

Humoral and Cellular Immune Responses Induced by Bivalent DNA Vaccines Expressing Fusion Capsid Proteins of Porcine Circovirus Genotypes 2a and 2b

Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Sequence Retrieval and Phylogenetic Analysis
2.2. Prediction of the Immunogen’s Properties
2.3. B Cell Epitope Prediction
2.4. CD8 T Cell Epitope Prediction
2.5. Generation of DNA Vaccine
2.6. Preparation of the PCV2 Capsid Proteins
2.7. Cell Culture and Transfection
2.8. Immunofluorescence Staining
2.9. Western Blot Analysis
2.10. Mouse Immunization
2.11. Indirect Enzyme-Linked Immunosorbent Assay (ELISA)
2.12. IFN-γ Enzyme-Linked Immunosorbent Spot (IFN-γ ELISpot)
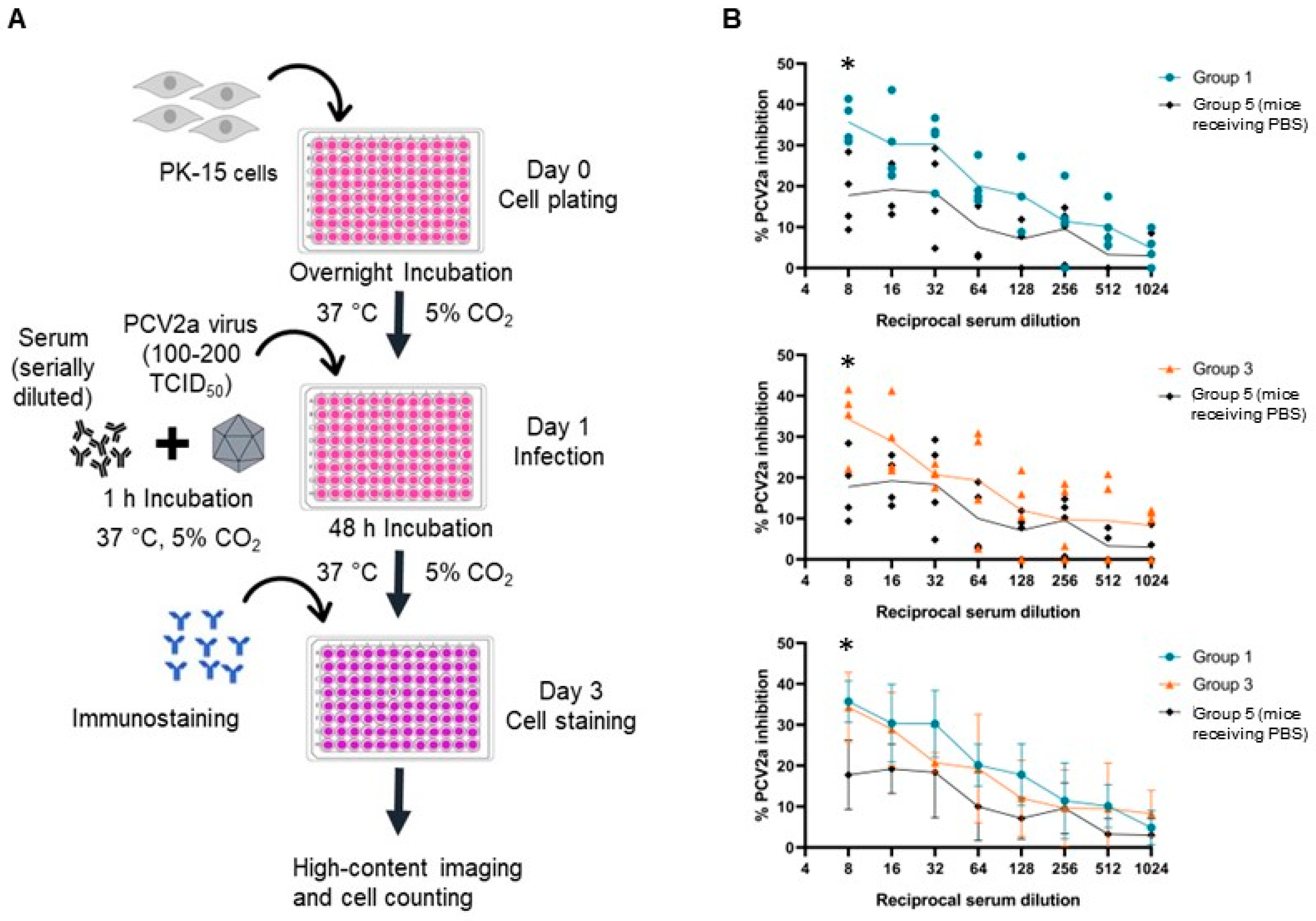
2.13. Serum Neutralization Assay
2.14. Statical Analysis
3. Results
3.1. Sequence Retrieval, Analysis and Generation of the PCV2a and PCV2b Capsid Consensus Sequences
3.2. Design and In Silico Validation of Bivalent PCV2 Immunogens
3.3. Generation of Bivalent DNA Plasmids and Protein Expression Study
3.4. B Cell and CD8 T Cell Epitope Prediction
3.5. Mouse Immunization and Total IgG Response
3.6. Th1/Th2 Immune Response Induced by the PCV2b-2a-Based DNA Vaccines
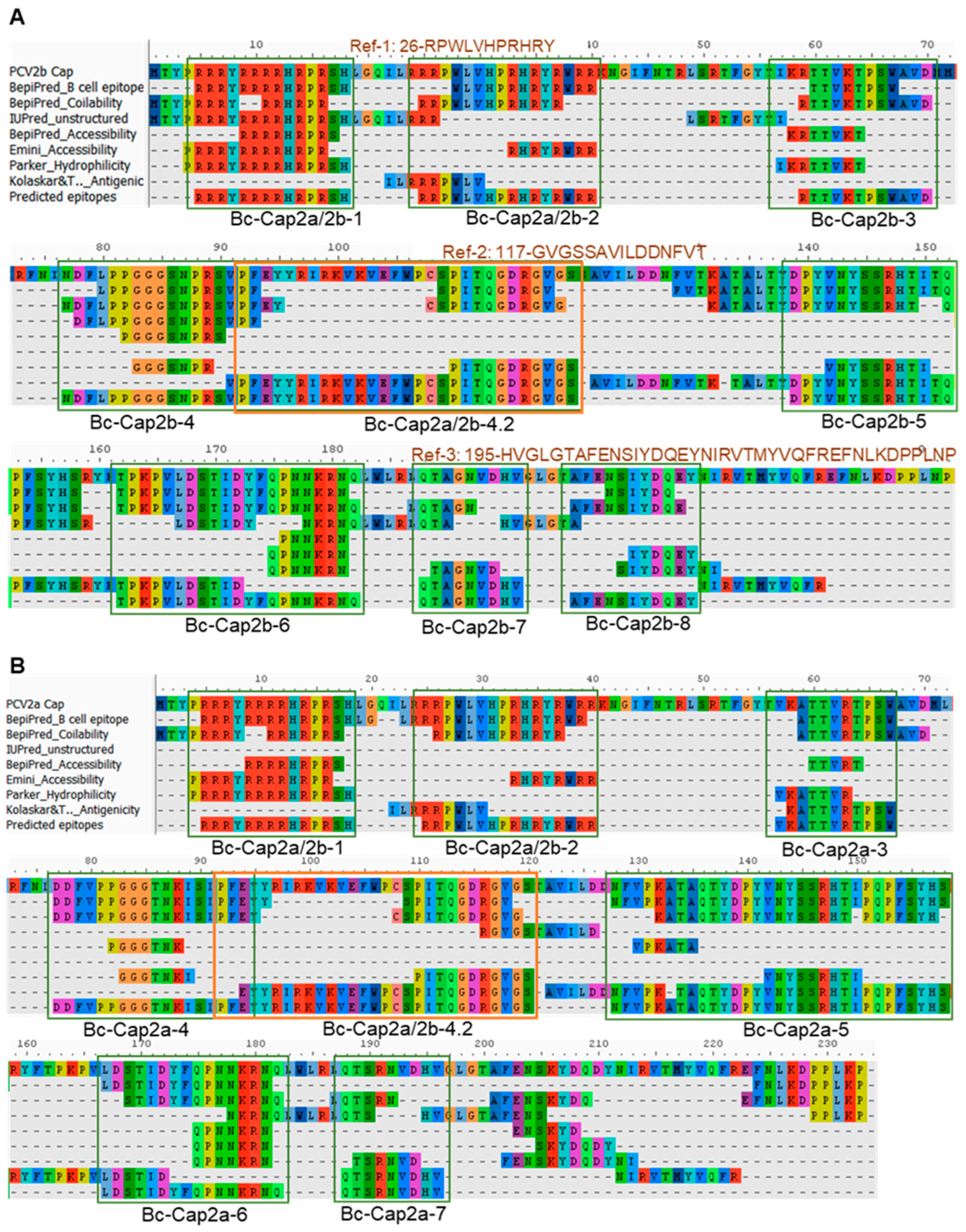
3.7. Linear B Cell Epitope Profile of Cap2a and Cap2b
3.8. Cellular Immune Response and CD8 T Cell Epitope Profile
3.9. Neutralizing Activity of the Antibodies from Immunized Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, Y.; Cai, X.; Zhan, Y.; Yuan, X.; Liu, T.; Tan, L.; Li, Y.; Zhan, L.; Yang, L.; Liu, W.; et al. Truncated Rep protein of porcine circovirus 2 (PCV2) caused by a naturally occurring mutation reduced virus replication in PK15 cells. BMC Vet. Res. 2019, 15, 248. [Google Scholar] [CrossRef]
- Chae, C. A review of porcine circovirus 2-associated syndromes and diseases. Vet. J. 2005, 169, 326–336. [Google Scholar] [CrossRef]
- Bao, F.; Mi, S.; Luo, Q.; Guo, H.; Tu, C.; Zhu, G.; Gong, W. Retrospective study of porcine circovirus type 2 infection reveals a novel genotype PCV2f. Transbound. Emerg. Dis. 2018, 65, 432–440. [Google Scholar] [CrossRef]
- Franzo, G.; Segalés, J. Porcine circovirus 2 (PCV-2) genotype update and proposal of a new genotyping methodology. PLoS ONE 2018, 13, e0208585. [Google Scholar] [CrossRef]
- Morozov, I.; Sirinarumitr, T.; Sorden, S.D.; Halbur, P.G.; Morgan, M.K.; Yoon, K.J.; Paul, P.S. Detection of a novel strain of porcine circovirus in pigs with postweaning multisystemic wasting syndrome. J. Clin. Microbiol. 1998, 36, 2535–2541. [Google Scholar] [CrossRef]
- Dupont, K.; Nielsen, E.O.; Baekbo, P.; Larsen, L.E. Genomic analysis of PCV2 isolates from Danish archives and a current PMWS case–control study supports a shift in genotypes with time. Vet. Microbiol. 2008, 128, 56–64. [Google Scholar] [CrossRef]
- Larochelle, R.; Magar, R.; D’Allaire, S. Genetic characterization and phylogenetic analysis of porcine circovirus type 2 (PCV2) strains from cases presenting various clinical conditions. Virus Res. 2002, 90, 101–112. [Google Scholar] [CrossRef]
- Segalés, J.; Olvera, A.; Grau-Roma, L.; Charreyre, C.; Nauwynck, H.; Larsen, L.; Dupont, K.; McCullough, K.; Ellis, J.; Krakowka, S.J.V.R.; et al. PCV-2 genotype definition and nomenclature. Vet. Rec. 2008, 162, 867–869. [Google Scholar] [CrossRef]
- Opriessnig, T.; Ramamoorthy, S.; Madson, D.M.; Patterson, A.R.; Pal, N.; Carman, S.; Meng, X.J.; Halbur, P.G. Differences in virulence among porcine circovirus type 2 isolates are unrelated to cluster type 2a or 2b and prior infection provides heterologous protection. J. Gen. Virol. 2008, 89, 2482–2491. [Google Scholar] [CrossRef]
- Harding, J.C.S.; Ellis, J.A.; Mclntosh, K.A.; Krakowka, S. Dual heterologous porcine circovirus genogroup 2a/2b infection induces severe disease in germ-free pigs. Vet. Microbiol. 2010, 145, 209–219. [Google Scholar] [CrossRef]
- Gagnon, C.A.; Tremblay, D.; Tijssen, P.; Venne, M.H.; Houde, A.; Elahi, S.M. The emergence of porcine circovirus 2b genotype (PCV-2b) in swine in Canada. Can. Vet. J. 2007, 48, 811–819. [Google Scholar] [PubMed]
- Zhai, S.L.; Chen, S.N.; Wei, Z.Z.; Zhang, J.W.; Huang, L.; Lin, T.; Yue, C.; Ran, D.L.; Yuan, S.S.; Wei, W.K.; et al. Co-existence of multiple strains of porcine circovirus type 2 in the same pig from China. Virol. J. 2011, 8, 517. [Google Scholar] [CrossRef]
- Fort, M.; Sibila, M.; Allepuz, A.; Mateu, E.; Roerink, F.; Segalés, J. Porcine circovirus type 2 (PCV2) vaccination of conventional pigs prevents viremia against PCV2 isolates of different genotypes and geographic origins. Vaccine 2008, 26, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Bandrick, M.; Balasch, M.; Heinz, A.; Taylor, L.; King, V.; Toepfer, J.; Foss, D. A bivalent porcine circovirus type 2 (PCV2), PCV2a-PCV2b, vaccine offers biologically superior protection compared to monovalent PCV2 vaccines. Vet. Res. 2022, 53, 12. [Google Scholar] [CrossRef]
- Mahé, D.; Blanchard, P.; Truong, C.; Arnauld, C.; Le Cann, P.; Cariolet, R.; Madec, F.; Albina, E.; Jestin, A. Differential recognition of ORF2 protein from type 1 and type 2 porcine circoviruses and identification of immunorelevant epitopes. J. Gen. Virol. 2000, 81, 1815–1824. [Google Scholar] [CrossRef]
- Truong, C.; Mahé, D.; Blanchard, P.; Le Dimna, M.; Madec, F.; Jestin, A.; Albina, E. Identification of an immunorelevant ORF2 epitope from porcine circovirus type 2 as a serological marker for experimental and natural infection. Arch. Virol. 2001, 146, 1197–1211. [Google Scholar] [CrossRef]
- Fort, M.; Fernandes, L.T.; Nofrarias, M.; Diaz, I.; Sibila, M.; Pujols, J.; Mateu, E.; Segales, J. Development of cell-mediated immunity to porcine circovirus type 2 (PCV2) in caesarean-derived, colostrum-deprived piglets. Vet. Immunol. Immunopathol. 2009, 129, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Cousins, D.J.; Staynov, D.Z.; Lee, T.H. Regulation of Interleukin-5 and Granulocyte-Macrophage Colony-Stimulating Factor Expression. Am. J. Respir. Crit. Care Med. 1994, 150 5 Pt 2, S50–S53. [Google Scholar] [CrossRef]
- Nimer, S.D.; Uchida, H. Regulation of granulocyte-macrophage colony-stimulating factor and interleukin 3 expression. Stem Cells 1995, 13, 324–335. [Google Scholar] [CrossRef]
- Disis, M.L.; Bernhard, H.; Shiota, F.M.; Hand, S.L.; Gralow, J.R.; Huseby, E.S.; Gillis, S.M.A.C.; Cheever, M.A. Granulocyte-macrophage colony-stimulating factor: An effective adjuvant for protein and peptide-based vaccines. Blood 1996, 88, 202–210. [Google Scholar] [CrossRef]
- Howard, M.; Farrar, J.O.H.N.; Hilfiker, M.A.R.Y.; Johnson, B.; Takatsu, K.; Hamaoka, T.; Paul, W.E. Identification of a T cell-derived b cell growth factor distinct from interleukin 2. J. Exp. Med. 1982, 155, 914–923. [Google Scholar] [CrossRef]
- Choi, H.J.; Gwon, Y.D.; Jang, Y.; Cho, Y.; Heo, Y.K.; Lee, H.J.; Kim, K.C.; Choi, J.; Lee, J.B.; Kim, Y.B. Effect of AcHERV-GmCSF as an influenza virus vaccine adjuvant. PLoS ONE 2015, 10, e0129761. [Google Scholar] [CrossRef]
- DaSilva, K.A.; Brown, M.E.; Westaway, D.; McLaurin, J. Immunization with amyloid-β using GM-CSF and IL-4 reduces amyloid burden and alters plaque morphology. Neurobiol. Dis. 2006, 23, 433–444. [Google Scholar] [CrossRef]
- Osorio, Y.; Ghiasi, H. Comparison of Adjuvant Efficacy of Herpes Simplex Virus Type 1 Recombinant Viruses Expressing T H 1 and T H 2 Cytokine Genes. J. Virol. 2003, 77, 5774–5783. [Google Scholar] [CrossRef] [PubMed]
- Charlton, B.; Lafferty, K.J. The Th1/Th2 balance in autoimmunity. Curr. Opin. Immunol. 1995, 7, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Leitner, W.W.; Seguin, M.C.; Ballou, W.R.; Seitz, J.P.; Schultz, A.M.; Sheehy, M.J.; Lyon, J.A. Immune responses induced by intramuscular or gene gun injection of protective deoxyribonucleic acid vaccines that express the circumsporozoite protein from Plasmodium berghei malaria parasites. J. Immunol. (Baltim. Md. 1950) 1997, 159, 6112–6119. [Google Scholar] [CrossRef]
- Li, Z.; Howard, A.; Kelley, C.; Delogu, G.; Collins, F.; Morris, S. Immunogenicity of DNA vaccines expressing tuberculosis proteins fused to tissue plasminogen activator signal sequences. Infect. Immun. 1999, 67, 4780–4786. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.K.; Lager, K.M.; Kohutyuk, O.I.; Vincent, A.L.; Henry, S.C.; Baker, R.B.; Rowland, R.R.; Dunham, A.G. Detection of two porcine circovirus type 2 genotypic groups in United States swine herds. Arch. Virol. 2007, 152, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Polyiam, K.; Ruengjitchatchawalya, M.; Mekvichitsaeng, P.; Kaeoket, K.; Hoonsuwan, T.; Roshorm, Y.M. Immunodominant and Neutralizing Linear B-Cell Epitopes Spanning the Spike and Membrane Proteins of Porcine Epidemic Diarrhea Virus. Front. Immunol. 2022, 12, 785293. [Google Scholar] [CrossRef] [PubMed]
- Polyiam, K.; Phoolcharoen, W.; Butkhot, N.; Srisaowakarn, C.; Thitithanyanont, A.; Auewarakul, P.; Hoonsuwan, T.; Ruengjitchatchawalya, M.; Mekvichitsaeng, P.; Roshorm, Y.M. Immunodominant linear B cell epitopes in the spike and membrane proteins of SARS-CoV-2 identified by immunoinformatics prediction and immunoassay. Sci. Rep. 2021, 11, 20383. [Google Scholar] [CrossRef]
- Solis Worsfold, C.; Dardari, R.; Law, S.; Eschbaumer, M.; Nourozieh, N.; Marshall, F.; Czub, M. Assessment of neutralizing and non-neutralizing antibody responses against Porcine circovirus 2 in vaccinated and non-vaccinated farmed pigs. J. Gen. Virol. 2015, 96, 2743–2748. [Google Scholar] [CrossRef]
- Harakuni, T.; Sugawa, H.; Komesu, A.; Tadano, M.; Arakawa, T. Heteropentameric cholera toxin B subunit chimeric molecules genetically fused to a vaccine antigen induce systemic and mucosal immune responses: A potential new strategy to target recombinant vaccine antigens to mucosal immune systems. Infect. Immun. 2005, 73, 5654–5665. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.E.; Aquino, G.; Ostoa-Saloma, P.; Laclette, J.P.; Rocha-Zavaleta, L. Cholera toxin B-subunit gene enhances mucosal immunoglobulin A, Th1-type, and CD8+ cytotoxic responses when coadministered intradermally with a DNA vaccine. Clin. Vaccine Immunol. 2004, 11, 711–719. [Google Scholar] [CrossRef]
- Yang, S.; Cohen, C.J.; Peng, P.D.; Zhao, Y.; Cassard, L.; Yu, Z.; Zheng, Z.; Jones, S.; Restifo, N.P.; Rosenberg, S.A.; et al. Development of optimal bicistronic lentiviral vectors facilitates high-level TCR gene expression and robust tumor cell recognition. Gene Ther. 2008, 15, 1411–1423. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Qian, J.J.; Yi, S.; Harding, T.C.; Tu, G.H.; VanRoey, M.; Jooss, K. Stable antibody expression at therapeutic levels using the 2A peptide. Nat. Biotechnol. 2005, 23, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Meas, S.; Mekvichitsaeng, P.; Roshorm, Y.M. Co-expression of self-cleaved multiple proteins derived from Porcine Reproductive and Respiratory Syndrome Virus by bicistronic and tricistronic DNA vaccines. Protein Expr. Purif. 2021, 177, 105763. [Google Scholar] [CrossRef]
- Guo, L.; Lu, Y.; Huang, L.; Wei, Y.; Liu, C. Identification of a new antigen epitope in the nuclear localization signal region of porcine circovirus type 2 capsid protein. Intervirology 2011, 54, 156–163. [Google Scholar] [CrossRef]
- Hung, L.C.; Yang, C.Y.; Cheng, I.C. Peptides mimicking viral proteins of porcine circovirus type 2 were profiled by the spectrum of mouse anti-PCV2 antibodies. BMC Immunol. 2017, 18, 25. [Google Scholar] [CrossRef]
- Finkelman, F.D.; Holmes, J.; Katona, I.M.; Urban, J.F., Jr.; Beckmann, M.P.; Park, L.S.; Schooley, K.A.; Coffman, R.L.; Mosmann, T.R.; Paul, W.E. Lymphokine control of in vivo immunoglobulin isotype selection. Annu. Rev. Immunol. 1990, 8, 303–333. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tikoo, S.K.; Babiuk, L.A. Nuclear localization of the ORF2 protein encoded by porcine circovirus type 2. Virology 2001, 285, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Xiao, S.; Tong, T.; Wang, S.; Xie, L.; Jiang, Y.; Chen, H.; Fang, L. Immunogenicity of porcine circovirus type 2 capsid protein targeting to different subcellular compartments. Mol. Immunol. 2008, 45, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Becker, Y. Immunological and regulatory functions of uninfected and virus infected immature and mature subtypes of dendritic cells—A review. Virus Genes 2003, 26, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Nelms, K.; Keegan, A.D.; Zamorano, J.; Ryan, J.J.; Paul, W.E. The IL-4 receptor: Signaling mechanisms and biologic functions. Annu. Rev. Immunol. 1999, 17, 701–738. [Google Scholar] [CrossRef]
- Mao, Q.; Zhang, W.; Ma, S.; Qiu, Z.; Li, B.; Xu, C.; He, H.; Fan, S.; Wu, K.; Chen, J.; et al. Fusion expression and immune effect of PCV2 cap protein tandem multiantigen epitopes with CD154/GM-CSF. Vet. Sci. 2021, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Bagarazzi, M.L.; Yan, J.; Morrow, M.P.; Shen, X.; Parker, R.L.; Lee, J.C.; Giffear, M.; Pankhong, P.; Khan, A.S.; Broderick, K.E.; et al. Immunotherapy against HPV16/18 generates potent TH1 and cytotoxic cellular immune responses. Sci. Transl. Med. 2012, 4, 155ra138. [Google Scholar] [CrossRef] [PubMed]
- Safar, H.A.; Mustafa, A.S.; Amoudy, H.A.; El-Hashim, A. The effect of adjuvants and delivery systems on Th1, Th2, Th17 and Treg cytokine responses in mice immunized with Mycobacterium tuberculosis-specific proteins. PLoS ONE 2020, 15, e0228381. [Google Scholar] [CrossRef]
- Zhang, H.; Qian, P.; Liu, L.; Qian, S.; Chen, H.; Li, X. Virus-like particles of chimeric recombinant porcine circovirus type 2 as antigen vehicle carrying foreign epitopes. Viruses 2014, 6, 4839–4855. [Google Scholar] [CrossRef]
- Shen, H.G.; Zhou, J.Y.; Huang, Z.Y.; Guo, J.Q.; Xing, G.; He, J.L.; Yan, Y.; Gong, L.Y. Protective immunity against porcine circovirus 2 by vaccination with ORF2-based DNA and subunit vaccines in mice. J. Gen. Virol. 2008, 89, 1857–1865. [Google Scholar] [CrossRef]
- Shang, S.B.; Jin, Y.L.; Jiang, X.T.; Zhou, J.Y.; Zhang, X.; Xing, G.; He, J.; Yan, Y. Fine mapping of antigenic epitopes on capsid proteins of porcine circovirus, and antigenic phenotype of porcine circovirus type 2. Mol. Immunol. 2009, 46, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.; Yan, A.; Luo, W.; Hu, Y.F.; Li, R.C.; Jiang, D.L.; Yu, X.L. Epitope screening of the PCV2 Cap protein by use of a random peptide-displayed library and polyclonal antibody. Virus Res. 2013, 177, 103–107. [Google Scholar] [CrossRef]
- Khayat, R.; Brunn, N.; Speir, J.A.; Hardham, J.M.; Ankenbauer, R.G.; Schneemann, A.; Johnson, J.E. The 2.3-angstrom structure of porcine circovirus 2. J. Virol. 2011, 85, 7856–7862. [Google Scholar] [CrossRef]
- Qin, F.; Xia, F.; Chen, H.; Cui, B.; Feng, Y.; Zhang, P.; Chen, J.; Luo, M. A guide to nucleic acid vaccines in the prevention and treatment of infectious diseases and cancers: From basic principles to current applications. Front. Cell Dev. Biol. 2021, 9, 633776. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, L.S.; Gilpin, D.F.; Douglas, A.; McNeilly, F.; McNair, I.; Adair, B.M.; Allan, G.M. T lymphocyte epitope mapping of porcine circovirus type 2. Viral Immunol. 2007, 20, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Hou, L.; Liu, J. Host immune response to infection with porcine circoviruses. Anim. Dis. 2021, 1, 23. [Google Scholar] [CrossRef]
- Meerts, P.; Gucht, S.V.; Cox, E.; Vandebosch, A.; Nauwynck, H.J. Correlation between type of adaptive immune response against porcine circovirus type 2 and level of virus replication. Viral Immunol. 2005, 18, 333–341. [Google Scholar] [CrossRef] [PubMed]
- The problem with neoantigen prediction. Nat. Biotechnol. 2017, 35, 97. [CrossRef]
- Stranzl, T.; Larsen, M.V.; Lundegaard, C.; Nielsen, M. NetCTLpan: Pan-specific MHC class I pathway epitope predictions. Immunogenetics 2010, 62, 357–368. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) | Length (nt) |
|---|---|---|
| 5′-flanking-RE-F | ATTGTCGACGGATCCCTGCAGGAAGCTTTTCCC | 33 |
| V5(PCV)FuF2A-F | AATCCTCTGCTGGGCCTCGACAGAGCCAAGAGAAGCGGCAGC | 42 |
| SS-PCV2b-F1 | CCCCTGGTGAGCAGCCAGTGCGTGAACCTGATGACCTACCCTAGAAGGAGG | 51 |
| SS-PCV2b-F2 | ATGTTCGTGTTCCTGGTGCTGCTGCCCCTGGTGAGCAGCCAGTGC | 45 |
| PstI-SS-F3 | CCCTGCAGGAAGCTTTTCCCCGGGGCCACCATGTTCGTGTTCCTGGTGCTG | 51 |
| V5(PCV)FuF2A-R | GCTGCCGCTTCTCTTGGCTCTGTCGAGGCCCAGCAGAGGATT | 42 |
| Stop-RE-R | TAAGCCCGGGGCGGCCGCTTATCA | 24 |
| No. | Name | Position | Genotype | Peptide | Length |
|---|---|---|---|---|---|
| 1 | Bc-Cap2a/2b-1 | 5–18 | PCV2b/PCV2a | RRRYRRRRHRPRSH | 14 |
| 2 | Bc-Cap2a/2b-2 | 25–40 | PCV2b/PCV2a | RRPWLVHPRHRYRWRR | 16 |
| 3 | Bc-Cap2b-3 | 59–70 | PCV2b | RTTVKTPSWAVD | 12 |
| 4 | Bc-Cap2b-4 | 77–120 | PCV2b | NDFLPPGGGSNPRSVPFEYYRIRKVKVEFWPCSPITQGDRGVGS | 44 |
| 5 | Bc-Cap2b-4.1 | 77–95 | PCV2b | NDFLPPGGGSNPRSVPFEY | 19 |
| 6 | Bc-Cap2a/2b-4.2 | 92–120 | PCV2b/PCV2a | PFEYYRIRKVKVEFWPCSPITQGDRGVGS | 29 |
| 7 | Bc-Cap2b-5 | 139–152 | PCV2b | DPYVNYSSRHTITQ | 14 |
| 8 | Bc-Cap2b-6 | 162–182 | PCV2b/PCV2a | TPKPVLDSTIDYFQPNNKRNQ | 21 |
| 9 | Bc-Cap2b-7 | 188–196 | PCV2b | QTAGNVDHV | 9 |
| 10 | Bc-Cap2b-8 | 201–211 | PCV2b | AFENSIYDQEY | 11 |
| 11 | Bc-Cap2a-3 | 57–67 | PCV2a | VKATTVRTPSW | 11 |
| 12 | Bc-Cap2a-4 | 77–94 | PCV2a | DDFVPPGGGTNKISIPFE | 18 |
| 13 | Bc-Cap2a-5 | 128–158 | PCV2a | NFVPKATAQTYDPYVNYSSRHTIPQPFSYHS | 31 |
| 14 | Bc-Cap2a-6 | 167–182 | PCV2b/PCV2a | LDSTIDYFQPNNKRNQ | 16 |
| 15 | Bc-Cap2a-7 | 188–196 | PCV2a | QTSRNVDHV | 9 |
| 16 | Ref-1 | 26–36 | PCV2b/PCV2a | RPWLVHPRHRY | 11 |
| 17 | Ref-2 | 117–131 | PCV2b | GVGSSAVILDDNFVT | 15 |
| 18 | Ref-3 | 195–233 | PCV2b | HVGLGTAFENSIYDQEYNIRVTMYVQFREFNLKDPPLNP | 39 |
| Peptide Name | Start-End Position | Genotype | Peptide | Length | Allele-Binder | Prediction Tool |
|---|---|---|---|---|---|---|
| CapCTL-1 | 1–8 | PCV2b, PCV2a | MTYPRRRY | 8 | SLA-1, SLA-3 | NetCTLpan |
| CapCTL-2 | 89–96 | PCV2b | RSVPFEYY | 8 | SLA-1, SLA-3 | NetCTLpan |
| CapCTL-3 | 103–111 | PCV2b, PCV2a | VEFWPCSPI | 9 | SLA-2 | NetCTLpan |
| CapCTL-4 | 136–144 | PCV2b | LTYDPYVNY | 9 | SLA-1, SLA-2 | NetCTLpan |
| CapCTL-5 | 148–156 | PCV2b | HTITQPFSY | 9 | SLA-1 | NetCTLpan |
| CapCTL-6 | 148–156 | PCV2a | HTIPQPFSY | 9 | SLA-1, SLA-2 | NetCTLpan |
| CapCTL-7 | 150–160 | PCV2b | ITQPFSYHSRY | 11 | SLA-1, SLA-2, SLA-3 | NetCTLpan |
| CapCTL-8 | 48–55 | PCV2b, PCV2a | RLSRTFGY | 8 | SLA-1, SLA-2 | NetCTLpan |
| CapCTL-9 | 89–96 | PCV2a | ISIPFEYY | 8 | SLA-1, SLA-3 | NetCTLpan |
| CapCTL-10 | 130–138 | PCV2b | VTKATALTY | 9 | SLA-1, SLA-2, SLA-3 | NetCTLpan, NetMHC |
| CapCTL-11 | 136–144 | PCV2a | QTYDPYVNY | 9 | SLA-1 | NetCTLpan |
| CapCTL-12 | 144–154 | PCV2b | YSSRHTITQPF | 11 | SLA-2 | NetCTLpan |
| CapCTL-13 | 37–45 | PCV2b/PCV2a | RWRRKNGIF | 9 | SLA-3 | NetMHC |
| CapCTL-14 | 133–141 | PCV2b | ATALTYDPY | 9 | SLA-1, SLA-2 | NetMHC |
| CapCTL-15 | 166–174 | PCV2b/PCV2a | VLDSTIDYF | 9 | SLA-1 | NetMHC |
| CapCTL-16 | 85–94 | PCV2a | GTNKISIPF | 9 | SLA-1 | NetMHC |
| CapCTL-17 | 133–141 | PCV2a | ATAQTYDPY | 9 | SLA-1, SLA-2 | NetMHC |
| H2-Cap2b2a-1 | 81–93 | PCV2b | PPGGGSNPRSVPF | 13 | H-2-Dd | NetMHC |
| H2-Cap2b2a-2 | 140–151 | PCV2b/PCV2a | PYVNYSSRHTI | 12 | H-2-Kd | NetMHC |
| H2-Cap2b2a-3 | 159–171 | PCV2b/PCV2a | RYFTPKPVLDSTI | 13 | H-2-Kd | NetMHC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meas, S.; Chaimongkolnukul, K.; Narkpuk, J.; Mekvichitsaeng, P.; Poomputsa, K.; Wanasen, N.; Roshorm, Y.M. Humoral and Cellular Immune Responses Induced by Bivalent DNA Vaccines Expressing Fusion Capsid Proteins of Porcine Circovirus Genotypes 2a and 2b. Vaccines 2024, 12, 324. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines12030324
Meas S, Chaimongkolnukul K, Narkpuk J, Mekvichitsaeng P, Poomputsa K, Wanasen N, Roshorm YM. Humoral and Cellular Immune Responses Induced by Bivalent DNA Vaccines Expressing Fusion Capsid Proteins of Porcine Circovirus Genotypes 2a and 2b. Vaccines. 2024; 12(3):324. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines12030324
Chicago/Turabian StyleMeas, Sochanwattey, Khuanjit Chaimongkolnukul, Jaraspim Narkpuk, Phenjun Mekvichitsaeng, Kanokwan Poomputsa, Nanchaya Wanasen, and Yaowaluck Maprang Roshorm. 2024. "Humoral and Cellular Immune Responses Induced by Bivalent DNA Vaccines Expressing Fusion Capsid Proteins of Porcine Circovirus Genotypes 2a and 2b" Vaccines 12, no. 3: 324. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines12030324