
Oral Administration of Coronavirus Spike Protein Provides Protection to Newborn Pigs When Challenged with PEDV


Abstract
:1. Introduction
2. Materials and Methods
2.1. Production of Maize Expressing S1 Antigen and Preparation of the Corn Material
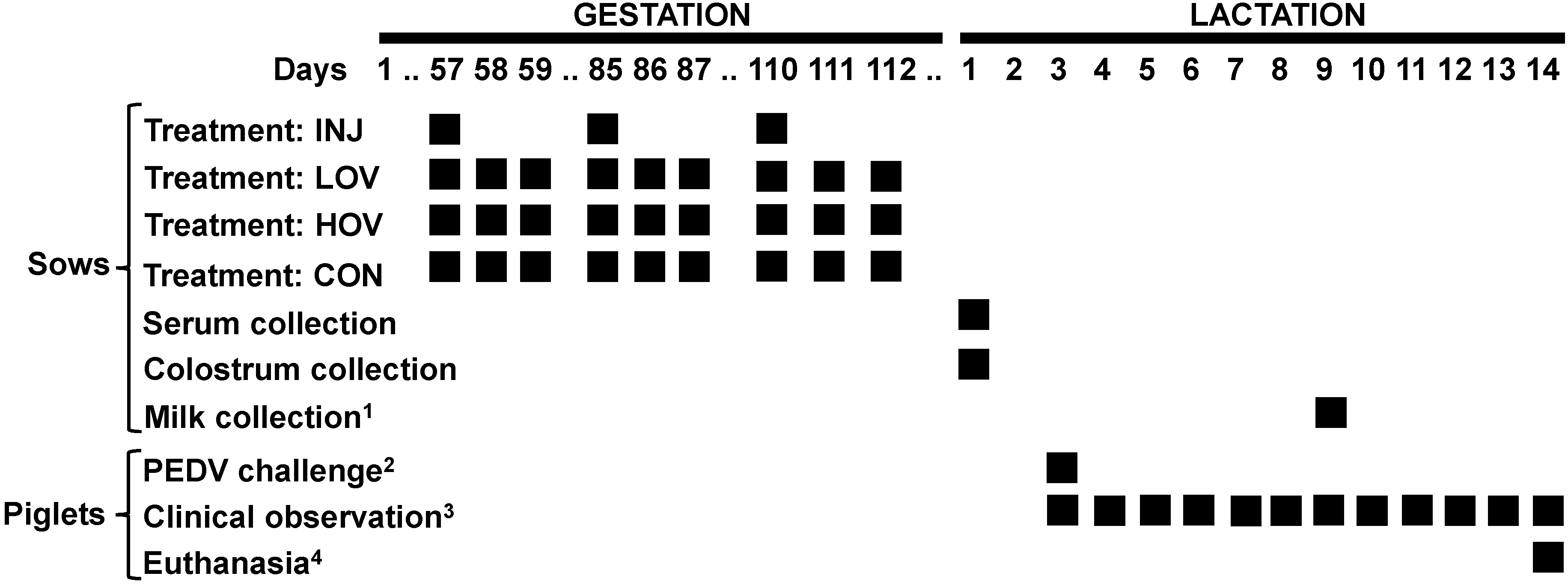
2.2. Animals and Experimental Design
2.3. Neutralizing Antibodies and Cytokines in Serum and Milk
2.4. Statistical Analyses
3. Results
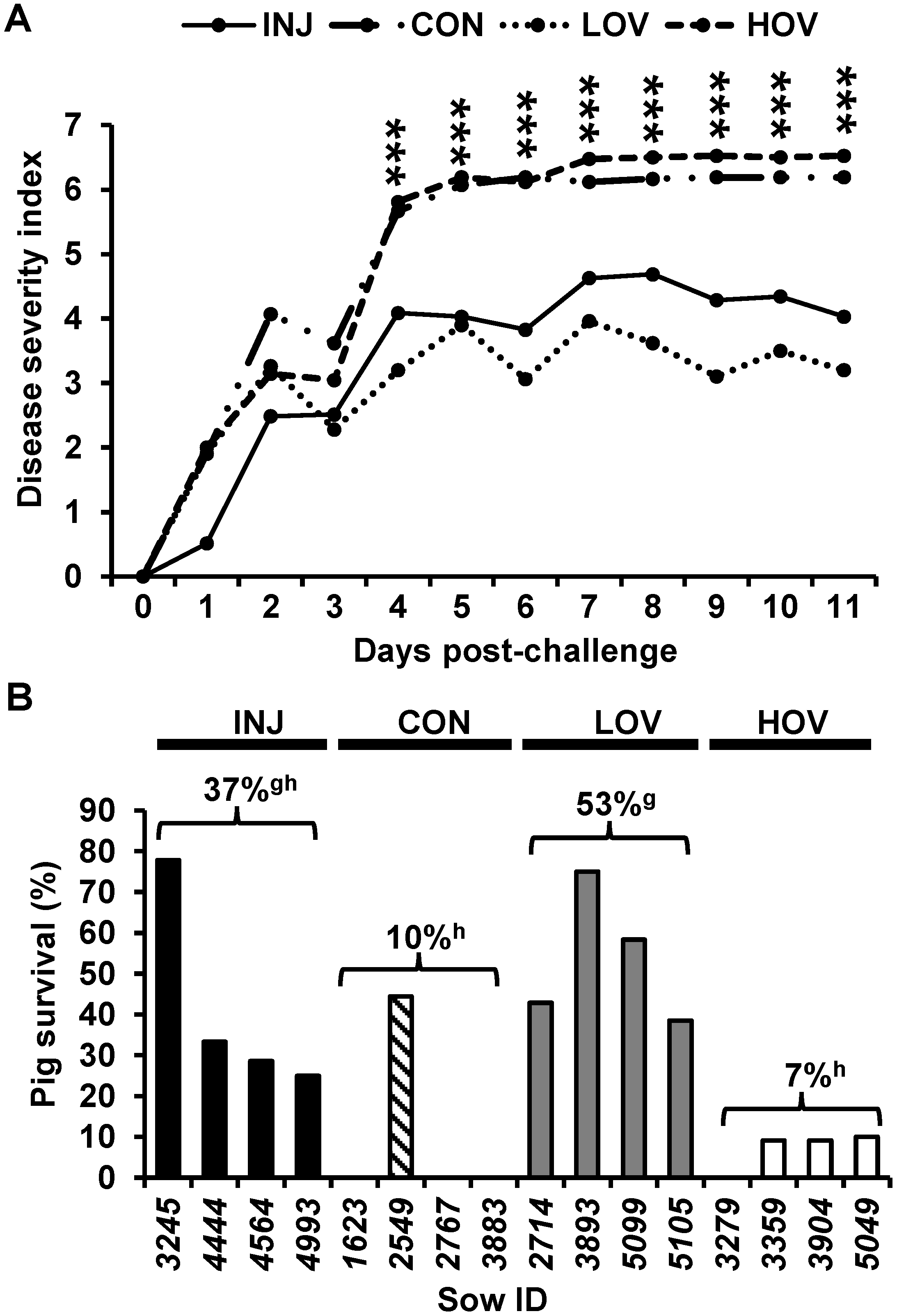
3.1. Vaccination of Sows with the S Antigen Protects Nursing Piglets against Challenge with PED Virus
3.2. Vaccination of Sows with S1 Antigen Elicits Neutralizing Antibodies Response in Serum and Milk
3.3. Administration of S1 Antigen Effects the Level of Cytokines in Sows’ Sera and Milk
3.4. NAB and GM-CSF in Milk Are Correlated with Protection in Piglets Challenged with PEDV
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Gerdts, V.; Zakhartchouk, A. Vaccines for porcine epidemic diarrhea virus and other swine coronaviruses. Vet. Microbiol. 2017, 206, 45–51. [Google Scholar] [CrossRef]
- Gao, Q.; Zheng, Z.; Wang, H.; Yi, S.; Zhang, G.; Gong, L. The New Porcine Epidemic Diarrhea Virus Outbreak May Mean That Existing Commercial Vaccines Are Not Enough to Fully Protect Against the Epidemic Strains. Front. Vet. Sci. 2021, 8, 697839. [Google Scholar] [CrossRef]
- Saif, L.J.; Wang, Q.; Vlasova, A.N.; Jung, K.; Xiao, S. Coronaviruses. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2019; Chapter 31; pp. 488–523. [Google Scholar]
- Coussement, W.; Ducatelle, R.; Debouck, P.; Hoorens, J. Pathology of experimental cv777 coronavirus enteritis in piglets.1. Histological and histochemical-study. Vet. Pathol. 1982, 19, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Machado, G.; Vilalta, C.; Recamonde-Mendoza, M.; Corzo, C.; Torremorell, M.; Perez, A.; VanderWaal, K. Identifying outbreaks of Porcine Epidemic Diarrhea virus through animal movements and spatial neighborhoods. Sci. Rep. 2019, 9, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C. Porcine epidemic diarrhea virus: An emerging and re-emerging epizootic swine virus. Virology 2015, 12, 193, Erratum in: Virology 2016, 13, 19. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Lee, K.-W.; Choi, H.-W.; Lee, C. Immunogenicity and protective efficacy of recombinant S1 domain of the porcine epidemic diarrhea virus spike protein. Arch. Virol. 2014, 159, 2977–2987. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, K.; Perlman, S. Coronaviruses, Including Severe Acute Respiratory Syndrome (SARS) and Middle East Respiratory Syndrome (MERS). Mandell Douglas Bennetts Princ. Pract. Infect. Dis. 2015, 1928–1936.e2. [Google Scholar] [CrossRef]
- Reguera, J.; Santiago, C.; Mudgal, G.; Ordono, D.; Enjuanes, L.; Casasnovas, J.M. Structural bases of coronavirus attachment to host aminopeptidase N and its inhibition by neutralizing antibodies. PLoS Pathog. 2012, 8, e1002859. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Feng, L.; Shi, H.; Chen, J.; Cui, X.; Chen, H.; Liu, S.; Tong, Y.; Wang, Y.; Tong, G. Identification of two novel B cell epitopes on porcine epidemic diarrhea virus spike protein. Vet. Microbiol. 2008, 131, 73–81. [Google Scholar] [CrossRef]
- Makadiya, N.; Brownlie, R.; van den Hurk, J.; Berube, N.; Allan, B.; Gerdts, V.; Zakhartchouk, A. S1 domain of the porcine epidemic diarrhea virus spike protein as a vaccine antigen. Virol. J. 2016, 13, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okda, F.A.; Lawson, S.; Singrey, A.; Nelson, J.; Hain, K.S.; Joshi, L.R.; Christopher-Hennings, J.; Nelson, E.A.; Diel, D.G. The S2 glycoprotein subunit of porcine epidemic diarrhea virus contains immunodominant neutralizing epitopes. Virology 2017, 509, 185–194. [Google Scholar] [CrossRef]
- Chang, S.-H.; Bae, J.-L.; Kang, T.-J.; Kim, J.; Chung, G.-H.; Lim, C.-W.; Laude, H.; Yang, M.-S.; Jang, Y.-S. Identification of the epitope region capable of inducing neutralizing antibodies against the porcine epidemic diarrhea virus. Mol. Cells 2002, 14, 295–299. [Google Scholar]
- Piao, D.-C.; Lee, Y.-S.; Bok, J.-D.; Cho, C.-S.; Hong, Z.-S.; Kang, S.-K.; Choi, Y.-J. Production of soluble truncated spike protein of porcine epidemic diarrhea virus from inclusion bodies of Escherichia coli through refolding. Protein Expr. Purif. 2016, 126, 77–83. [Google Scholar] [CrossRef]
- Van Noi, N.; Chung, Y.-C. Optimization of expression and purification of recombinant S1 domain of the porcine epidemic diarrhea virus spike (PEDV-S1) protein in Escherichia coli. Biotechnol. Biotechnol. Equip. 2017, 31, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, L.; Mou, C.; Zhang, E.; Wang, Y.; Cao, Y.; Yang, Q. Mucosal immune responses induced by oral administration recombinant Bacillus subtilis expressing the COE antigen of PEDV in newborn piglets. Biosci. Rep. 2019, 39, BSR20182028. [Google Scholar] [CrossRef] [Green Version]
- Mota, R.M.; Moreira, J.L.; Souza, M.R.; Horta, M.F.; Teixeira, S.M.; Neumann, E.; Nicoli, J.R.; Nunes, A.C. Genetic transformation of novel isolates of chicken Lactobacillus bearing probiotic features for expression of heterologous proteins: A tool to develop live oral vaccines. BMC Biotechnol. 2006, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Di-qiu, L.; Jun-wei, G.; Xin-yuan, Q.; Yan-ping, J.; Song-mei, L.; Yi-jing, L. High-level mucosal and systemic immune responses induced by oral administration with Lactobacillus-expressed porcine epidemic diarrhea virus (PEDV) S1 region combined with Lactobacillus-expressed N protein. Appl. Microbiol. Biotechnol. 2012, 93, 2437–2446. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Jiang, X.; Jiang, Y.; Tang, L.; Xu, Y.; Qiao, X.; Liu, M.; Cui, W.; Ma, G.; Li, Y. Oral Immunization against PEDV with Recombinant Lactobacillus casei Expressing Dendritic Cell-Targeting Peptide Fusing COE Protein of PEDV in Piglets. Viruses 2018, 10, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, T.-J.; Kim, Y.-S.; Jang, Y.-S.; Yang, M.-S. Expression of the synthetic neutralizing epitope gene of porcine epidemic diarrhea virus in tobacco plants without nicotine. Vaccine 2005, 23, 2294–2297. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.-L.; Lee, J.-G.; Kang, T.-J.; Jang, H.-S.; Jang, Y.-S.; Yang, M.-S. Induction of antigen-specific systemic and mucosal immune responses by feeding animals transgenic plants expressing the antigen. Vaccine 2003, 21, 4052–4058. [Google Scholar] [CrossRef]
- Huy, N.-X.; Kim, M.-Y. Improved expression of porcine epidemic diarrhea antigen by fusion with cholera toxin B subunit and chloroplast transformation in Nicotiana tabacum. Plant Cell Tissue Organ. Cult. 2019, 137, 213–223. [Google Scholar]
- Huy, N.-X.; Kim, S.-H.; Yang, M.-S.; Kim, T.-G. Immunogenicity of a neutralizing epitope from porcine epidemic diarrhea virus: M cell targeting ligand fusion protein expressed in transgenic rice calli. Plant Cell Rep. 2012, 31, 1933–1942. [Google Scholar] [CrossRef] [PubMed]
- Hood, E.E.; Howard, J.A. Over-expression of Novel Proteins in Maize. In Molecular Genetic Approaches to Maize Improvement. Biotechnology in Agriculture and Forestry; Kriz, A.L., Larkins, B.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 63. [Google Scholar]
- Streatfield, S.J.; Jilka, J.M.; Hood, E.E.; Turner, D.D.; Bailey, M.R.; Mayor, J.M.; Woodard, S.L.; Beifuss, K.K.; Horn, M.E.; Delaney, D.E.; et al. Plant-based vaccines: Unique advantages. Vaccine 2001, 19, 2742–2748. [Google Scholar] [CrossRef]
- Modelska, A.; Dietzschold, B.; Sleysh, N.; Fu, Z.F.; Steplewski, K.; Hooper, D.C.; Koprowski, H.; Yusibov, V. Immunization against rabies with plant-derived antigen. Proc. Natl. Acad. Sci. USA 1998, 95, 2481–2485. [Google Scholar] [CrossRef] [Green Version]
- Hood, E.; Kusnadi, Z.; Nikolov, Z.; Howard, J.A. Molecular farming of industrial proteins from transgenic maize. Adv. Exp. Med. Biol. 1999, 464, 127–147. [Google Scholar]
- Lamphear, B.J.; Streatfield, S.J.; Jilka, J.M.; Brooks, C.A.; Barker, D.K.; Turner, D.D.; Delaney, D.E.; Garcia, M.; Wiggins, B.; Woodard, S.L.; et al. Delivery of subunit vaccines in maize seed. J. Control Release 2002, 85, 169–180. [Google Scholar] [CrossRef]
- Lamphear, B.J.; Jilka, J.M.; Kesl, L.; Welter, M.; Howard, J.A.; Streatfield, S.J. A corn-based delivery system for animal vaccines: An oral transmissible gastroenteritis virus vaccine boosts lactogenic immunity in swine. Vaccine 2004, 22, 2420–2424. [Google Scholar] [CrossRef]
- Hayden, C.A.; Fischer, M.E.; Andrews, B.L.; Chilton, H.C.; Turner, D.D.; Walker, J.H.; Tizard, I.R.; Howard, J.A. Oral delivery of wafers made from HBsAg-expressing maize germ induces long-term immunological systemic and mucosal responses. Vaccine 2015, 33, 2881–2886. [Google Scholar] [CrossRef] [Green Version]
- Tacket, C.O. Plant-Based Oral Vaccines: Results of Human Trials. In Current Topics in Microbiology and Immunology; Karasev, A.V., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 332. [Google Scholar]
- Man, K.; Yang, B.; Xue, J.; Qin, X.; Zhou, H.; Xu, F.; Xu, C.; Yang, F.; Zhang, L.; Li, X. Expression of core neutralizing epitope gene of porcine epidemic diarrhea virus in maize. J. Agric. Sci. Technol. 2014, 16, 28–35. [Google Scholar]
- Egelkrout, E.; Hayden, C.; Fake, G.; Keener, T.; Arruda, P.; Saltzman, R.; Walker, J.; Howard, J. Oral delivery of maize-produced porcine epidemic diarrhea virus spike protein elicits neutralizing antibodies in pigs. Plant Cell Tissue Organ. Cult. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Sarmento, L.V.; Poonsuk, K.; Tian, L.; Mora-Diaz, J.C.; Main, R.G.; Baum, D.H.; Zimmerman, J.J.; Gimenez-Lirola, L.G. Detection of porcine epidemic diarrhear virus-neutralizing antibody using high-throughput imaging cytometry. J. Vet. Diagn. Investig. 2020, 32, 324–328. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 11 September 2021).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.; Buerkner, P.; Herve, M.; Love, J.; Riebl, J.; Singmann, H. Emmeans: Estimated Marginal Means. R Package Version 1.7. 2021. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 11 September 2021).
- Lüdecke, D.; Ben-Shachar, M.B.; Patil, I.; Waggoner, P.; Makowski, D. Performance: An R package for Assessment, Comparision and Testing of Statistical Models. J. Open Source Softw. 2021, 6, 3139. Available online: https://CRAN.R-project.org/package=performance (accessed on 11 September 2021). [CrossRef]
- Clement, T.; Singrey, A.; Lawson, S.; Okda, F.; Nelson, J.; Diel, D.; Nelson, E.A.; Christopher-Hennings, J. Measurement of neutralizing antibodies against porcine epidemic diarrhea virus in sow serum, colostrum, and milk samples and in piglet serum samples after feedback. J. Swine Health Prod. 2016, 24, 147–153. [Google Scholar]
- Li, C.; Li, W.; Lucio de Esesarte, E.; Guo, H.; van den Elzen, P.; Aarts, E.; van den Born, E.; Rottier, P.J.M.; Bosch, B.-J. Cell attachment domains of the porcine epidemic diarrhea virus spike protein are key targets of neutralizing antibodies. J. Virol. 2017, 91, e00273-17. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.; Stone, S.; Drebes, D.; Greiner, L.L.; Dvorak, C.M.T.; Murtaugh, M.P. Characterization of anti-porcine epidemic diarrhea virus neutralizing activity in mammary secretions. Virus Res. 2016, 226, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansariniya, H.; Seifati, S.M.; Zaker, E.; Zare, F. Comparison of Immune Response between SARS, MERS, and COVID-19 Infection, Perspective on Vaccine Design and Development. BioMed Res. Int. 2021, 2021, 8870425. [Google Scholar] [CrossRef]
- Curtis, J.; Bourne, F.J. Immunoglobulin quantitation in sow serum, colostrum and milk and the serum of young pigs. Biochim. Biophys. Acta Protein Struct. 1971, 236, 319–332. [Google Scholar] [CrossRef]
- Boyaka, P.N. Inducing mucosal IgA: A challenge for vaccine adjuvants and delivery systems. J. Immunol. 2017, 199, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Markowska-Daniel, I.; Pomorska-Mol, M.; Pejsak, Z. Dynamic changes of immunoglobulin concentrations in pig colostrum and serum around parturition. Pol. J. Vet. Sci. 2010, 13, 21–27. [Google Scholar]
- Langel, S.N.; Wang, Q.; Vlasova, A.N.; Saif, L.J. Host Factors Affeting Generation of Immunity Against Porcine Epidemic Diaarhea Virus in Pregnant and Lactating Swine and Passive Protection of Neonates. Pathogenes 2020, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.V.; Yuan, L.; Azevedo, M.S.; Jeong, K.I.; Gonzalez, A.M.; Saif, L.J. Transfer of maternal cytokines to suckling piglets: In vivo and in vitro models with implications for immunomodulation of neonatal immunity. Vet. Immunol. Immunopathol. 2007, 117, 236–248. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Doan, Q.C.; Regester, G.O. Detection and characterization of transforming growth factor-beta in porcine colostrum. Biol. Neonate 1999, 75, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.C.; Achuthan, A.A.; Hamilton, J.A. GM-CSF: A Promising Target in Inflammation and Autoimmunity. Immunotargets Ther. 2020, 9, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Hansen, G.; Hercus, T.R.; McClure, B.J.; Stomski, F.C.; Dottore, M.; Powell, J.; Ramshaw, H.; Woodcock, J.M.; Xu, Y.; Guthridge, M.; et al. The structure of the GM-CSF receptor complex reveals a distinct mode of cytokine receptor activation. Cell 2008, 134, 496–507. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Probability | Groupings |
|---|---|---|
| INJ | 0.26 | ab |
| CON | 0.09 | a |
| LOV | 0.56 | b |
| HOV | 0.08 | a |
| Cytokine | INJ | CON | LOV | HOV |
|---|---|---|---|---|
| GM-CSF | 200.1 a ± 183.7 | 1128.1 b ± 547.6 | 159.3 a ± 179.1 | 381.2 ab ± 238.4 |
| IFNγ | 3903.7 ± 4063.2 | 10694.7 ± 1352.6 | 4567.7 ± 4689.0 | 5448.4 ± 5079.1 |
| IL-1α | 158.8 ± 104.6 | 472.1 ± 145.6 | 325.7 ± 186.9 | 262.2 ± 190.4 |
| IL-1β | 1624.8 ± 1095.4 | 4874.9 ± 2181.3 | 3092.4 ± 1972.9 | 3583.5 ± 3039.3 |
| IL-1RA | 938.9 ± 619.6 | 2992.1 ± 874.8 | 2290.8 ± 1451.3 | 2330.7 ± 1992.9 |
| IL-2 | 1371.6 ± 979.2 | 4386.1 ± 1417.7 | 2959.8 ± 1959.1 | 3026.7 ± 2549.3 |
| IL-4 | 4404.2 ± 3329.9 | 15752.0 ± 5540.5 | 10763.5 ± 7541.7 | 10326.9 ± 8721.3 |
| IL-6 | 468.3 ± 368.8 | 1138.9 ± 501.4 | 700.3 ± 554.9 | 1274.6 ± 1185.8 |
| IL-8 | 8710.6 ± 5239.0 | 6812.2 ± 3008.8 | 8509.7 ± 3806.0 | 8254.3 ± 1823.0 |
| IL-10 | 2135.5 ± 1565.5 | 7833.4 ± 3245.6 | 5569.7 ± 3664.9 | 4791.4 ± 4135.0 |
| IL-12 | 589.2 ± 439.1 | 1702.4 ± 535.8 | 1196.6 ± 763.0 | 1157.8 ± 968.0 |
| IL-18 | 2855.0 ± 2099.3 | 7994.6 ± 3349.0 | 5366.2 ± 3520.9 | 6667.1 ± 5481.6 |
| TNFα | 421.7 ± 501.5 | 518.1 ± 273.1 | 225.0 ± 271.3 | 703.5 ± 623.6 |
| Predictor | p-Value | Odds Ratio 95% Lower Limit | Odds Ratio 95% Upper Limit |
|---|---|---|---|
| Standardized Weight | ≤0.001 | 1.96 | 6.11 |
| Standardized Milk GM-CSF | 0.02 | 0.05 | 0.72 |
| Standardized Sera GM-CSF | 0.27 | 0.68 | 4.01 |
| Milk NABs | ≤0.001 | 3.71 | 65.89 |
| Sera NABs | 0.38 | 0.11 | 2.18 |
| Sow ID | Treatment | Milk GM-CSF (pg/mL) | Piglets Surviving (%) |
|---|---|---|---|
| 3245 | INJ | 219 | 78 |
| 4444 | INJ | 92 | 17 |
| 4564 | INJ | 489 | 28 |
| 4993 | INJ | 0 | 25 |
| 1623 | CON | 1193 | 0 |
| 2767 | CON | 1201 | 0 |
| 2549 | CON | 291 | 44 |
| 3883 | CON | 1828 | 0 |
| 2714 | LOV | 436 | 43 |
| 3893 | LOV | 199 | 75 |
| 5099 | LOV | 0 | 38 |
| 5105 | LOV | 0 | 58 |
| 3279 | HOV | 557 | 0 |
| 3359 | HOV | 608 | 9 |
| 3904 | HOV | 0 | 9 |
| 5049 | HOV | 358 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maj, M.; Fake, G.M.; Walker, J.H.; Saltzman, R.; Howard, J.A. Oral Administration of Coronavirus Spike Protein Provides Protection to Newborn Pigs When Challenged with PEDV. Vaccines 2021, 9, 1416. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9121416
Maj M, Fake GM, Walker JH, Saltzman R, Howard JA. Oral Administration of Coronavirus Spike Protein Provides Protection to Newborn Pigs When Challenged with PEDV. Vaccines. 2021; 9(12):1416. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9121416
Chicago/Turabian StyleMaj, Magdalena, Gina M. Fake, John H. Walker, Ryan Saltzman, and John A. Howard. 2021. "Oral Administration of Coronavirus Spike Protein Provides Protection to Newborn Pigs When Challenged with PEDV" Vaccines 9, no. 12: 1416. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9121416