1. Introduction
Infectious canine hepatitis (ICH) is an acute septic infectious disease, and its etiological agent is canine adenovirus type 1 (CAdV-1). Hepatitis, corneal oedema (blue eye), jaundice and anemia are the main clinical symptoms. Some studies showed that CAdV-1 continues to circulate in carnivores [
1,
2,
3]. CAdV type 2 (CAdV-2) can cause infectious tracheobronchitis (ITB) and pneumonia in dogs, also known as kennel cough, fever, serous to mucinous rhinorrhea, tonsillitis and laryngotracheitis in dogs, which has a high prevalence in canine populations [
4,
5].
CadVs have non-enveloped capsids with pseudo-icosahedral symmetry, linear double-stranded DNA of about 30 kb and are included in the genus
Mastadenovirus of the family
Adenoviridae [
6]. The genome of CAdV includes 30 open reading frames (ORFs) flanked by two identical 161 bp inverted terminal repeats [
7]. The major coat protein of CAdV is hexon (Hex), which plays a major role in viral tropism and neutralization [
8]. The N-terminal domain (with a core size ~484 amino acids [aa]) and C-terminal domain (core size ~221 aa) adopt the same PNGase F-like fold, although they are significantly different in length. The CAdV capsid structure is relatively well conserved, and is primarily composed of homo-trimers of Hex; 240 trimers form the icosahedron’s 20 facets [
9,
10]. There are 12 vertex penton capsomers, each with a fiber protruding from the surface [
10,
11,
12]. The trimeric Hex has a pseudohexagonal base with three towers extending upwards [
10,
11]. Apart from these, the capsid is also stabilized by other Hex-associated proteins [
13]. It is estimated that most of the adenoviral neutralizing antibodies are generated against Hex [
14].
In this study, we characterized the potent neutralizing monoclonal antibody (mAb) 2C1, and a partially neutralizing mAb 7D7, raised against CAdV-2. We expressed and purified a truncated His-fused Hex (552–850 aa) containing most of C-terminal domain; then tested the neutralizing mAbs for recognition in Western blot (WB) assays. Using a phage display approach, we finely mapped the recognized epitopes to 634RIKQRETPAL643 and 736PESYKDRMYS745 on the CAdV-2 Hex. Since the two linear epitopes are highly conserved among CAdV-1 and CAdV-2 strains, these research results may assist in the design of structure-based novel epitopes vaccines or therapeutic vaccines and antibody therapeutics, and the development of rapid detection techniques for CAdV antigens or antibodies.
2. Materials and Methods
2.1. Virus Strains, Hybridoma Lines, Plasmids and Phage Peptide Library
The CAdV-2 strain used in this study was stored at the Harbin Veterinary Research Institute of the Chinese Academy of Agricultural Science. Hybridoma lines secreting CAdV-2-specific mAbs were generated in our lab previously [
15]. The pET-30a (+) vector was purchased from Tiangen, China. A commercially available Ph.D.-12TM Phage Display Peptide Library Kit was purchased from New England BioLabs Inc. CAdV-2 was cultured in Madin–Darby canine kidney (MDCK) cells in Dulbecco’s modified eagle’s medium with 10% fetal calf serum (Gibco, USA) at 37 °C, and
Escherichia coli ER2738 and Rosetta (DE3) were cultured in Luria–Bertani (LB) medium.
2.2. Characterization of mAbs against CAdV-2
Hybridoma lines secreting CAdV-2-specific mAbs were cultured, and the isotype was determined using the SBA Clonotyping
TM System/HRP Kit (Southern Biotech, USA). Specificity characterization of CAdV-2-specific mAbs 2C1 and 7D7 was carried out by indirect immunofluorescence assay (IFA) and WB according to standard procedures [
15], but WB using denaturing SDS-PAGE and native PAGE, respectively. Denaturing SDS-PAGE was carried out according to standard procedures. In brief, 15 μL of virus sample was mixed with 5 μL 4X SDS-PAGE loading buffer and boiled for 10 min. Samples were then loaded into 12% SDS-PAGE gel (Invitrogen), and electrophoresis was performed at room temperature using a constant voltage (120 V). In NSDS-PAGE, 15 μL of virus sample was mixed with 5 μL of 4X NSDS sample buffer (150 mM Tris base, 100 mM Tris HCl, 10% (v/v) glycerol, 0.0185% (w/v) Coomassie G-250, 0.00625% (
w/v) phenol red, pH 8.5, Invitrogen). Then, samples were loaded into the gel and electrophoresis was conducted using the chilled NSDS-PAGE running buffer (50 mM Tris Base, 50 mM MOPS, 0.0375% SDS, pH 7.3) under 4 °C at 120 V [
16].
2.3. Micro-Neutralization Assay
The micro-neutralization assay was performed according to the Office International Epizooties (OIE) manual. Briefly, ascetic fluid of 2C1or 7D7 was heat-inactivated for 30 min at 56 °C. Two-fold serial dilutions of heat-inactivated ascetic fluid of 2C1or 7D7 were mixed with an equal volume of viral solution containing 200 50% tissue culture infectious dose (TCID
50) CAdV-2, anti-CAdV-2 mouse serum as positive control and a mAb without activity as a negative control, and then mixtures were incubated for 1 h at 37 °C in 96-well microplates; afterward, 150 μL medium containing 10
4 MDCK cells was added to every well. The plates were incubated for 4 days at 37 °C in a humidified atmosphere with 5% CO
2, and we inspected cytopathic effects via an inverted optical microscope. The experiment was repeated 3 times. The Kärber method was used to calculate the 50% neutralizing titer (NT50) [
17,
18].
2.4. Immunoprecipitation and MALDI-TOF-MS
In order to identify the CAdV-2 protein that could bind to the mAbs, 1 μL Triton X-100 was added in 100 μL CAdV-2 suspension from infected cells for 1 h at 4 °C; then cell lysates from infected cells were added to pre-cleared protein A/G agarose beads for 2 h, followed by incubation with purified mAb ascite fluids of 2C1 or 7D7 overnight at 4 °C. 30 μL immunoprecipitates were suspended in sample buffer and boiled for 10 min, then centrifuged and analyzed by SDS-PAGE. Protein bands were subjected to MALDI-TOF MS analysis, and mass spectra were acquired by an Ettan MALDI-TOF Pro mass spectrometer (GE Healthcare, Uppsala, Sweden) by Sensichip Infortech Co. Ltd. (Shanghai, China).
2.5. Expression and Identification of Truncated Hex in E. coli
According to the sequence of CAdV-2 Hex (GenBank accession number AC000020.1), the primers were designed to amplify an 897-bp fragment (F: 5′-1654-TAT
GAATTCGACCTCCGAGTGGATGGG-1671-3′, R: 5′-2533-GAG
CTCGAGGGAGTTAGAGTACAGCAG—2550-3′). The fragment was cloned into a pET30a (+) vector (Tiangen, Beijing, China) to construct the expression plasmids. The expression plasmid was transformed into
E. coli BL
21 competent cells (Tiangen); the truncated Hex protein was produced via isopropyl-β-D-thiogalactopyranoside (IPTG, GE Healthcare, Chicago, IL, USA) induction using pET30a- Hex
E. coli at 37 °C. The truncated Hex protein was detected by SDS-PAGE and Western blotting (WB) analysis by anti-His mAb. Immunoreactive bands were verified by an enhanced chemiluminescence system (ECL; PerkinElmer Life Sciences, Fremont, CA, USA) [
15]. The three-dimensional (3D) structure of the expressed protein was predicted with PyMOL software based on data from the SWISS-MODEL online server.
2.6. Biopanning of Phage Random Peptide Library against Anti-CAdV-2 mAbs
A commercially Phage Display Peptide Library Kit (New England BioLabs Inc., Ipswich, MA, USA) was used to carry out four rounds of biopanning according to the manufacturer’s manual. Briefly, 10 μg purified mAb 2C1 or 7D7 was coated in a 96-well plate and incubated overnight at 4 °C. The plates were washed with blocking buffer with 1% BSA at room temperature for 2 h and then washed 7 times with Tris-Buffered Saline and Tween 20 (TBST). Next, the phages in the phage library (M13) was diluted to 2 × 10
12 PFU/mL and added to antibody-coated wells (100 μL/well), then incubated for 1 h at room temperature on the shaker. Bound phage were subsequently eluted with 100 μL of 0.2 M glycine/HCl (pH 2.2) containing 1 mg/mL BSA and the eluate was neutralized with 15 μL of 1 M Tris-HCl (pH 9.1) [
19]. Eluted phage were amplified in
E. coli (ER2738), and titrated on LB plates containing IPTG and X-Gal for the subsequent rounds of selection. The number of input phage was 10
11 phages in subsequent each round of biopanning, too. After 4 rounds of screening [
19,
20], the eluted phages were plated and individual clones were picked randomly for phage ELISA as described [
17,
21].
2.7. DNA Sequencing and Displayed Peptide Analysis
The single phages with high absorbance were chosen for sequencing. DNA of phage clones was extracted using a DNA extraction kit (Tiangen, China), and sequenced with the 96 gIII sequencing primer (Shanghai Biological Engineering Technology Co. Ltd.; Beijing, China). The deduced aa sequences from the insert sequences were analyzed by the DNASTAR software and compared with Hex (AP_000622.1) from CAdV-2. In order to confirm the minimal epitopes recognized by 2C1 and 7D7 in Hex, a series of peptides spanning 4–10 aa of Hex with a purity greater than 95% (F1-F17) was synthesized (Shanghai Biological Engineering Technology Co. Ltd.; Beijing, China) as coating antigens for peptide ELISA assays (
Table 1).
2.8. Biological Information Analysis
To explore novel Hex epitope’s conservation among different CAdV strains and other adenovirus strains, aa sequences from all CAdV reference strains available in GenBank were compared with other adenovirus strains in the NCBI database (
Table 2) using Megalign of DNASTAR software. The spatial distribution of the epitopes identified in Hex were analyzed by mapping epitope locations on a 3D model of CAdV-2 Hex (GenBank accession number AC000020.1) using PyMOL software based on results from the SWISS-MODEL online server. Hex secondary structural features were predicted using PROTEAN of DNASTAR software.
3. Results
3.1. Characterization of Two Neutralizing mAbs—2C1 and 7D7
Two mAbs, 2C1 and 7D7, against CAdV-2, were generated and classified by subtype to into the IgG1 subclass, and light chains were the κ-type (
Table 3). Furthermore, a micro-neutralization assay on MDCK was used to determine the neutralizing activities of both mAbs. The mAb 2C1 could neutralize potently CAdV-2 with an NT50 of 4096, but mAb 7D7 could neutralize partly against CAdV-2 with an NT50 of 64 (
Table 3).
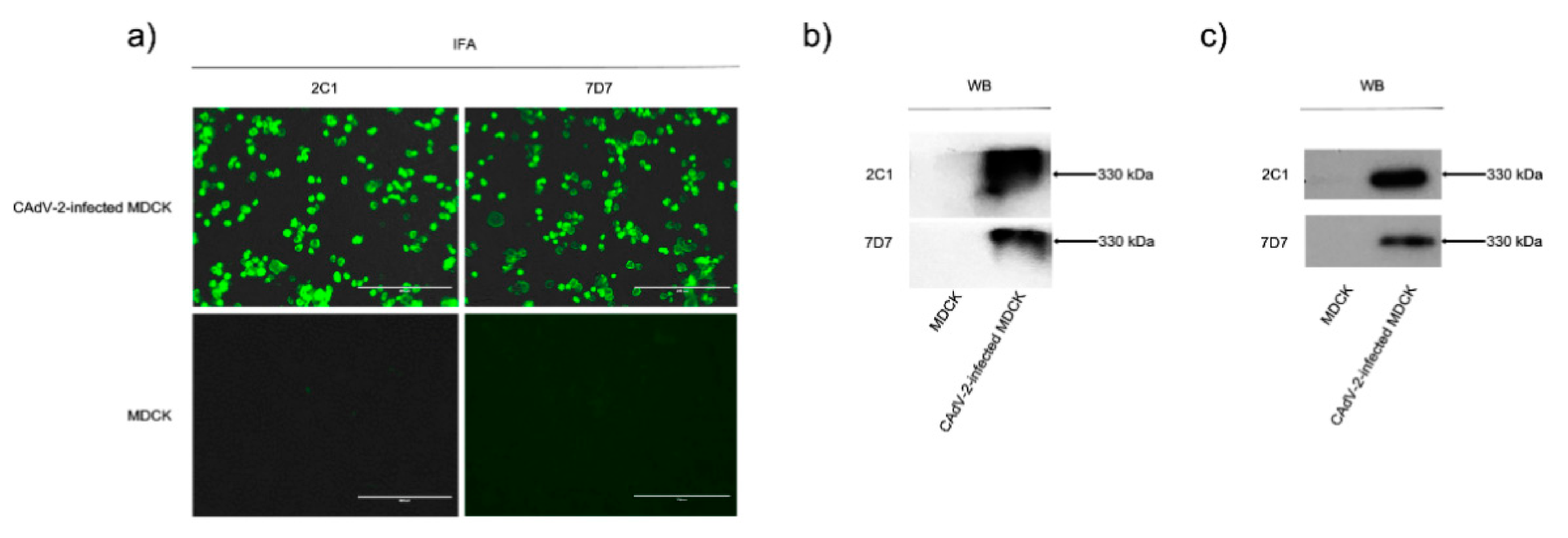
We confirmed that mAbs 2C1 and 7D7 reacted specifically with CAdV-2-infected MDCK cells in an indirect immunofluorescence assay (IFA) and native CAdV-2 protein in a WB after native PAGE (
Figure 1a,b), and also bound purified CAdV-2 antigen in ELISA (
Figure S1). Both mAbs could recognize denatured CAdV-2 proteins in WB (
Figure 1c), indicating that they are binding to linear epitopes.
3.2. MAbs 2C1 and 7D7 Specifically Recognize the CAdV-2 Hexon Protein
WB results showed that the mAbs 2C1 and 7D7 recognized a 330 kDa band after both native PAGE and SDS-PAGE of CAdV-2 (
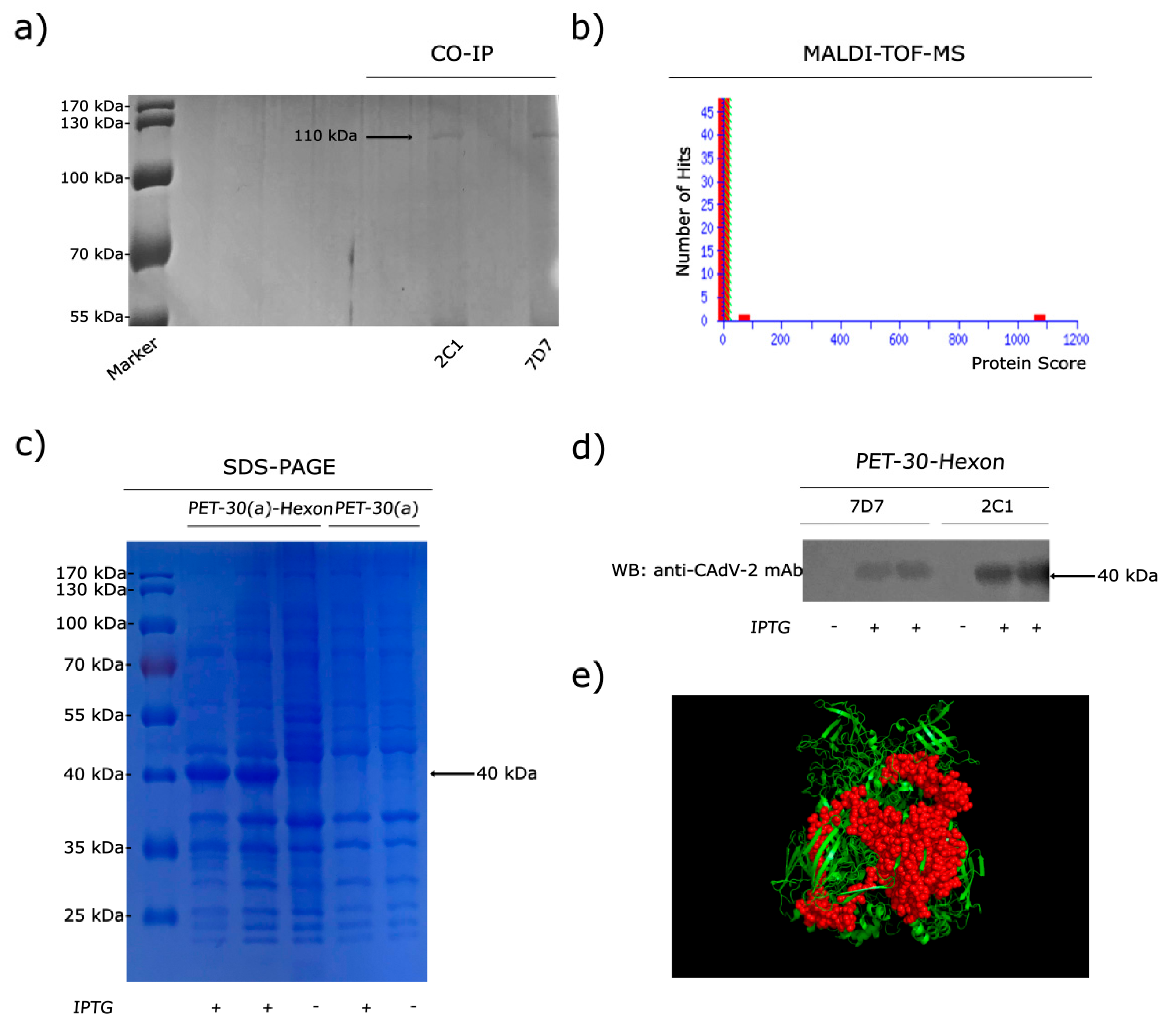
Figure 1b,c). In order to determine which protein was binding, virions were immunoprecipitated (IP) with mAbs 2C1 or 7D7. Immunoreactive protein bands were visualized in SDS-PAGE (
Figure 2a), and immunoprecipitates were analyzed through MALDI-TOF-MS based on the peptide mass matching. By searching in the NCBI database, the reactive protein (protein scores 1074 larger than 26 were significant,
p < 0.05) was identified as CAdV-1 Hex (taxon identifier: 69150), which has a molecular mass of 110 kDa (
Figure 2b). Thus, the 330 kDa protein recognized in WB corresponds to the trimeric form of Hex, which was not reduced to monomeric Hex after 2-ME treatment, suggesting that the bonds between monomers are not disulfide bonds but non-covalent bonds.
In order to define the domain of the protein binding to the mAbs, a truncated recombinant protein spanning the C-terminal 300 aa of the CAdV-2 Hex was generated, which is predominantly expressed in the form of inclusion body in
E. coli after 0.5 mM IPTG induction. SDS-PAGE analysis revealed a 40-kDa band in the original expression products (
Figure 2c). WB results showed that the truncated Hex peptide was specifically recognized by mAbs 2C1 and 7D7 (
Figure 2d). A predicted 3D structure of truncated monomeric form of the Hex peptide was generated using PyMOL software on the basis of data predicted through the online server of SWISS-MODEL (
Figure 2e).
3.3. Phage-Displayed Mimic Epitopes Biopanned by mAbs 2C1 and 7D7
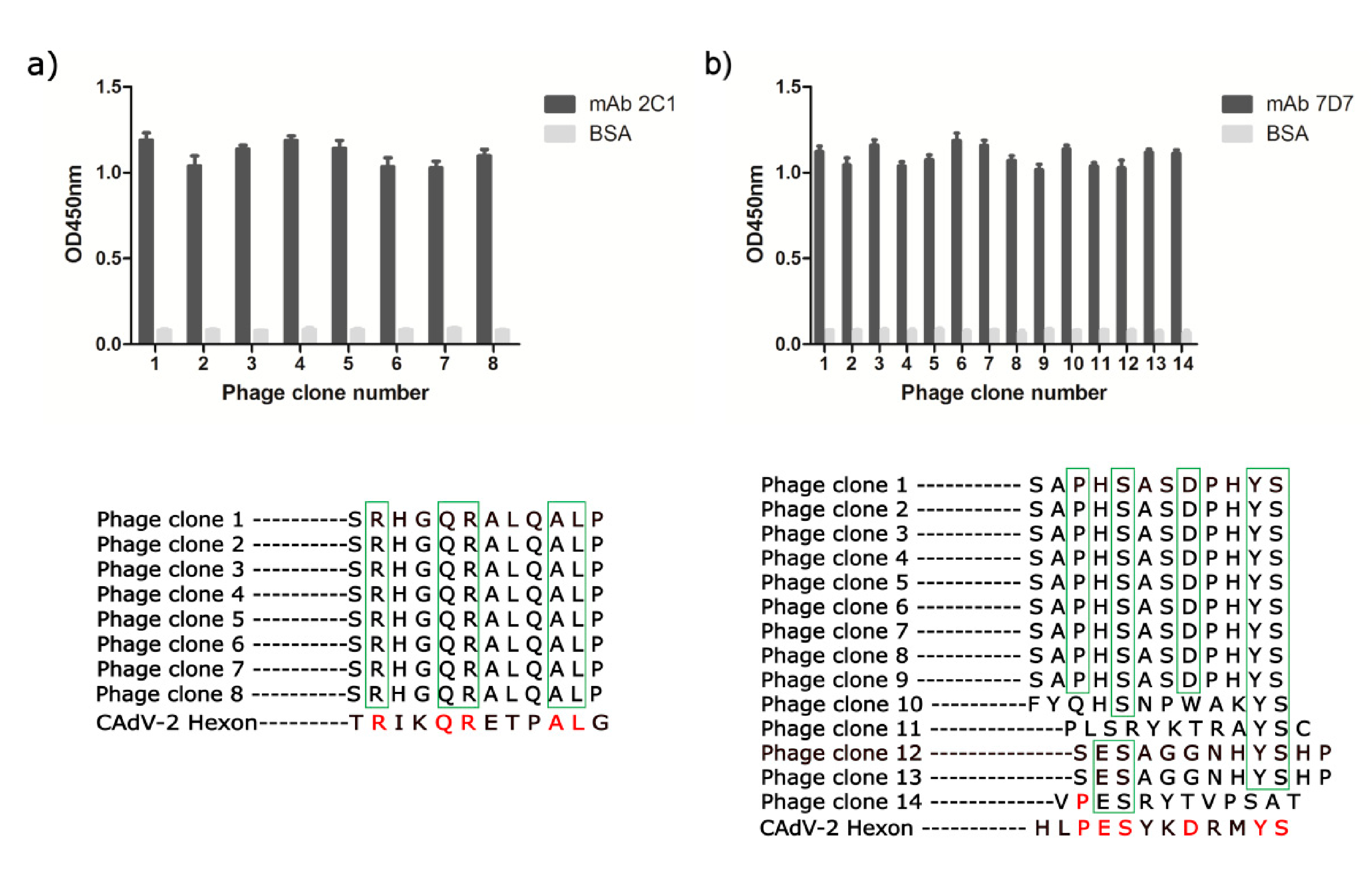
To determine the linear epitopes recognized by neutralizing mAbs 2C1 and 7D7, affinity purified mAbs 2C1 and 7D7 were used to perform phage-displayed 12-mer random peptide library biopanning. After four rounds of panning, an enrichment of phage that bound to mAbs 2C1 or 7D7 was achieved. 15 individual phage clones were isolated from the final eluted phage plate, and their reactivities to mAbs 2C1 or 7D7 were assessed by phage ELISA, with monophages exhibiting high absorbance selected for sequencing. Thus, 8 and 14 individual positive phage clones from 2C1 or 7D7, respectively, were selected for sequencing of inserts (
Figure 3). All eight positive phage clones with high affinity to mAb 2C1 displayed an identical 12-mer peptide “SRHGQRALQALP,” similar to the sequence
634RIKQRETPAL
643 observed in the CAdV-2 Hex (
Figure 3a). The high-affinity phage clones with mAb 7D7 displayed a consensus motif PESXXDXXYS (X is any aa), similar to the sequence
736PESYKDRMYS
745 observed in the CAdV-2 Hex (
Figure 3b).
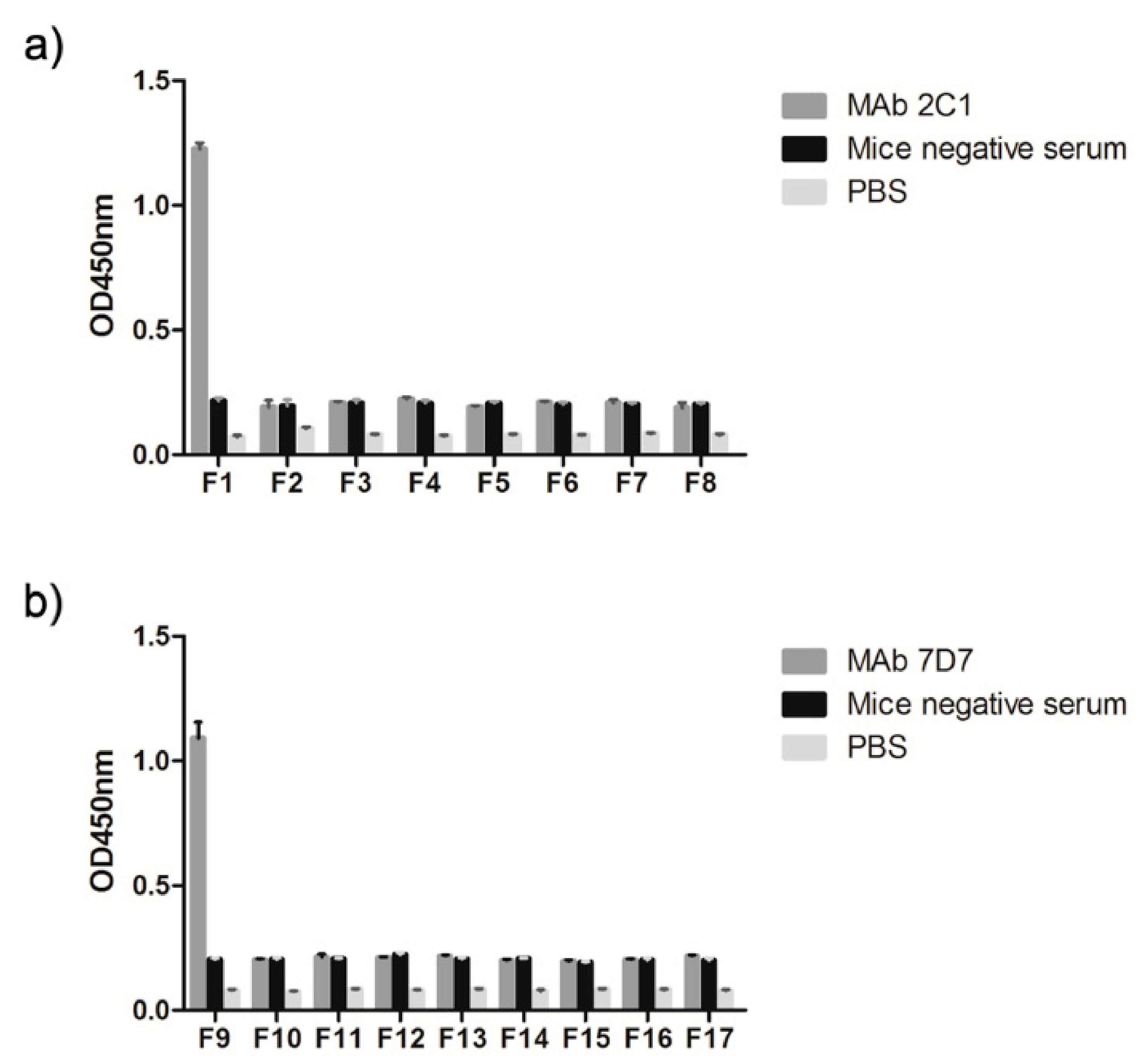
The preliminary localization of the linear neutralizing epitope recognized by 2C1 and 7D7 was verified using a panel of synthetic peptides of variable length (4–10 mers) to identify the minimal reactivity unit of each epitope. These 17 synthetic peptides as coating antigens were used to perform ELISAs, with an irrelevant protein and the recombinant 300-aa Hex fragment used as negative and positive controls, respectively. The result showed that peptides F1 and F9 were recognized by mAbs 2C1 and 7D7, respectively (
Figure 4), suggesting that the core sequences
634RIKQRETPAL
643 and
736PESYKDRMYS
745 are the minimal required sequences for antibody binding.
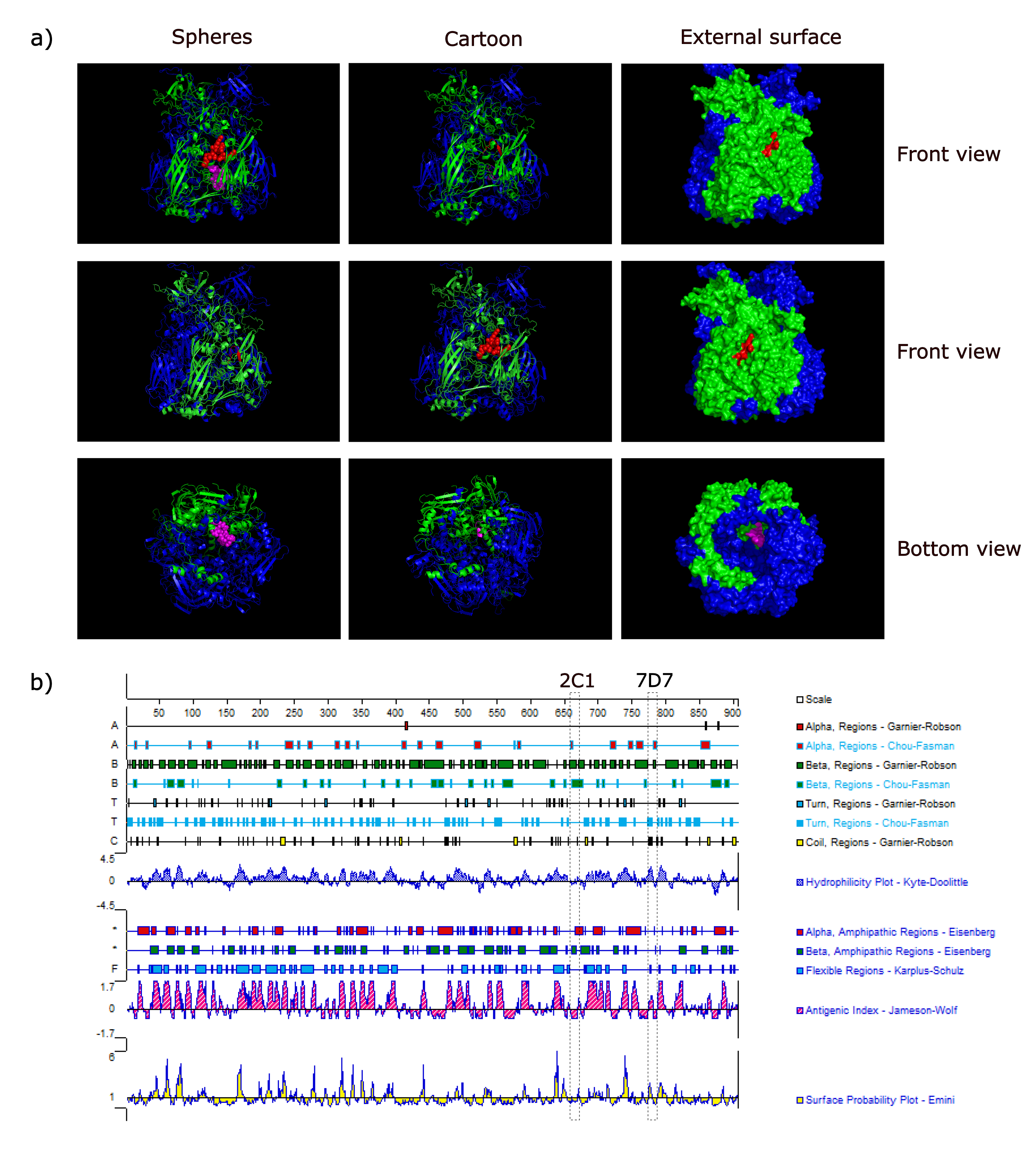
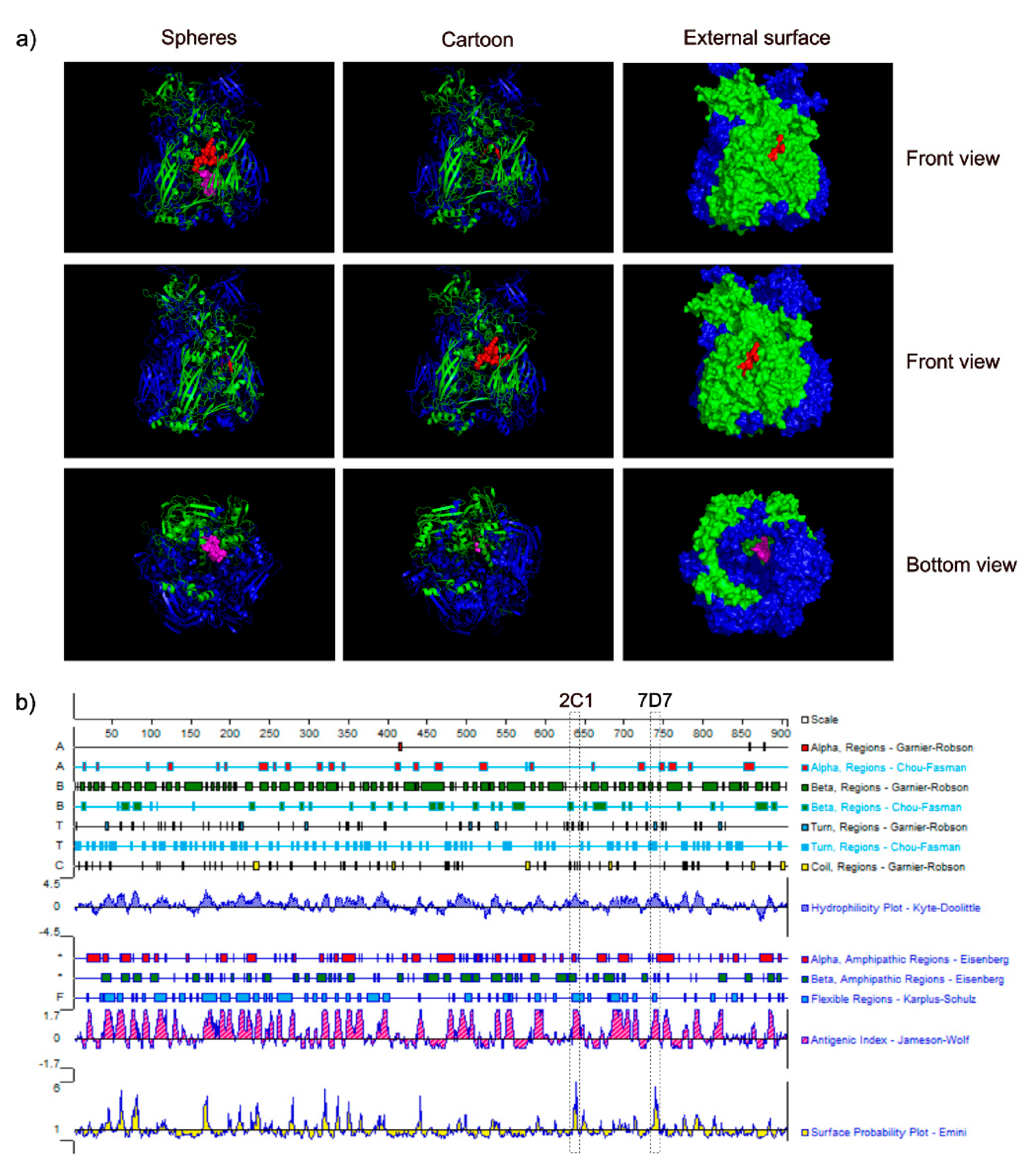
3.4. Spatial Location of the Epitopes
To understand the structure of two epitopes identified by mAbs 2C1 and 7D7, we studied the arrangement and localization of epitopes in the 3D structure of trimeric forms of Hex. This analysis revealed that the epitope of 2C1-recognized was exposed fully on the surface of the predicted CAdV-2 trimeric Hex structure, forming part of a beta-sheet (
Figure 5a). In contrast, the epitope recognized by mAb 7D7 is located within the inner region of the predicted trimeric structure as part of a beta-sheet (
Figure 5a), and is thus a cryptic epitope. Analysis using PROTEAN predicted that both epitope sequences should form part of a beta-sheet (
Figure 5b), which is the same as the predicted 3D structure shown in
Figure 5a. Furthermore, surface-exposed
634RIKQRETPAL
643 is predicted to have strong hydrophilicity (the score: 2.04) and a high antigenic index (the score: 3), which suggested that it may be an important B-cell epitope on the CAdV-2 Hex.
3.5. Conservation of Novel Epitopes in Different AdV Strains
To determine whether the two novel epitopes recognized by 2C1 or 7D7 are conserved among other AdV strains, the aa sequences of both epitopes were aligned with putative aa sequences from different AdV isolates available in GenBank. As shown in
Figure 6a, the 2C1 linear epitope
634RIKQRETPAL
643 was found in all CAdV Hex genes available in GenBank. The 7D7 epitope
736PESYKDRMYS
745 was similarly conserved among the hexon proteins of all CAdV isolates available in GenBank, though one CAdV-2 strain (ABH11656) had a single aa polymorphism (Y- > C) (
Figure 6a). However, neither epitope sequence was highly conserved in other different AdV isolates available in GenBank, except porcine mastadenovirus C had a single aa polymorphism (I- > L) in epitope
634RIKQRETPAL
643; and cynomolgus adenovirus, human adenovirus 3 and simian adenovirus 1 had a single aa polymorphism (S- > G/N) in epitope
736PESYKDRMYS
745 (
Figure 6b).
4. Discussion
Viruses display various epitopes on their outer surface that generally play roles in antibody-mediated immune responses leading to protection. Mapping these epitopes as a function of their recognition by mAbs is a powerful tool for the diagnosis of diseases [
22]. Effective protection against viral infection usually requires inducing a high level of neutralizing antibodies [
23], which can recognize specific antigenic epitopes or sites located on the surfaces of viral particles. Thus, in the development of epitope-based vaccines, the first step is the identification of antigenic epitopes recognized by neutralizing mAbs. Neutralizing antigenic sites have been identified on many viruses, including foot-and-mouth disease virus, HAdV-14 and waterfowl parvoviruses [
17,
18,
24,
25]. However, to date there is a lack of information finely mapping the neutralizing epitopes on CAdV.
In the present study, the strongly CAdV-2 neutralizing mAb 2C1 and a partially neutralizing mAb 7D7 have been characterized in vitro. The passive immunization and protection test will be further investigated in future studies to determine the actual neutralizing effect in vivo. The results of biopanning a phage-display library provided clues for us to locate the true epitopes in monomeric Hex, and synthesized peptides further revealed the precise core determinants of the mAbs 2C1 and 7D7 binding sites were 634RIKQRETPAL643 and 736PESYKDRMYS745. The two epitopes are highly conserved among CAdV isolates, but not in other adenovirus isolates available in GenBank, which all have 1–9 aa differences (most have more than three aa differences). This suggests that two mAbs agaist two epitopes could be used simultaneously to develop a diagnostic method for CAdV in dogs.
The structural model showed that the identified B-cell epitope recognized by 2C1 existed on the surface of the trimeric Hex and formed part of a beta-sheet, which is in line with PROTEAN prediction. This information suggests that the 2C1-recognized epitope is likely an important epitope, which may allow us to develop CAdV epitopes vaccines and perhaps better induce neutralizing antibodies in immunized animals. Moreover, mAb 2C1 also can be used to develop therapeutic vaccines or antibody therapeutics as a result of its good neutralization ability regarding the virus. However, the 7D7-recognized B-cell epitope is present in the inner region of the trimeric Hex, although PROTEAN software predicted that this epitope was located on the surface of monomeric Hex. Thence, mAb 7D7 cannot be used to develop therapeutic vaccines or antibody therapeutics because of its weaker neutralization ability regarding the virus.
In our study, native trimers of CAV-2 Hex from whole virions and denatured trimers of CAV-2 Hex from virus lysates could both be detected by the mAbs, which suggests the mAbs recognized two linear epitopes. The trimeric Hex was shown to be resistant to the formation of monomeric chain units by 2-ME treatment under SDS-PAGE with boiling, which is consistent with previous reports [
26]. It suggests that the connections between the monomer chains are through non-covalent bonds but not disulfide bonds.
5. Conclusions
Two novel linear neutralizing epitopes on the Hex of CAdV were identified, which provides valuable data for the design of epitope-based vaccines or therapeutic vaccines, antibody therapeutics and diagnostic markers.
Supplementary Materials
The following are available online at
https://0-www-mdpi-com.brum.beds.ac.uk/2076-393X/9/2/135/s1, Figure S1: mAbs 2C1 and 7D7 reacted specifically with purified CAdV-2 antigen in ELISA. The purified CAdV-2 was coated with 100 ng/well in the ELISA plate, mAbs 2C1; 7D7 ascetic fluid (1:10,000) was the first antibody; HRP-labeled goat anti-mouse IgG (1:10,000) was the second antibody; PBS was the negative control and mouse anti- CAdV-2 serum was the positive control for ELISA.
Author Contributions
S.W., C.W. and X.C. conceived the study and designed the experimental procedures. S.W., X.R., W.X., H.W. and Y.Z. performed the experiments. S.W., X.R., W.X., H.H., G.W. and Y.Z. analyzed the data. S.W., C.W., H.W., H.H. and G.W. contributed reagents and materials. S.W., C.W. and X.C. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key Research and Development Program (2017YFD0500600) of China and the Key Program of Natural Science Foundation of Heilongjiang Province (ZD2017007).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by The Committee on the Ethics of Animal Experiments of the Harbin Veterinary Research Institute of the Chinese Academy of Agricultural Sciences (approval number SY-2016-MI-013).
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Balboni, A.; Verin, R.; Morandi, F.; Poli, A.; Prosperi, S.; Battilani, M. Molecular epidemiology of canine adenovirus type 1 and type 2 in free-ranging red foxes (Vulpes vulpes) in Italy. Vet. Microbiol. 2013, 162, 551–557. [Google Scholar] [CrossRef]
- Balboni, A.; Mollace, C.; Giunti, M.; Dondi, F.; Prosperi, S.; Battilani, M. Investigation of the presence of canine ade-novirus (CAdV) in owned dogs in Northern Italy. Res. Vet. Sci. 2014, 97, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.; Fee, S.A.; Hartley, G.; Learmount, J.; O’Hagan, M.J.H.; Meredith, A.L.; Bronsvoort, B.M.D.C.; Porphyre, T.; Sharp, C.P.; Philbey, A.W. Serological and molecular epidemiology of canine adenovirus type 1 in red foxes (Vulpes vulpes) in the United Kingdom. Sci. Rep. 2016, 6, 36051. [Google Scholar] [CrossRef] [Green Version]
- Headley, S.; Michelazzo, M.D.M.Z.; Elias, B.; Viana, N.E.; Pereira, Y.L.; Pretto-Giordano, L.G.; Da Silva, J.F.; Da Silva, F.E.S.; Vilas-Boas, L.A.; Flaiban, K.K.M.D.C.; et al. Disseminated melanized fungal infection due to Cladosporium halotolerans in a dog coinfected with canine adenovirus-1 and canine parvovirus-2. Braz. J. Microbiol. 2019, 50, 859–870. [Google Scholar] [CrossRef]
- Sowman, H.R.; Cave, N.J.; Dunowska, M. A survey of canine respiratory pathogens in New Zealand dogs. New Zealand Vet. J. 2018, 66, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Bulut, O.; Yapici, O.; Avci, O.; Simsek, A.; Atli, K.; Dik, I.; Yavru, S.; Hasircioglu, S.; Kale, M.; Mamak, N. The Serological and Virological Investigation of Canine Adenovirus Infection on the Dogs. Sci. World J. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibanes, S.; Kremer, E.J. Canine adenovirus type 2 vector generation via I-Sce1-mediated intracellular genome re-lease. PLoS ONE 2013, 8, e71032. [Google Scholar] [CrossRef] [PubMed]
- Solanki, M.; Zhang, W.; Jing, L.; Ehrhardt, A. Adenovirus hexon modifications influence in vitro properties of pseudotyped human adenovirus type 5 vectors. J. Gen. Virol. 2016, 97, 160–168. [Google Scholar] [CrossRef]
- Stewart, P.L.; Burnett, R.M.; Cyrklaff, M.; Fuller, S.D. Image reconstruction reveals the complex molecular organiza-tion of adenovirus. Cell 1991, 67, 145–154. [Google Scholar] [CrossRef]
- Schoehn, G.; El Bakkouri, M.; Fabry, C.M.S.; Billet, O.; Estrozi, L.F.; Le, L.; Curiel, D.T.; Kajava, A.V.; Ruigrok, R.W.H.; Kremer, E.J. Three-Dimensional Structure of Canine Adenovirus Serotype 2 Capsid. J. Virol. 2008, 82, 3192–3203. [Google Scholar] [CrossRef] [Green Version]
- Rux, J.J.; Kuser, P.R.; Burnett, R.M. Structural and Phylogenetic Analysis of Adenovirus Hexons by Use of High-Resolution X-Ray Crystallographic, Molecular Modeling, and Sequence-Based Methods. J. Virol. 2003, 77, 9553–9566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Raaij, M.J.; Mitraki, A.; Lavigne, G.; Cusack, S. A triple beta-spiral in the adenovirus fibre shaft reveals a new structural motif for a fibrous protein. Nature 1999, 401, 935–938. [Google Scholar] [CrossRef] [PubMed]
- Saban, S.D.; Silvestry, M.; Nemerow, G.R.; Stewart, P.L. Visualization of alpha-helices in a 6-angstrom resolution cryoelectron microscopy structure of adenovirus allows refinement of capsid protein assignments. J. Virol. 2006, 80, 12049–12059. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.M.; Nanda, A.; Havenga, M.J.; Abbink, P.; Lynch, D.M.; Ewald, B.A.; Liu, J.; Thorner, A.R.; Swanson, P.E.; Gorgone, D.A.; et al. Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-existing anti-vector immun-ity. Nature 2006, 441, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wen, Y.; An, T.; Duan, G.; Sun, M.; Ge, J.; Li, X.; Yang, K.; Cai, X. Development of an Immunochromato-graphic Strip for Rapid Detection of Canine Adenovirus. Front. Microbiol. 2019, 10, 2882. [Google Scholar] [CrossRef]
- Nowakowski, A.B.; Wobig, W.J.; Petering, D.H. Native SDS-PAGE: High resolution electrophoretic separation of proteins with retention of native properties including bound metal ions. Met. Integr. Biometal Sci. 2014, 6, 1068–1078. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhao, L.; Li, W.; Zhou, G.; Yu, L. Identification of a conformational epitope on the VP1 G-H Loop of type Asia1 foot-and-mouth disease virus defined by a protective monoclonal antibody. Vet. Microbiol. 2011, 148, 189–199. [Google Scholar] [CrossRef]
- Liu, W.; Yang, B.; Wang, M.; Wang, H.; Yang, D.; Ma, W.; Zhou, G.; Yu, L. Identification of a conformational neutral-izing epitope on the VP1 protein of type A foot-and-mouth disease virus. Res. Vet. Sci. 2017, 115, 374–381. [Google Scholar] [CrossRef]
- Zheng, N.; Chai, Z.; Fu, F.; Jiang, F.; Wang, X.; Zhang, X.; Wang, Z.; Li, X. Identification of a Novel Haemophilus parasuis-Specific B Cell Epitope Using Monoclonal Antibody against the OppA Protein. PLoS ONE 2014, 9, e84516. [Google Scholar] [CrossRef]
- Xiao, N.; Cao, J.; Zhou, H.; Ding, S.Q.; Kong, L.Y.; Li, J.N. Identification of three novel B-cell epitopes of VMH pro-tein from Vibrio mimicus by screening a phage display peptide library. Vet. Immunol. Immunopathol. 2016, 182, 22–28. [Google Scholar] [CrossRef]
- Xue, M.; Shi, X.; Zhang, J.; Zhao, Y.; Cui, H.; Hu, S.; Gao, H.; Cui, X.; Wang, Y. Identification of a Conserved B-cell Epitope on Reticuloendotheliosis Virus Envelope Protein by Screening a Phage-displayed Random Peptide Library. PLoS ONE 2012, 7, e49842. [Google Scholar] [CrossRef]
- Kouzmitcheva, G.A.; Petrenko, V.A.; Smith, G.P. Identifying Diagnostic Peptides for Lyme Disease through Epitope Discovery. Clin. Diagn. Lab. Immunol. 2001, 8, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, R.; Tang, H.; Jin, M.; Chen, H.-C.; Qian, P. Induction of protective immunity in swine by immunization with live attenuated recombinant pseudorabies virus expressing the capsid precursor encoding regions of foot-and-mouth disease virus. Vaccine 2008, 26, 2714–2722. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Tian, X.; Jiang, Z.; Huang, J.; Liu, Q.; Lu, X.; Luo, Q.; Zhou, R. Neutralizing epitopes mapping of human ad-enovirus type 14 hexon. Vaccine 2015, 33, 6659–6665. [Google Scholar] [CrossRef]
- Lian, C.; Zhang, R.; Lan, J.; Yang, Y.; Li, H.; Sui, N.; Xie, Z.; Jiang, S. Identification of a common conserved neutral-izing linear B-cell epitope in the VP3 protein of waterfowl parvoviruses. Avian Pathol. 2020, 49, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Verkhovskaia, L.V.; Ulasov, V.N.; Kiseleva, E.K.; Khil’Ko, S.N.; I Tikhonenko, T. Purification and properties of the hexon antigen of canine adenovirus serotype CAV-1. Biokhimiia 1990, 55, 1996–2001. [Google Scholar] [PubMed]
| Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}