian StyleElshafiee, Esraa A., Ishara M. Isham, Shahnas M. Najimudeen, Ana Perez-Contreras, Catalina Barboza-Solis, Madhu Ravi, and Mohamed Faizal Abdul-Careem. 2022. "Host Responses Following Infection with Canadian-Origin Wildtype and Vaccine Revertant Infectious Laryngotracheitis Virus" Vaccines 10, no. 5: 782. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10050782