Evaluation of A Baculovirus-Expressed VP2 Subunit Vaccine for the Protection of White-Tailed Deer (Odocoileus virginianus) from Epizootic Hemorrhagic Disease

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
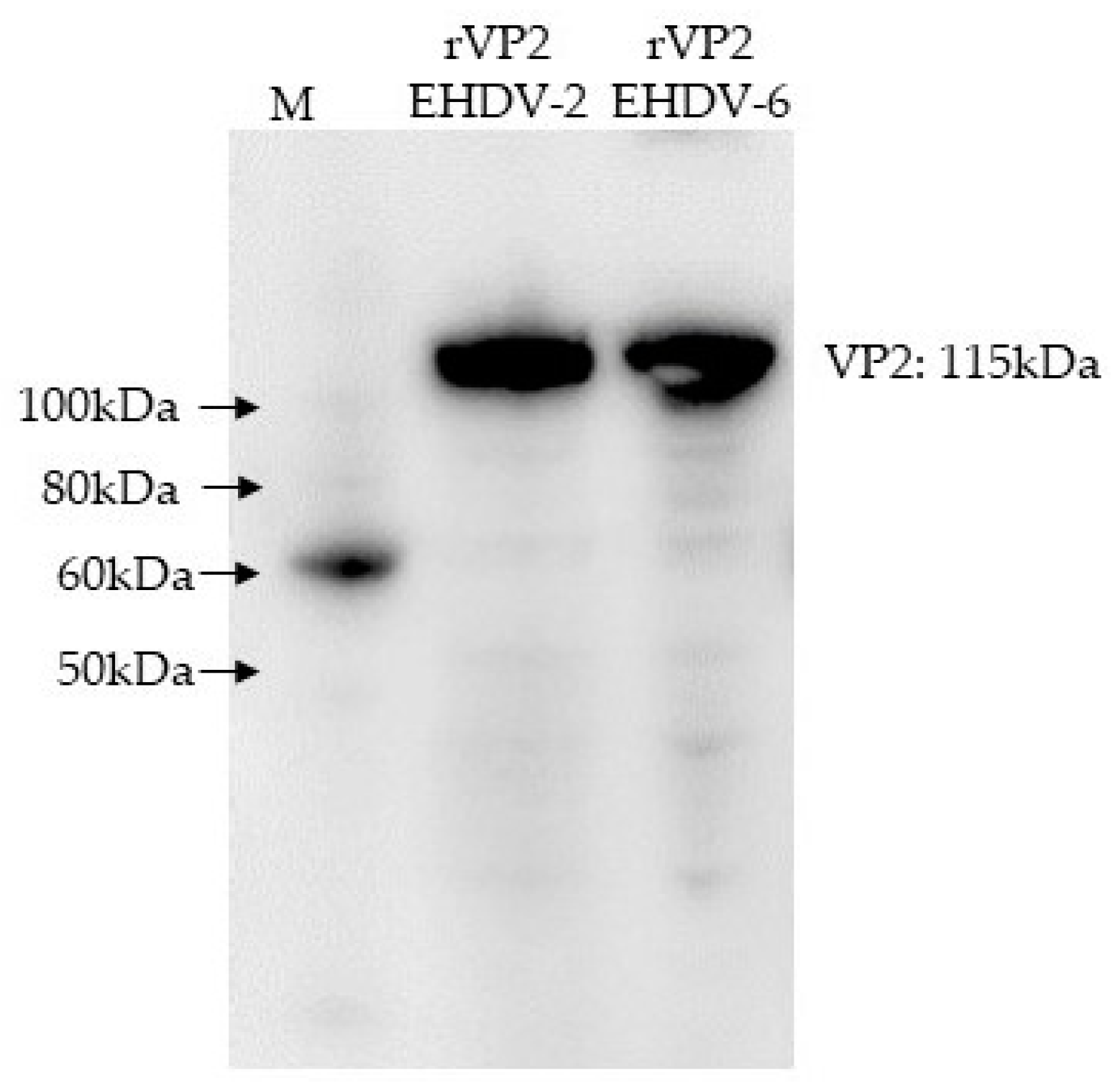
2.2. Generation of EHDV Recombinant Proteins
2.3. Animals and Experimental Design
2.3.1. Mice
2.3.2. Cattle
2.3.3. White-Tailed Deer (WTD)
2.3.4. Postmortem Examination and Analysis
2.4. Viruses
2.5. Virus Neutralization Assay
2.6. Virus Detection
2.6.1. Real-time Quantitative PCR
2.6.2. Virus Plaque Assay
2.7. Statistical Analyses
3. Results
3.1. Neutralizing Antibody Responses in Vaccinated Mice
3.2. Neutralizing Antibody Responses in Vaccinated Cattle
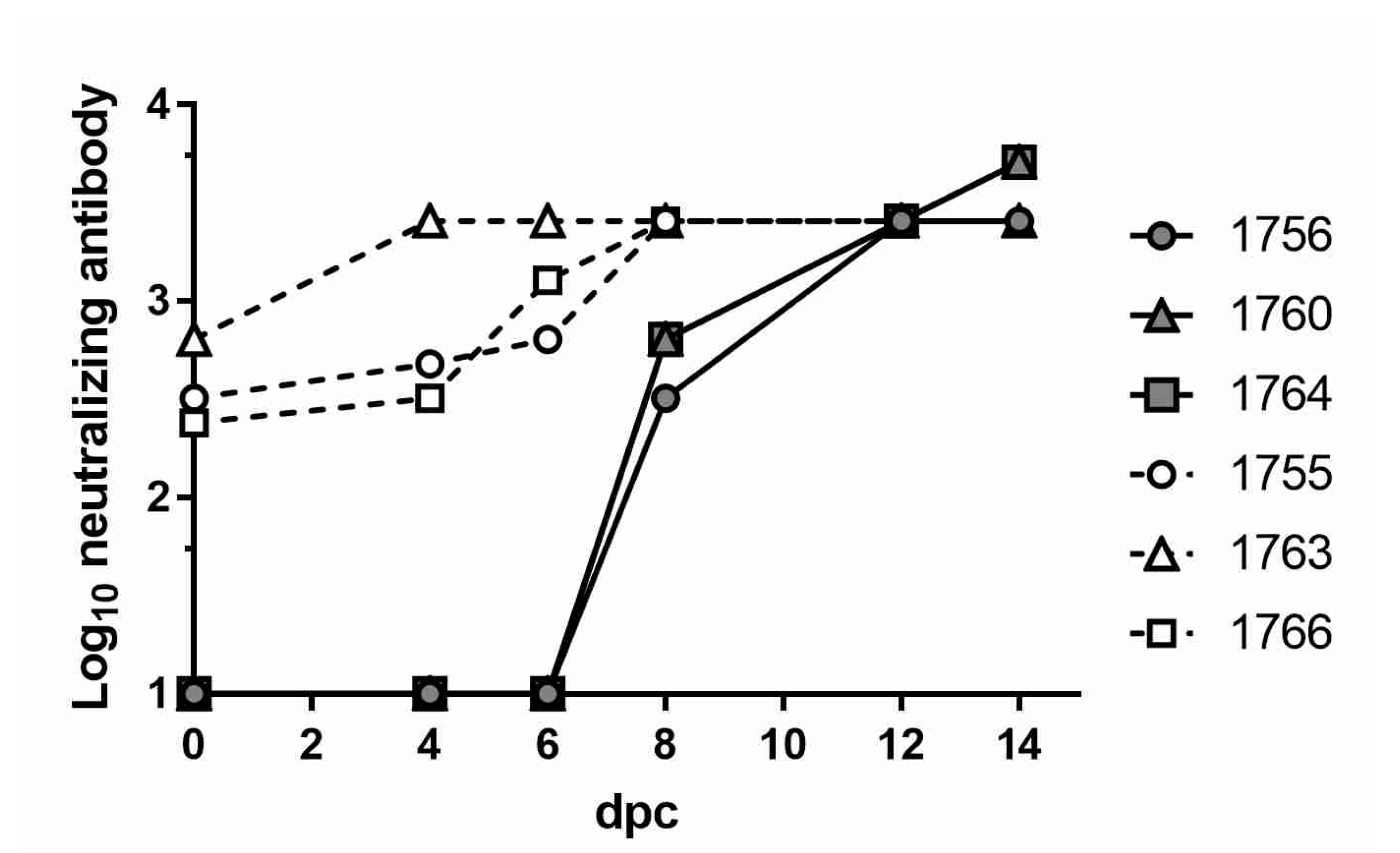
3.3. Neutralizing Antibody Responses in Vaccinated and EHDV-Challenged WTD
3.4. Clinical Parameters in EHDV-2 Challenged WTD
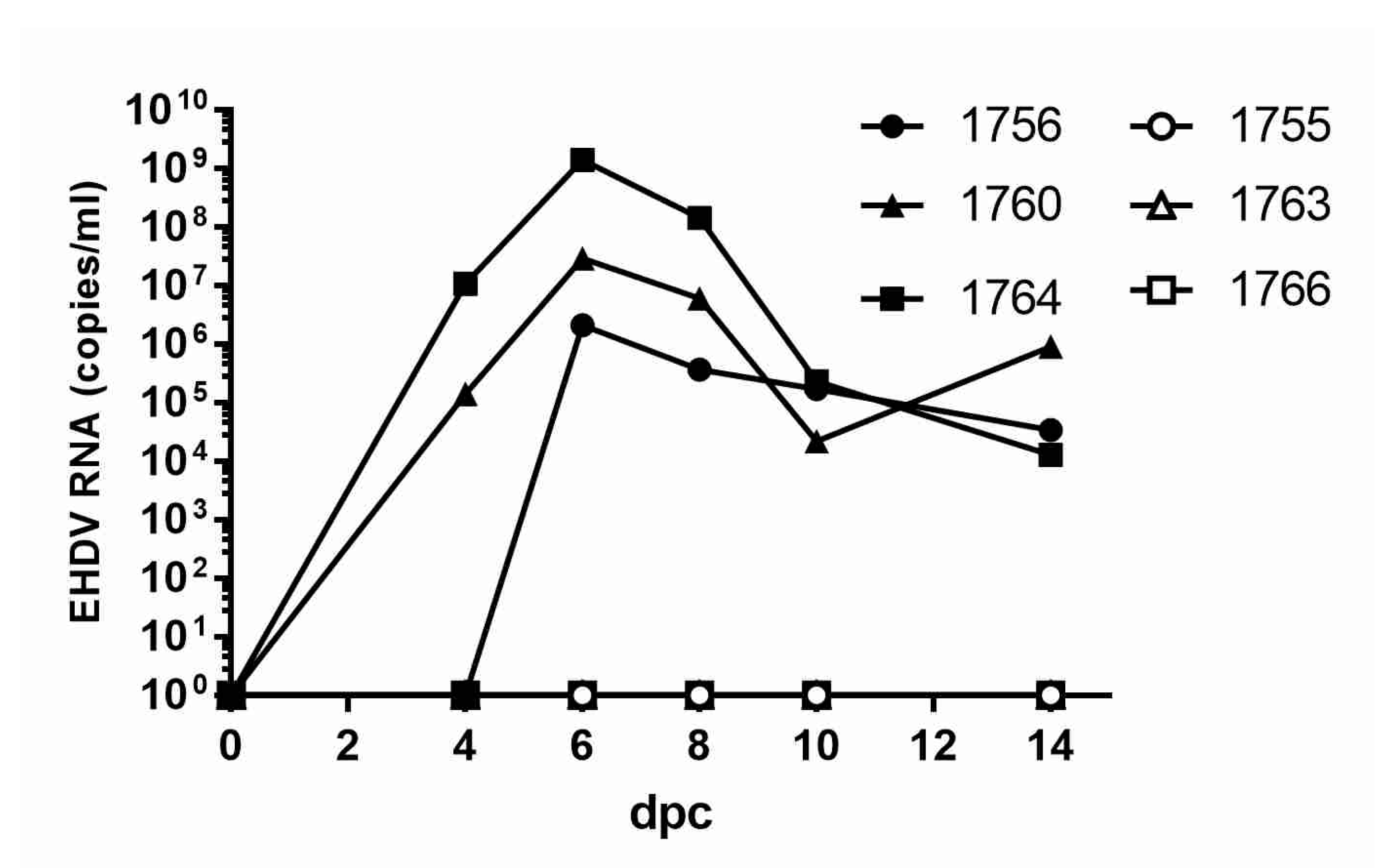
3.5. Blood Viral RNA Loads and Viremia in Vaccinated/Challenged WTD
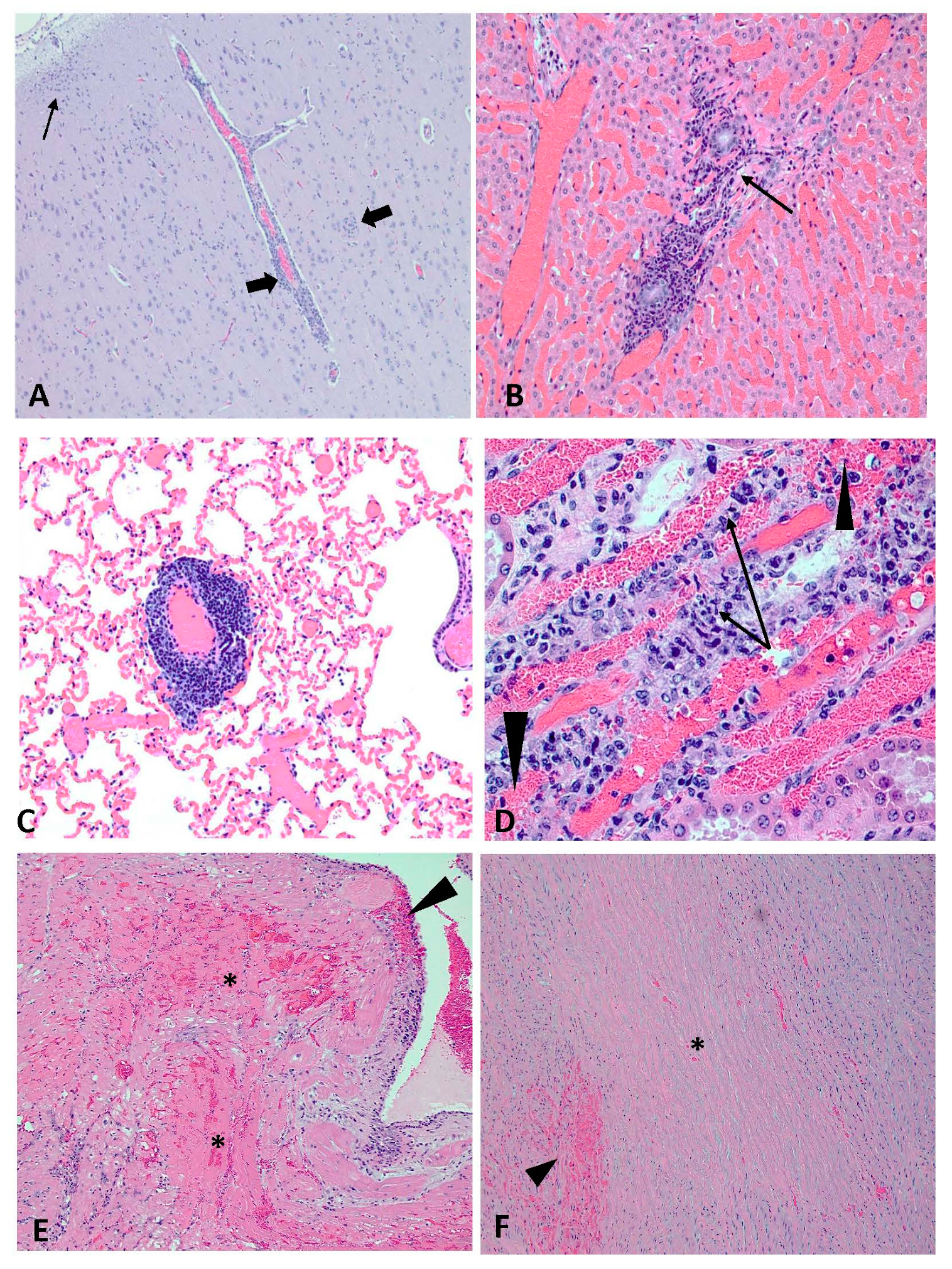
3.6. Postmortem Analyses and Detection of Viral RNA in Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mertens, P.P.C.; Maan, S.; Samuel, A.; Attoui, H. Orbivirus, Reoviridae, Virus Taxonomy, VIIIth Report of the ICTV, 2nd ed.; Elsevier Academic Press: San Diego, CA, USA, 2005; pp. 466–483. [Google Scholar]
- Savini, G.; Afonso, A.; Mellor, P.; Aradaib, I.; Yadin, H.; Sanaa, M.; Wilson, W.; Monaco, F.; Domingo, M. Epizootic heamorragic disease. Res. Vet. Sci. 2011, 91, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Pfannenstiel, R.S.; Mullens, B.A.; Ruder, M.G.; Zurek, L.; Cohnstaedt, L.W.; Nayduch, D. Management of North American Culicoides Biting Midges: Current Knowledge and Research Needs. Vector Borne Zoonotic Dis. 2015, 15, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Anthony, S.J.; Maan, S.; Maan, N.; Kgosana, L.; Bachanek-Bankowska, K.; Batten, C.; Darpel, K.E.; Sutton, G.; Attoui, H.; Mertens, P.P. Genetic and phylogenetic analysis of the outer-coat proteins VP2 and VP5 of epizootic haemorrhagic disease virus (EHDV): comparison of genetic and serological data to characterise the EHDV serogroup. Virus Res. 2009, 145, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Ruder, M.G.; Lysyk, T.J.; Stallknecht, D.E.; Foil, L.D.; Johnson, D.J.; Chase, C.C.; Dargatz, D.A.; Gibbs, E.P. Transmission and Epidemiology of Bluetongue and Epizootic Hemorrhagic Disease in North America: Current Perspectives, Research Gaps, and Future Directions. Vector Borne Zoonotic Dis. 2015, 15, 348–363. [Google Scholar] [CrossRef]
- Crum, J.A.; Mead, D.G.; Jackwood, M.W.; Phillips, J.E.; Stallknecht, D.E. Genetic Relatedness of Epizootic Hemorrhagic Disease Virus Serotype 2 from 2012 Outbreak in the USA. J. Wildl. Dis. 2018. [Google Scholar] [CrossRef]
- Allison, A.B.; Goekjian, V.H.; Potgieter, A.C.; Wilson, W.C.; Johnson, D.J.; Mertens, P.P.; Stallknecht, D.E. Detection of a novel reassortant epizootic hemorrhagic disease virus (EHDV) in the USA containing RNA segments derived from both exotic (EHDV-6) and endemic (EHDV-2) serotypes. J. Gen. Virol. 2010, 91, 430–439. [Google Scholar] [CrossRef]
- Stallknecht, D.E.; Allison, A.B.; Park, A.W.; Phillips, J.E.; Goekjian, V.H.; Nettles, V.F.; Fischer, J.R. Apparent increase of reported hemorrhagic disease in the midwestern and northeastern USA. J. Wildl. Dis. 2015, 51, 348–361. [Google Scholar] [CrossRef]
- Ruder, M.G.; Johnson, D.; Ostlund, E.; Allison, A.B.; Kienzle, C.; Phillips, J.E.; Poulson, R.L.; Stallknecht, D.E. The First 10 Years (2006-15) of Epizootic Hemorrhagic Disease Virus Serotype 6 in the USA. J. Wildl. Dis. 2017, 53, 901–905. [Google Scholar] [CrossRef]
- Stallknecht, D.E.; Howerth, E.W. Epidemiology of bluetongue and epizootic haemorrhagic disease in wildlife: surveillance methods. Vet. Ital. 2004, 40, 203–207. [Google Scholar]
- Flacke, G.L.; Yabsley, M.J.; Hanson, B.A.; Stallknecht, D.E. Hemorrhagic disease in Kansas: enzootic stability meets epizootic disease. J. Wildl. Dis. 2004, 40, 288–293. [Google Scholar] [CrossRef] [Green Version]
- Gaydos, J.K.; Crum, J.M.; Davidson, W.R.; Cross, S.S.; Owen, S.F.; Stallknecht, D.E. Epizootiology of an epizootic hemorrhagic disease outbreak in West Virginia. J. Wildl. Dis. 2004, 40, 383–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.; Outlaw, J.; Earle, M.; Richardson, J. Economic Impact of U.S. Deer Breeding and Hunting Operations; 17-4; Texas A&M University: College Station, TX, USA, 2017. [Google Scholar]
- Cauvin, A.; Dinh, E.T.N.; Orange, J.P.; Shuman, R.M.; Blackburn, J.K.; Wisely, S.M. Antibodies to Epizootic Hemorrhagic Disease Virus (EHDV) in Farmed and Wild Florida White-Tailed Deer (Odocoileus virginianus). J. Wildl. Dis. 2020, 1, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Inaba, U. Ibaraki disease and its relationship to bluetongue. Aust. Vet. J. 1975, 51, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Maclachlan, N.J.; Zientara, S.; Wilson, W.C.; Richt, J.A.; Savini, G. Bluetongue and epizootic hemorrhagic disease viruses: recent developments with these globally re-emerging arboviral infections of ruminants. Curr. Opin. Virol. 2019, 34, 56–62. [Google Scholar] [CrossRef]
- Maclachlan, N.J.; Zientara, S.; Savini, G.; Daniels, P.W. Epizootic haemorrhagic disease. Rev. Sci. Tech. 2015, 34, 341–351. [Google Scholar] [CrossRef] [Green Version]
- World Organisation for Animal Health (OIE) Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Available online: http://www.oie.int/standard-setting/terrestrial-manual/access-online/ (accessed on 7 December 2019).
- McVey, D.S.; MacLachlan, N.J. Vaccines for Prevention of Bluetongue and Epizootic Hemorrhagic Disease in Livestock: A North American Perspective. Vector Borne Zoonotic Dis 2015, 15, 385–396. [Google Scholar] [CrossRef]
- Wilson, W.C.; Ruder, M.G.; Klement, E.; Jasperson, D.C.; Yadin, H.; Stallknecht, D.E.; Mead, D.G.; Howerth, E. Genetic characterization of epizootic hemorrhagic disease virus strains isolated from cattle in Israel. J. Gen. Virol. 2015, 96, 1400–1410. [Google Scholar] [CrossRef] [Green Version]
- Feenstra, F.; van Rijn, P.A. Current and next-generation bluetongue vaccines: Requirements, strategies, and prospects for different field situations. Crit. Rev. Microbiol. 2017, 43, 142–155. [Google Scholar] [CrossRef]
- Calvo-Pinilla, E.; Castillo-Olivares, J.; Jabbar, T.; Ortego, J.; de la Poza, F.; Marin-Lopez, A. Recombinant vaccines against bluetongue virus. Virus Res. 2014, 182, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Roy, P. Multiple gene expression in baculovirus system. Third generation vaccines for bluetongue disease and African horsesickness disease. Ann. N. Y. Acad. Sci. 1996, 791, 318–332. [Google Scholar] [CrossRef]
- Roy, P.; Urakawa, T.; Van Dijk, A.A.; Erasmus, B.J. Recombinant virus vaccine for bluetongue disease in sheep. J. Virol. 1990, 64, 1998–2003. [Google Scholar] [CrossRef] [Green Version]
- Roy, P.; Bishop, D.H.; Howard, S.; Aitchison, H.; Erasmus, B. Recombinant baculovirus-synthesized African horsesickness virus (AHSV) outer-capsid protein VP2 provides protection against virulent AHSV challenge. J. Gen. Virol. 1996, 77, 2053–2057. [Google Scholar] [CrossRef]
- Wilson, W.C.; Fukusho, A.; Roy, P. Diagnostic complementary DNA probes for genome segments 2 and 3 of epizootic hemorrhagic disease virus serotype 1. Am. J. Vet. Res. 1990, 51, 855–860. [Google Scholar]
- Palmer, M.V.; Cox, R.J.; Waters, W.R.; Thacker, T.C.; Whipple, D.L. Using White-tailed Deer (Odocoileus virginianus) in Infectious Disease Research. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 350–360. [Google Scholar]
- Mills, M.K.; Ruder, M.G.; Nayduch, D.; Michel, K.; Drolet, B.S. Dynamics of epizootic hemorrhagic disease virus infection within the vector, Culicoides sonorensis (Diptera: Ceratopogonidae). PLoS ONE 2017, 12, e0188865. [Google Scholar] [CrossRef]
- McHolland, L.E.; Mecham, J.O. Characterization of cell lines developed from field populations of Culicoides sonorensis (Diptera: Ceratopogonidae). J. Med. Entomol. 2003, 40, 348–351. [Google Scholar] [CrossRef] [Green Version]
- Wilson, W.C.; O’Hearn, E.S.; Tellgren-Roth, C.; Stallknecht, D.E.; Mead, D.G.; Mecham, J.O. Detection of all eight serotypes of Epizootic hemorrhagic disease virus by real-time reverse transcription polymerase chain reaction. J. Vet. Diagn. Invest. 2009, 21, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Quist, C.F.; Howerth, E.W.; Stallknecht, D.E.; Brown, J.; Pisell, T.; Nettles, V.F. Host defense responses associated with experimental hemorrhagic disease in white-tailed deer. J. Wildl. Dis. 1997, 33, 584–599. [Google Scholar] [CrossRef] [Green Version]
- Weyer, C.T.; Grewar, J.D.; Burger, P.; Rossouw, E.; Lourens, C.; Joone, C.; le Grange, M.; Coetzee, P.; Venter, E.; Martin, D.P.; et al. African Horse Sickness Caused by Genome Reassortment and Reversion to Virulence of Live, Attenuated Vaccine Viruses, South Africa, 2004-2014. Emerg. Infect. Dis. 2016, 22, 2087–2096. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Sabara, M.I. Production of a recombinant major inner capsid protein for serological detection of epizootic hemorrhagic disease virus. Clin. Diagn. Lab. Immunol. 2005, 12, 904–909. [Google Scholar] [CrossRef] [Green Version]
- Mecham, J.O.; Wilson, W.C. Antigen capture competitive enzyme-linked immunosorbent assays using baculovirus-expressed antigens for diagnosis of bluetongue virus and epizootic hemorrhagic disease virus. J. Clin. Microbiol. 2004, 42, 518–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, P.; Bishop, D.H.; LeBlois, H.; Erasmus, B.J. Long-lasting protection of sheep against bluetongue challenge after vaccination with virus-like particles: evidence for homologous and partial heterologous protection. Vaccine 1994, 12, 805–811. [Google Scholar] [CrossRef]
- Alshaikhahmed, K.; Roy, P. Generation of virus-like particles for emerging epizootic haemorrhagic disease virus: Towards the development of safe vaccine candidates. Vaccine 2016, 34, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Huismans, H.; van Der Walt, N.T.; Cloete, M.; Erasmus, B.J. Isolation of a capsid protein of bluetongue virus that induces a protective immune response in sheep. Virology 1987, 157, 172–179. [Google Scholar] [CrossRef]
- Mertens, P.P.C.; Pedley, S.; Cowley, J.; Burroughs, J.N.; Corteyn, A.H.; Jeggo, M.H.; Jennings, D.M.; Gorman, B.M. Analysis of the roles of bluetongue virus outer capsid proteins VP2 and VP5 in determination of virus serotype. Virology 1989, 170, 561–565. [Google Scholar] [CrossRef]
- Mendiola, S.Y.; Mills, M.K.; Maki, E.; Drolet, B.S.; Wilson, W.C.; Berghaus, R.; Stallknecht, D.E.; Breitenbach, J.; McVey, D.S.; Ruder, M.G. EHDV-2 Infection Prevalence Varies in Culicoides sonorensis after Feeding on Infected White-Tailed Deer over the Course of Viremia. Viruses 2019, 11, 371. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study 1 | Sera Collection | Vaccine Group | ||
| Adjuvant Control | Inactivated EHDV-2 | EHDV-2 rVP2 | ||
| 0 dpv | - | - | - | |
| 35 dpv | - | - | 53.33 ± 19.3 | |
| Study 2 | Vaccine Group | |||
| Adjuvant Control | Inactivated EHDV-6 | EHDV-6 rVP2 | ||
| 0 dpv | - | - | - | |
| 35 dpv | - | - | 1800 ± 1006 | |
| Vaccine Group | α-EHDV-2 Titers | α-EHDV-6 Titers | ||||
| 0 dpv | 21 dpv | 35 dpv | 0 dpv | 21 dpv | 35 dpv | |
| Adjuvant Control | - | - | - | - | - | - |
| EHDV-2 rVP2 | - | 10 ± 16.0 | 706.7 ± 384 | - | - | 5.8 ± 5.2 |
| EHDV-6 rVP2 | - | - | 4.2 ± 7.0 | - | 5.8 ± 8.0 | 226.7 ± 83 |
| Tissue | Sham-Vaccinated WTD | rVP2-Vaccinated WTD | ||||
|---|---|---|---|---|---|---|
| #1756 | #1760 | #1764 | #1755 | #1763 | #1766 | |
| Lung | 3.37 | 2.72 | 5.75 | ND | ND | ND |
| Spleen | ND | 2.63 | 3.78 | ND | ND | ND |
| Liver | ND | 3.18 (S) | 3.19 | ND | ND | ND |
| Kidney | 2.53 | 3.43 | ND | ND | ND | ND |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sunwoo, S.Y.; Noronha, L.E.; Morozov, I.; Trujillo, J.D.; Kim, I.J.; Schirtzinger, E.E.; Faburay, B.; Drolet, B.S.; Urbaniak, K.; McVey, D.S.; et al. Evaluation of A Baculovirus-Expressed VP2 Subunit Vaccine for the Protection of White-Tailed Deer (Odocoileus virginianus) from Epizootic Hemorrhagic Disease. Vaccines 2020, 8, 59. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8010059
Sunwoo SY, Noronha LE, Morozov I, Trujillo JD, Kim IJ, Schirtzinger EE, Faburay B, Drolet BS, Urbaniak K, McVey DS, et al. Evaluation of A Baculovirus-Expressed VP2 Subunit Vaccine for the Protection of White-Tailed Deer (Odocoileus virginianus) from Epizootic Hemorrhagic Disease. Vaccines. 2020; 8(1):59. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8010059
Chicago/Turabian StyleSunwoo, Sun Young, Leela E. Noronha, Igor Morozov, Jessie D. Trujillo, In Joong Kim, Erin E. Schirtzinger, Bonto Faburay, Barbara S. Drolet, Kinga Urbaniak, D. Scott McVey, and et al. 2020. "Evaluation of A Baculovirus-Expressed VP2 Subunit Vaccine for the Protection of White-Tailed Deer (Odocoileus virginianus) from Epizootic Hemorrhagic Disease" Vaccines 8, no. 1: 59. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8010059