Immunogenicity and Efficacy of a Recombinant Human Adenovirus Type 5 Vaccine against Zika Virus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Viruses, Human Serum and Cells
2.3. Zika Virus Sequence Analysis
2.4. Recombinant Replication Deficient Adenovirus Type 5
2.5. Focus Forming Assay
2.6. Western Blot
2.7. Antigen-Specific T Cell Staining
2.8. ELISA
2.9. Vaccination with Recombinant Replication Deficient Adenovirus Type 5
2.10. Animal Challenge
2.11. Clinical Monitoring
2.12. Statistical Analysis
3. Results
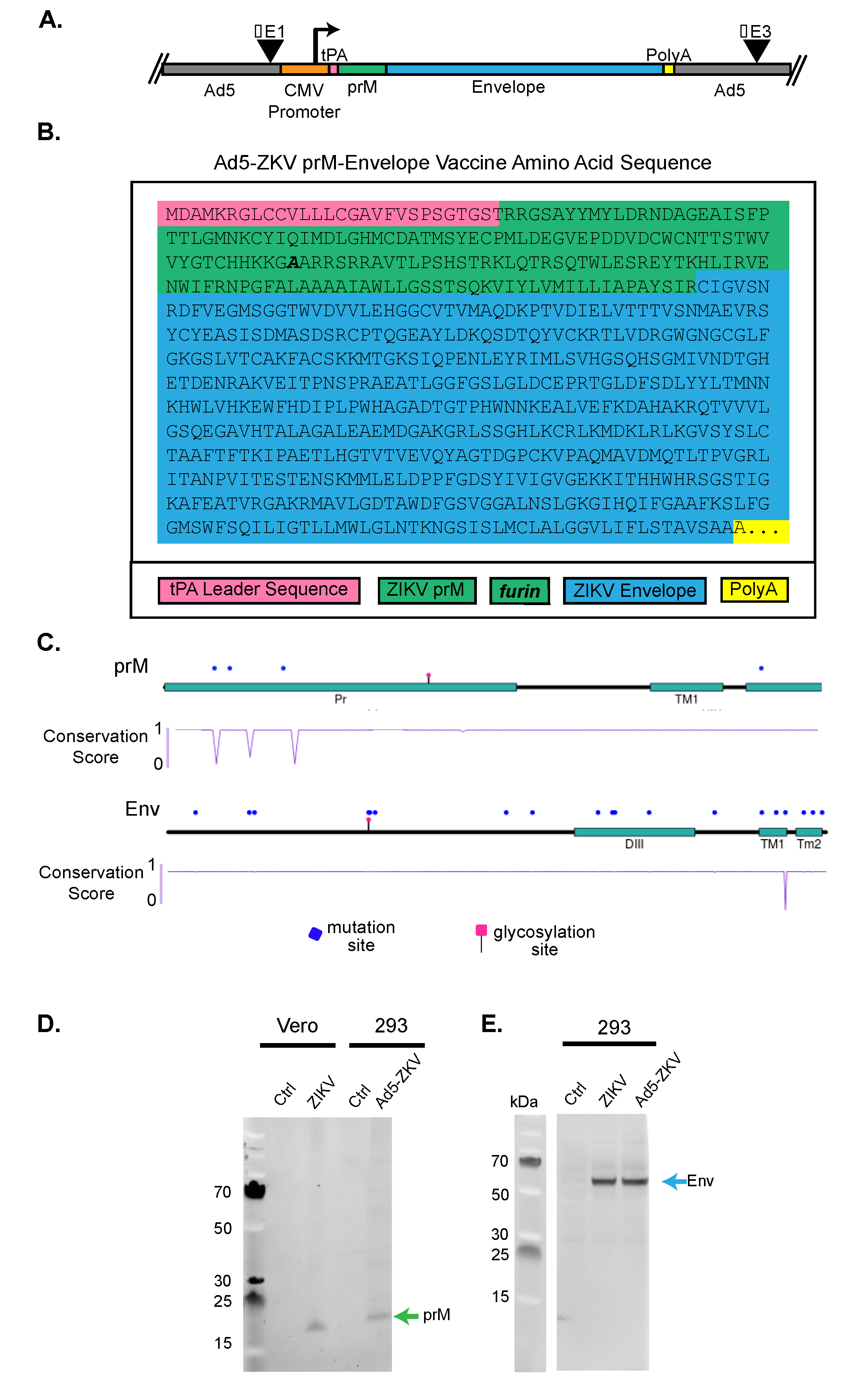
3.1. Generation of Recombinant Human Adenovirus Type 5 ZIKV Vaccine
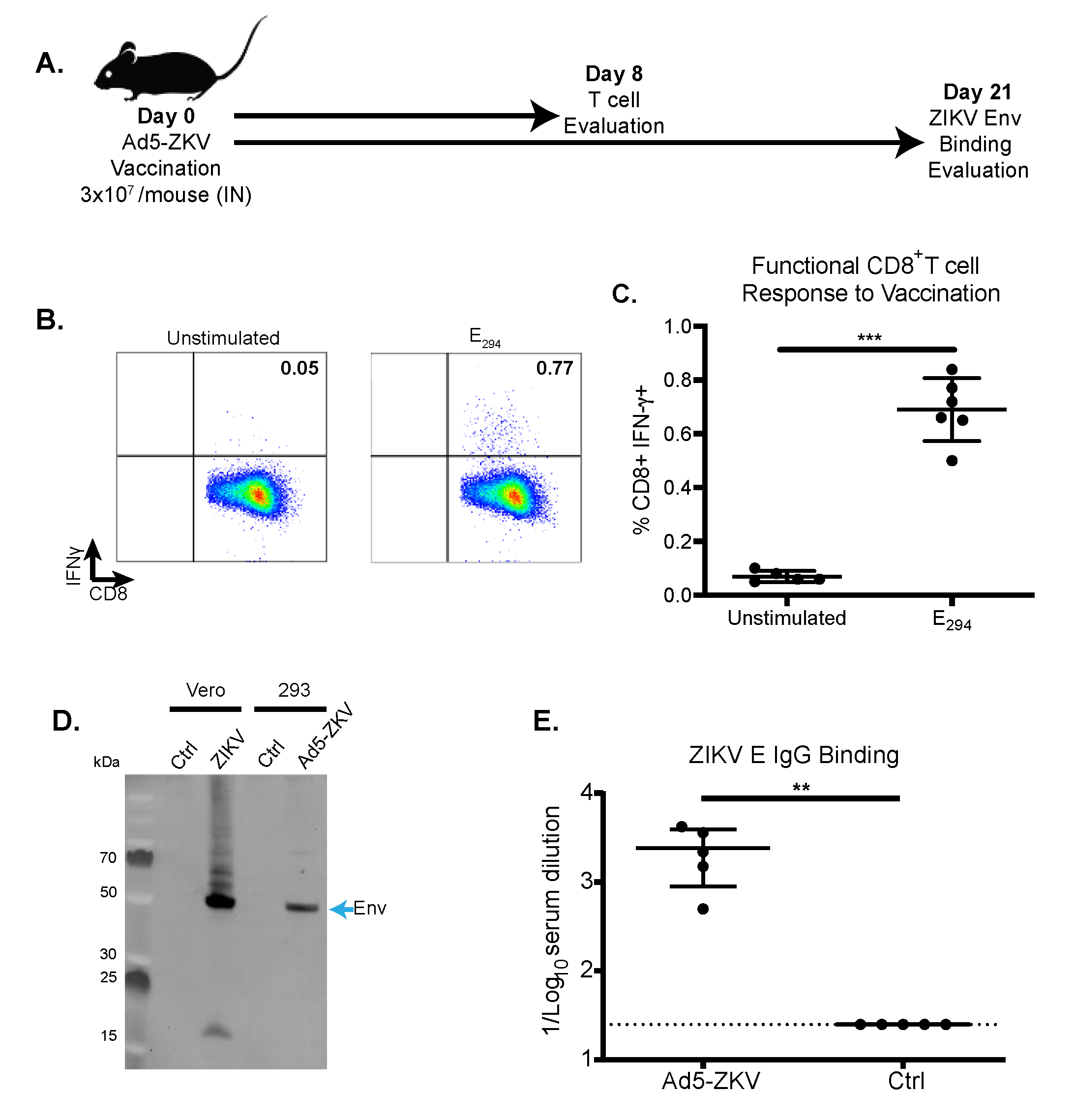
3.2. Immunogenicity of hAd5-ZKV Vaccine
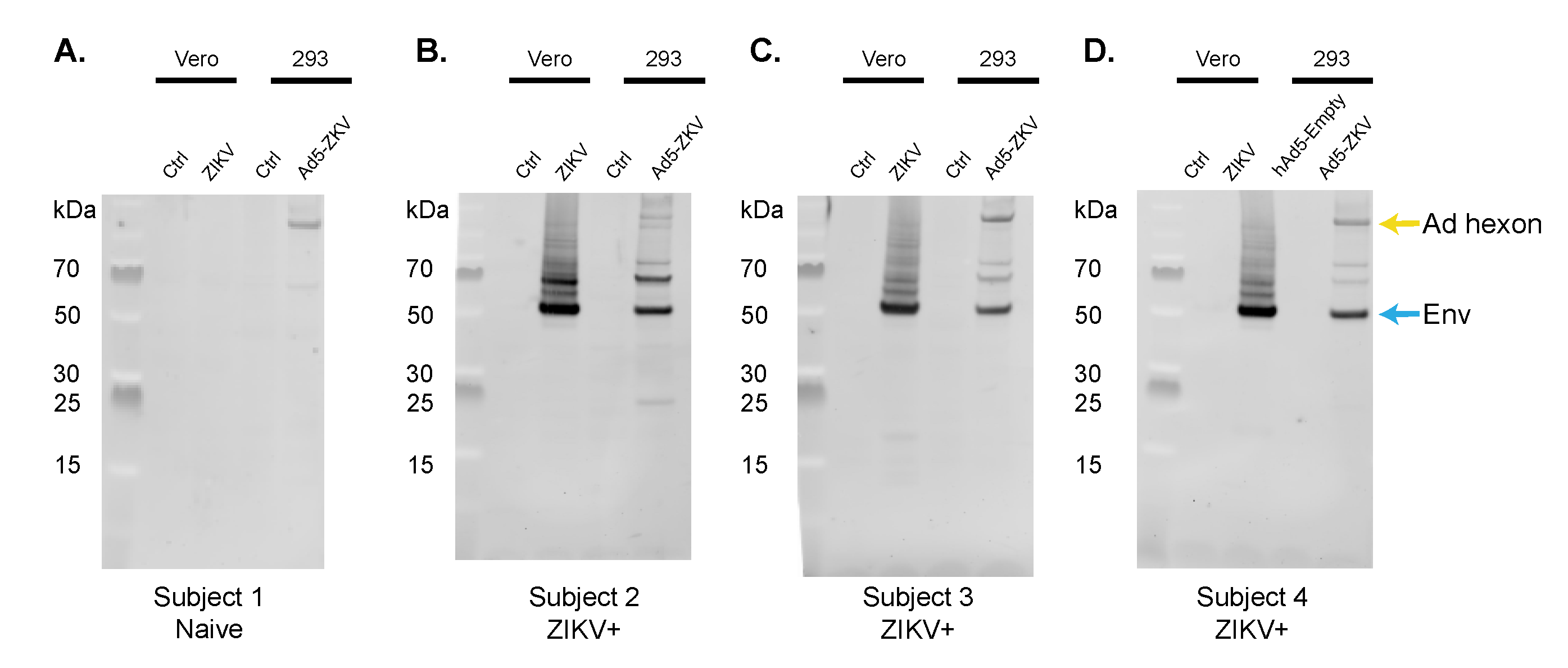
3.3. Immune Recognition by the Human Polyclonal Antibody Response
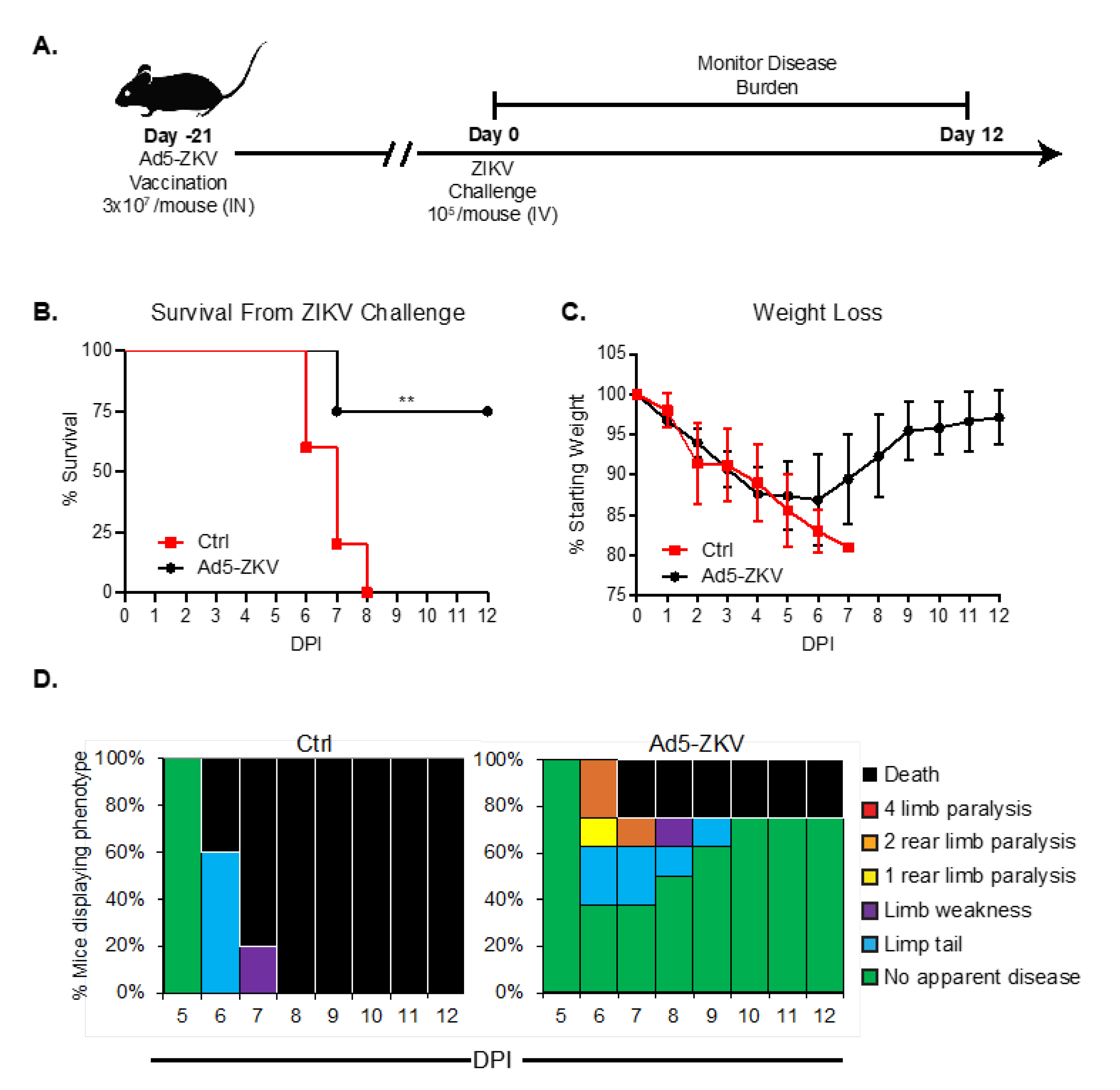
3.4. Protective Capacity of hAd5-ZKV Vaccine
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lazear, H.M.; Diamond, M.S. Zika Virus: New Clinical Syndromes and Its Emergence in the Western Hemisphere. J. Virol. 2016, 90, 4864–4875. [Google Scholar] [CrossRef] [Green Version]
- Boorman, J.P.; Porterfield, J.S. A Simple Technique for Infection of Mosquitoes with Viruses; Transmission of Zika Virus. Trans. R. Soc. Trop. Med. Hyg. 1956, 50, 238–242. [Google Scholar] [CrossRef]
- Hills, S.L.; Russell, K.; Hennessey, M.; Williams, C.; Oster, A.M.; Fischer, M.; Mead, P. Transmission of Zika Virus Through Sexual Contact with Travelers to Areas of Ongoing Transmission—Continental United States, 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 215–216. [Google Scholar] [CrossRef] [Green Version]
- Brasil, P.; Pereira, J.P., Jr.; Moreira, M.E.; Ribeiro Nogueira, R.M.; Damasceno, L.; Wakimoto, M.; Rabello, R.S.; Valderramos, S.G.; Halai, U.A.; Salles, T.S.; et al. Zika Virus Infection in Pregnant Women in Rio de Janeiro. N. Engl. J. Med. 2016, 375, 2321–2334. [Google Scholar] [CrossRef]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef]
- Olagnier, D.; Muscolini, M.; Coyne, C.B.; Diamond, M.S.; Hiscott, J. Mechanisms of Zika Virus Infection and Neuropathogenesis. DNA Cell Biol. 2016, 35, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Brasil, P.; Sequeira, P.C.; Freitas, A.D.; Zogbi, H.E.; Calvet, G.A.; de Souza, R.V.; Siqueira, A.M.; de Mendonca, M.C.; Nogueira, R.M.; de Filippis, A.M.; et al. Guillain-Barre Syndrome Associated with Zika Virus Infection. Lancet 2016, 387, 1482. [Google Scholar] [CrossRef] [Green Version]
- Cao-Lormeau, V.M.; Blake, A.; Mons, S.; Lastere, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barre Syndrome Outbreak Associated with Zika Virus Infection in French Polynesia: A Case-Control Study. Lancet 2016, 387, 1531–1539. [Google Scholar] [CrossRef] [Green Version]
- Krauer, F.; Riesen, M.; Reveiz, L.; Oladapo, O.T.; Martínez-Vega, R.; Porgo, T.V.; Haefliger, A.; Broutet, N.J.; Low, N.; WHO Zika Causality Working Group(W.H.O.Z.C.W.). Zika Virus Infection as a Cause of Congenital Brain Abnormalities and Guillain-Barré Syndrome: Systematic Review. PLoS Med. 2017, 14, e1002203. [Google Scholar] [CrossRef] [Green Version]
- Watrin, L.; Ghawche, F.; Larre, P.; Neau, J.P.; Mathis, S.; Fournier, E. Guillain-Barre Syndrome (42 Cases) Occurring During a Zika Virus Outbreak in French Polynesia. Medicine 2016, 95, e3257. [Google Scholar] [CrossRef]
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika Virus Outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef]
- Cao-Lormeau, V.M.; Roche, C.; Teissier, A.; Robin, E.; Berry, A.L.; Mallet, H.P.; Sall, A.A.; Musso, D. Zika Virus, French polynesia, South Pacific, 2013. Emerg. Infect. Dis. 2014, 20, 1085–1086. [Google Scholar] [CrossRef]
- Zanluca, C.; de Melo, V.C.A.; Mosimann, A.L.P.; Santos, G.I.V.D.; Santos, C.N.D.D.; Luz, K. First Report of Autochthonous Transmission of Zika Virus in Brazil. Mem. Inst. Oswaldo Cruz 2015, 110, 569–572. [Google Scholar] [CrossRef]
- Diamond, M.S.; Ledgerwood, J.E.; Pierson, T.C. Zika Virus Vaccine Development: Progress in the Face of New Challenges. Annu. Rev. Med. 2019, 70, 121–135. [Google Scholar] [CrossRef]
- Elong Ngono, A.; Shresta, S. Cross-Reactive T Cell Immunity to Dengue and Zika Viruses: New Insights Into Vaccine Development. Front. Immunol. 2019, 10, 1316. [Google Scholar] [CrossRef] [Green Version]
- Richner, J.M.; Diamond, M.S. Zika Virus Vaccines: Immune Response, Current Status, and Future Challenges. Curr. Opin. Immunol. 2018, 53, 130–136. [Google Scholar] [CrossRef]
- Erbelding, E.; Cassetti, C. Zika Virus and Future Research Directions. J. Infect. Dis. 2017, 216, S991–S994. [Google Scholar] [CrossRef] [Green Version]
- Hassert, M.; Brien, J.D.; Pinto, A.K. Mouse Models of Heterologous Flavivirus Immunity: A Role for Cross-Reactive T Cells. Front. Immunol. 2019, 10, 1045. [Google Scholar] [CrossRef] [Green Version]
- Hassert, M.; Harris, M.G.; Brien, J.D.; Pinto, A.K. Identification of Protective CD8 T Cell Responses in a Mouse Model of Zika Virus Infection. Front. Immunol. 2019, 10, 1678. [Google Scholar] [CrossRef] [Green Version]
- Jurado, K.A.; Yockey, L.J.; Wong, P.W.; Lee, S.; Huttner, A.J.; Iwasaki, A. Antiviral CD8 T cells induce Zika-virus-associated paralysis in mice. Nat. Microbiol. 2018, 3, 141–147. [Google Scholar] [CrossRef]
- Perez-Guzman, E.X.; Pantoja, P.; Serrano-Collazo, C.; Hassert, M.A.; Ortiz-Rosa, A.; Rodriguez, I.V.; Giavedoni, L.; Hodara, V.; Parodi, L.; Cruz, L.; et al. Time Elapsed between Zika and Dengue Virus Infections Affects Antibody and T Cell Responses. Nat. Commun. 2019, 10, 4316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassert, M.; Wolf, K.J.; Schwetye, K.E.; DiPaolo, R.J.; Brien, J.D.; Pinto, A.K. CD4+T Cells Mediate Protection Against Zika Associated Severe Disease in a Mouse Model of Infection. PLoS Pathog. 2018, 14, e1007237. [Google Scholar] [CrossRef] [PubMed]
- Brien, J.D.; Austin, S.K.; Sukupolvi-Petty, S.; O’Brien, K.M.; Johnson, S.; Fremont, D.H.; Diamond, M.S. Genotype-Specific Neutralization and Protection by Antibodies Against Dengue Virus type 3. J. Virol. 2010, 84, 10630–10643. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, B.; Brien, J.D.; Sukupolvi-Petty, S.; Austin, S.K.; Edeling, M.A.; Kim, T.; O’Brien, K.M.; Nelson, C.A.; Johnson, S.; Fremont, D.H.; et al. The Development of Therapeutic Antibodies that Neutralize Homologous and Heterologous Genotypes of Dengue Virus Type 1. PLoS Pathog. 2010, 6, e1000823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brien, J.D.; Sukupolvi-Petty, S.; Williams, K.L.; Lam, C.Y.; Schmid, M.A.; Johnson, S.; Harris, E.; Diamond, M.S. Protection by Immunoglobulin Dual-Affinity Retargeting Antibodies Against Dengue Virus. J. Virol. 2013, 87, 7747–7753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, M.J.; Duehr, J.; Dulin, H.; Broecker, F.; Brown, J.A.; Arumemi, F.O.; Bermúdez González, M.C.; Leyva-Grado, V.H.; Evans, M.J.; Simon, V.; et al. Human Antibodies Targeting Zika Virus NS1 Provide Protection Against Disease in a Mouse Model. Nat. Commun. 2018, 9, 4560. [Google Scholar] [CrossRef] [Green Version]
- Branche, E.; Simon, A.Y.; Sheets, N.; Kim, K.; Barker, D.; Nguyen, A.-V.T.; Sahota, H.; Young, M.P.; Salgado, R.; Mamidi, A.; et al. Human Polyclonal Antibodies Prevent Lethal Zika Virus Infection in Mice. Sci. Rep. 2019, 9, 9857. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Li, S.; Zhang, Y.; Han, X.; Jia, B.; Liu, H.; Liu, D.; Tan, S.; Wang, Q.; Bi, Y.; et al. CD8+ T Cell Immune Response in Immunocompetent Mice during Zika Virus Infection. J. Virol. 2017, 91, e00900–00917. [Google Scholar] [CrossRef] [Green Version]
- Beasley, D.W.; Lewthwaite, P.; Solomon, T. Current Use and Development of Vaccines for Japanese Encephalitis. Expert Opin. Biol. Ther. 2008, 8, 95–106. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Johnson, N.; Phipps, L.P.; Stephenson, J.R.; Fooks, A.R.; Solomon, T. Tick-Borne Encephalitis Virus - a Review of an Emerging Zoonosis. J. Gen. Virol. 2009, 90, 1781–1794. [Google Scholar] [CrossRef]
- Guy, B.; Guirakhoo, F.; Barban, V.; Higgs, S.; Monath, T.P.; Lang, J. Preclinical and Clinical Development of YFV 17D-Based Chimeric Vaccines Against Dengue, West Nile and Japanese Encephalitis Viruses. Vaccine 2010, 28, 632–649. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Vaughan, K.; Weiskopf, D.; Grifoni, A.; Diamond, M.S.; Sette, A.; Peters, B. Identifying Candidate Targets of Immune Responses in Zika Virus Based on Homology to Epitopes in Other Flavivirus Species. PLoS Curr. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Grant, A.; Ponia, S.S.; Tripathi, S.; Balasubramaniam, V.; Miorin, L.; Sourisseau, M.; Schwarz, M.C.; Sánchez-Seco, M.P.; Evans, M.J.; Best, S.M.; et al. Zika Virus Targets Human STAT2 to Inhibit Type I Interferon Signaling. Cell Host Microbe 2016, 19, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Sirohi, D.; Chen, Z.; Sun, L.; Klose, T.; Pierson, T.C.; Rossmann, M.G.; Kuhn, R.J. The 3.8 A Resolution Cryo-EM Structure of Zika Virus. Science 2016, 352, 467–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostyuchenko, V.A.; Lim, E.X.Y.; Zhang, S.; Fibriansah, G.; Ng, T.-S.; Ooi, J.S.G.; Shi, J.; Lok, S.-M. Structure of the Thermally Stable Zika Virus. Nature 2016, 533, 425–428. [Google Scholar] [CrossRef] [Green Version]
- Sevvana, M.; Long, F.; Miller, A.S.; Klose, T.; Buda, G.; Sun, L.; Kuhn, R.J.; Rossmann, M.G. Refinement and Analysis of the Mature Zika Virus Cryo-EM Structure at 3.1 Å Resolution. Structure 2018, 26, 1169–1177. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Song, J.; Lu, X.; Deng, Y.-Q.; Musyoki, A.M.; Cheng, H.; Zhang, Y.; Yuan, Y.; Song, H.; Haywood, J.; et al. Structures of the Zika Virus Envelope Protein and Its Complex with a Flavivirus Broadly Protective Antibody. Cell Host Microbe 2016, 19, 696–704. [Google Scholar] [CrossRef]
- Hasan, S.S.; Sevvana, M.; Kuhn, R.J.; Rossmann, M.G. Structural Biology of Zika Virus and Other Flaviviruses. Nat. Struct. Mol. Biol. 2018, 25, 13–20. [Google Scholar] [CrossRef]
- Collins, M.H.; Tu, H.A.; Gimblet-Ochieng, C.; Liou, G.A.; Jadi, R.S.; Metz, S.W.; Thomas, A.; McElvany, B.D.; Davidson, E.; Doranz, B.J.; et al. Human Antibody Response to Zika Targets Type-Specific Quaternary Structure Epitopes. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Andrade, P.; Gimblet-Ochieng, C.; Modirian, F.; Collins, M.; Cardenas, M.; Katzelnick, L.C.; Montoya, M.; Michlmayr, D.; Kuan, G.; Balmaseda, A.; et al. Impact of Pre-Existing Dengue Immunity on Human Antibody and Memory B Cell Responses to Zika. Nat. Commun. 2019, 10, 938. [Google Scholar] [CrossRef] [Green Version]
- Dussupt, V.; Sankhala, R.S.; Gromowski, G.D.; Donofrio, G.; De La Barrera, R.A.; Larocca, R.A.; Zaky, W.; Mendez-Rivera, L.; Choe, M.; Davidson, E.; et al. Potent Zika and Dengue Cross-Neutralizing Antibodies Induced by Zika Vaccination in a Dengue-Experienced Donor. Nat. Med. 2020, 26. [Google Scholar] [CrossRef] [PubMed]
- El Sahly, H.M.; Gorchakov, R.; Lai, L.; Natrajan, M.S.; Patel, S.M.; Atmar, R.L.; Keitel, W.A.; Hoft, D.F.; Barrett, J.; Bailey, J.; et al. Clinical, Virologic, and Immunologic Characteristics of Zika Virus Infection in a Cohort of US Patients: Prolonged RNA Detection in Whole Blood. Open Forum Infect. Dis. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Edupuganti, S.; Natrajan, M.S.; Rouphael, N.; Lai, L.; Xu, Y.; Feldhammer, M.; Hill, C.; Patel, S.M.; Johnson, S.J.; Bower, M.; et al. Biphasic Zika Illness with Rash and Joint Pain. Open Forum Infect. Dis. 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waggoner, J.J.; Rouphael, N.; Xu, Y.; Natrajan, M.; Lai, L.; Patel, S.M.; Levit, R.D.; Edupuganti, S.; Mulligan, M.J. Pericarditis Associated with Acute Zika Virus Infection in a Returning Traveler. Open Forum Infect. Dis. 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, L.; Rouphael, N.; Xu, Y.; Natrajan, M.S.; Beck, A.; Hart, M.; Feldhammer, M.; Feldpausch, A.; Hill, C.; Wu, H.; et al. Innate, T-, and B-Cell Responses in Acute Human Zika Patients. Clin. Infect. Dis. 2018, 66, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravichandran, S.; Hahn, M.; Belaunzarán-Zamudio, P.F.; Ramos-Castañeda, J.; Nájera-Cancino, G.; Caballero-Sosa, S.; Navarro-Fuentes, K.R.; Ruiz-Palacios, G.; Golding, H.; Beigel, J.H.; et al. Differential Human Antibody Repertoires Following Zika Infection and the Implications for Serodiagnostics and Disease Outcome. Nat. Commun. 2019, 10, 1943. [Google Scholar] [CrossRef]
- Wold, W.S.M.; Toth, K. Adenovirus Vectors for Gene Therapy, Vaccination and Cancer Gene Therapy. Curr. Gene Ther. 2013, 13, 421–433. [Google Scholar] [CrossRef]
- Junjhon, J.; Lausumpao, M.; Supasa, S.; Noisakran, S.; Songjaeng, A.; Saraithong, P.; Chaichoun, K.; Utaipat, U.; Keelapang, P.; Kanjanahaluethai, A.; et al. Differential Modulation of prM Cleavage, Extracellular Particle Distribution, and Virus Infectivity by Conserved Residues at Nonfurin Consensus Positions of the Dengue Virus pr-M Junction. J. Virol. 2008, 82, 10776–10791. [Google Scholar] [CrossRef] [Green Version]
- Brien, J.D.; Hassert, M.; Stone, E.T.; Geerling, E.; Cruz-Orengo, L.; Pinto, A.K. Isolation and Quantification of Zika Virus from Multiple Organs in a Mouse. J. Vis. Exp. 2019, 150. [Google Scholar] [CrossRef] [Green Version]
- Hatcher, E.L.; Zhdanov, S.A.; Bao, Y.; Blinkova, O.; Nawrocki, E.P.; Ostapchuck, Y.; Schaffer, A.A.; Brister, J.R. Virus Variation Resource - Improved Response to Emergent Viral Outbreaks. Nucleic Acids Res. 2017, 45, D482–D490. [Google Scholar] [CrossRef]
- Pickett, B.E.; Sadat, E.L.; Zhang, Y.; Noronha, J.M.; Squires, R.B.; Hunt, V.; Liu, M.; Kumar, S.; Zaremba, S.; Gu, Z.; et al. ViPR: An Open Bioinformatics Database and Analysis Resource for Virology Research. Nucleic Acids Res. 2012, 40, D593–D598. [Google Scholar] [CrossRef] [PubMed]
- Turner, T. Plot Protein: Visualization of Mutations. J. Clin. Bioinform. 2013, 3, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miravet, S.; Ontiveros, M.; Piedra, J.; Penalva, C.; Monfar, M.; Chillon, M. Construction, Production, and Purification of Recombinant Adenovirus Vectors. Methods Mol. Biol. 2014, 1089, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Brien, J.D.; Lazear, H.M.; Diamond, M.S. Propagation, Quantification, Detection, and Storage of West Nile Virus. Curr. Protoc. Microbiol. 2013, 31, 15D 13 11–15D 13 18. [Google Scholar] [CrossRef] [PubMed]
- Henchal, E.A.; Gentry, M.K.; McCown, J.M.; Brandt, W.E. Dengue Virus-Specific and Flavivirus Group Determinants Identified with Monoclonal Antibodies by Indirect Immunofluorescence. Am. J. Trop. Med. Hyg. 1982, 31, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Croyle, M.A.; Patel, A.; Tran, K.N.; Gray, M.; Zhang, Y.; Strong, J.E.; Feldmann, H.; Kobinger, G.P. Nasal Delivery of an Adenovirus-Based Vaccine Bypasses Pre-Existing Immunity to the Vaccine Carrier and Improves the Immune Response in Mice. PLoS ONE 2008, 3, e3548. [Google Scholar] [CrossRef]
- Vazquez, S.; Guzman, M.G.; Guillen, G.; Chinea, G.; Perez, A.B.; Pupo, M.; Rodriguez, R.; Reyes, O.; Garay, H.E.; Delgado, I.; et al. Immune Response to Synthetic Peptides of Dengue prM Protein. Vaccine 2002, 20, 1823–1830. [Google Scholar] [CrossRef]
- Moreno, G.K.; Newman, C.M.; Koenig, M.R.; Mohns, M.S.; Weiler, A.M.; Rybarczyk, S.; Weisgrau, K.L.; Vosler, L.J.; Pomplun, N.; Schultz-Darken, N.; et al. Long-Term Protection of Rhesus Macaques from Zika Virus Reinfection. J. Virol. 2019, 20, 1–19. [Google Scholar] [CrossRef]
- Nazerai, L.; Scholler, A.S.; Rasmussen, P.O.S.; Buus, S.; Stryhn, A.; Christensen, J.P.; Thomsen, A.R. A New In Vivo Model to Study Protective Immunity to Zika Virus Infection in Mice With Intact Type I Interferon Signaling. Front. Immunol. 2018, 9, 593. [Google Scholar] [CrossRef]
- Roby, J.A.; Setoh, Y.X.; Hall, R.A.; Khromykh, A.A. Post-Translational Regulation and Modifications of Flavivirus Structural Proteins. J. Gen. Virol. 2015, 96, 1551–1569. [Google Scholar] [CrossRef]
- Allison, S.L.; Stadler, K.; Mandl, C.W.; Kunz, C.; Heinz, F.X. Synthesis and Secretion of Recombinant Tick-Borne Encephalitis Virus Protein E in Soluble and Particulate Form. J. Virol. 1995, 69, 5816–5820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanciotti, R.S.; Lambert, A.J.; Holodniy, M.; Saavedra, S.; Signor Ldel, C. Phylogeny of Zika Virus in Western Hemisphere, 2015. Emerg. Infect. Dis. 2016, 22, 933–935. [Google Scholar] [CrossRef] [PubMed]
- Metsky, H.C.; Matranga, C.B.; Wohl, S.; Schaffner, S.F.; Freije, C.A.; Winnicki, S.M.; West, K.; Qu, J.; Baniecki, M.L.; Gladden-Young, A.; et al. Zika Virus Evolution and Spread in the Americas. Nature 2017, 546, 411–415. [Google Scholar] [CrossRef] [Green Version]
- Bullard, B.L.; Corder, B.N.; Gorman, M.J.; Diamond, M.S.; Weaver, E.A. Efficacy of a T Cell-Biased Adenovirus Vector as a Zika Virus Vaccine. Sci. Rep. 2018, 8, 18017. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Costa-Ramos, P.; Pham, J.; Tian, Y.; Rosales, S.L.; Seumois, G.; Sidney, J.; de Silva, A.D.; Premkumar, L.; Collins, M.H.; et al. Cutting Edge: Transcriptional Profiling Reveals Multifunctional and Cytotoxic Antiviral Responses of Zika Virus-Specific CD8(+) T Cells. J. Immunol. 2018, 201, 3487–3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricciardi, M.J.; Magnani, D.M.; Grifoni, A.; Kwon, Y.C.; Gutman, M.J.; Grubaugh, N.D.; Gangavarapu, K.; Sharkey, M.; Silveira, C.G.T.; Bailey, V.K.; et al. Ontogeny of the B- and T-Cell Response in a Primary Zika Virus Infection of a Dengue-Naive Individual during the 2016 Outbreak in Miami, FL. PLoS Negl. Trop. Dis. 2017, 11, e0006000. [Google Scholar] [CrossRef] [PubMed]
- Emanuel, J.; Callison, J.; Dowd, K.A.; Pierson, T.C.; Feldmann, H.; Marzi, A. A VSV-Based Zika Virus Vaccine Protects Mice from Lethal Challenge. Sci. Rep. 2018, 8, 11043. [Google Scholar] [CrossRef]
- Muthumani, K.; Griffin, B.D.; Agarwal, S.; Kudchodkar, S.B.; Reuschel, E.L.; Choi, H.; Kraynyak, K.A.; Duperret, E.K.; Keaton, A.A.; Chung, C.; et al. In Vivo Protection Against ZIKV Infection and Pathogenesis through Passive Antibody Transfer and Active Immunisation with a prMEnv DNA Vaccine. NPJ Vaccines 2016, 1, 16021. [Google Scholar] [CrossRef]
- Koblischke, M.; Stiasny, K.; Aberle, S.W.; Malafa, S.; Tsouchnikas, G.; Schwaiger, J.; Kundi, M.; Heinz, F.X.; Aberle, J.H. Structural Influence on the Dominance of Virus-Specific CD4 T Cell Epitopes in Zika Virus Infection. Front. Immunol. 2018, 9, 1196. [Google Scholar] [CrossRef]
- Grifoni, A.; Pham, J.; Sidney, J.; O’Rourke, P.H.; Paul, S.; Peters, B.; Martini, S.R.; de Silva, A.D.; Ricciardi, M.J.; Magnani, D.M.; et al. Prior Dengue Virus Exposure Shapes T Cell Immunity to Zika Virus in Humans. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Metz, S.W.; Thomas, A.; Brackbill, A.; Forsberg, J.; Miley, M.J.; Lopez, C.A.; Lazear, H.M.; Tian, S.; de Silva, A.M. Oligomeric State of the ZIKV E Protein Defines Protective Immune Responses. Nat. Commun. 2019, 10, 4606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, M.A.; Bevan, M.J. Effector and Memory CTL Differentiation. Annu. Rev. Immunol. 2007, 25, 171–192. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://clinicaltrials.gov/ (accessed on 1 July 2020).
- Shiver, J.W.; Fu, T.M.; Chen, L.; Casimiro, D.R.; Davies, M.E.; Evans, R.K.; Zhang, Z.Q.; Simon, A.J.; Trigona, W.L.; Dubey, S.A.; et al. Replication-Incompetent Adenoviral Vaccine Vector Elicits Effective Anti-Immunodeficiency-Virus Immunity. Nature 2002, 415, 331–335. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steffen, T.; Hassert, M.; Hoft, S.G.; Stone, E.T.; Zhang, J.; Geerling, E.; Grimberg, B.T.; Roberts, M.S.; Pinto, A.K.; Brien, J.D. Immunogenicity and Efficacy of a Recombinant Human Adenovirus Type 5 Vaccine against Zika Virus. Vaccines 2020, 8, 170. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8020170
Steffen T, Hassert M, Hoft SG, Stone ET, Zhang J, Geerling E, Grimberg BT, Roberts MS, Pinto AK, Brien JD. Immunogenicity and Efficacy of a Recombinant Human Adenovirus Type 5 Vaccine against Zika Virus. Vaccines. 2020; 8(2):170. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8020170
Chicago/Turabian StyleSteffen, Tara, Mariah Hassert, Stella G. Hoft, E. Taylor Stone, Jianfeng Zhang, Elizabeth Geerling, Brian T. Grimberg, M. Scot Roberts, Amelia K. Pinto, and James D. Brien. 2020. "Immunogenicity and Efficacy of a Recombinant Human Adenovirus Type 5 Vaccine against Zika Virus" Vaccines 8, no. 2: 170. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8020170