
Immunogenicity Assessment of Different Segments and Domains of Group A Streptococcal C5a Peptidase and Their Application Potential as Carrier Protein for Glycoconjugate Vaccine Development


Abstract
:
1. Introduction
2. Materials and Methods
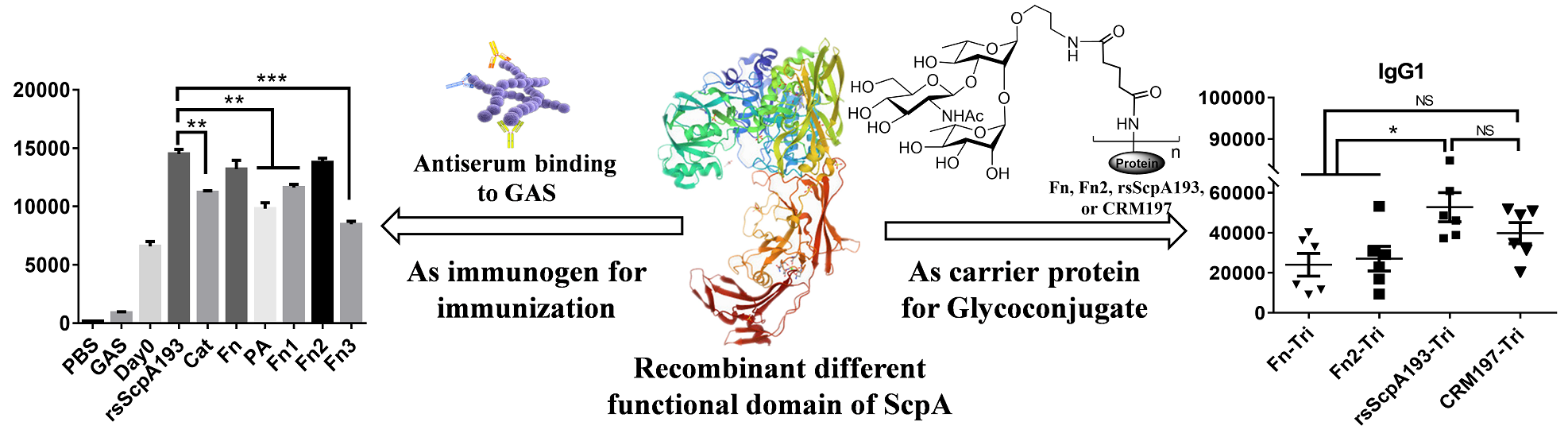
2.1. Expression and Purification of Protein Antigens
2.2. Evaluation of the Enzymatic Activities of rsScpA, rsScpA193 and Cat Proteins
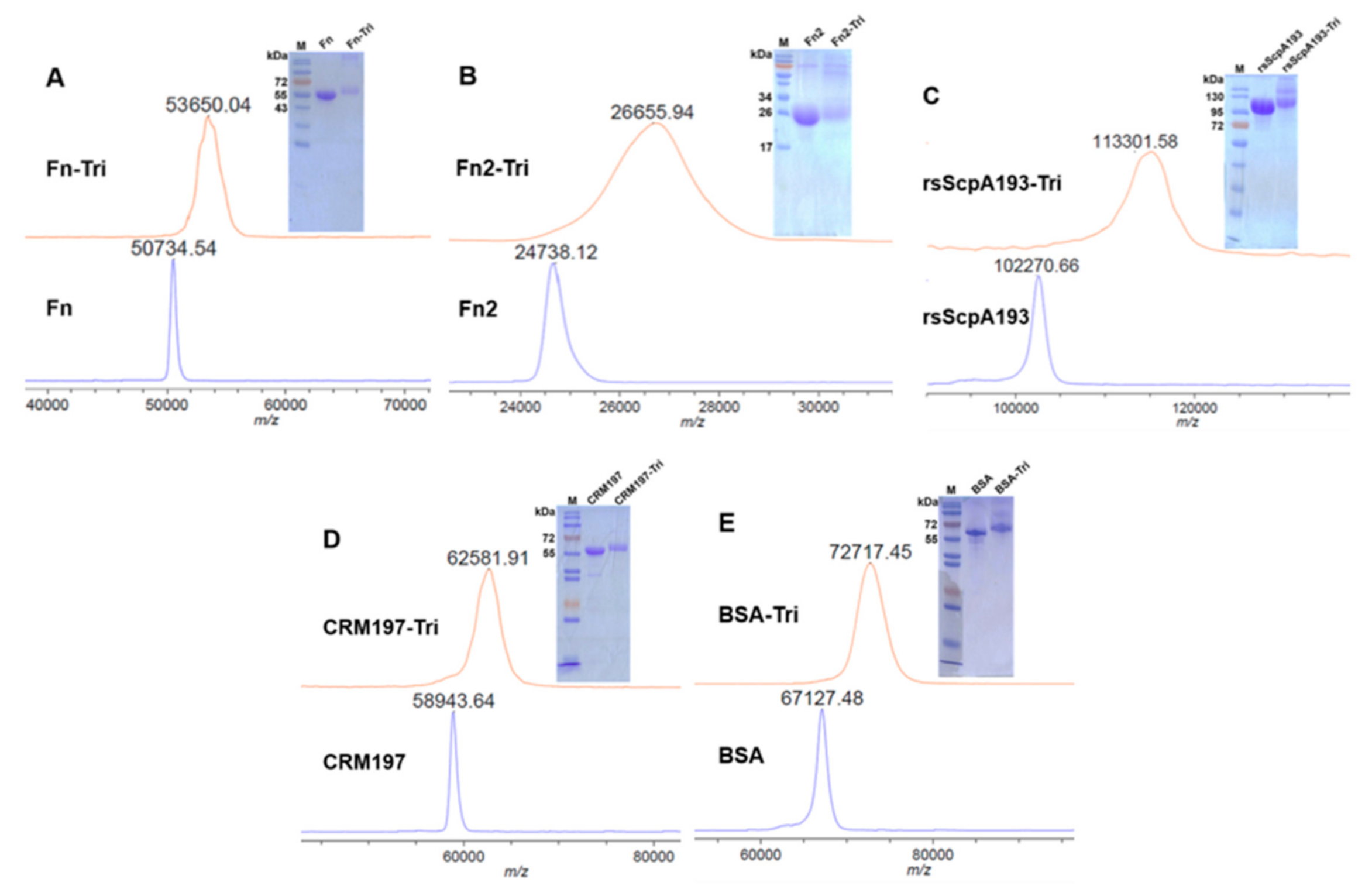
2.3. Preparation of the GAC Trisaccharide–Protein Conjugates
2.4. Immunization of Mice
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Flow Cytometry (FCM) Analysis
3. Results and Discussion
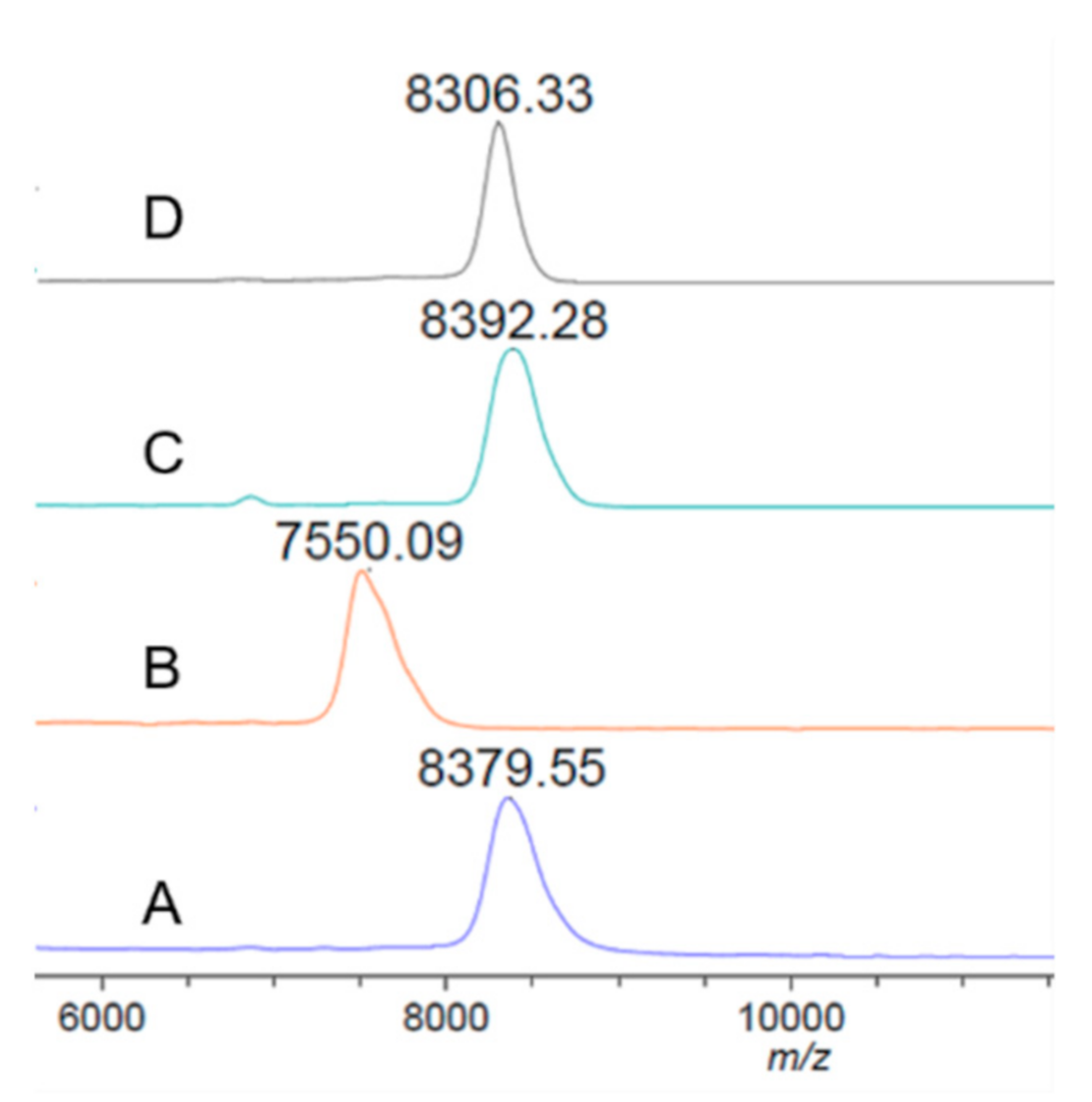
3.1. Preparation of Recombinant Target Proteins
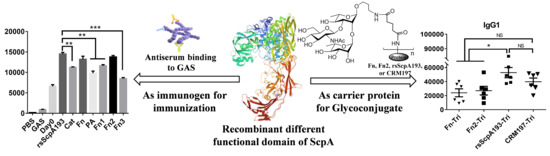
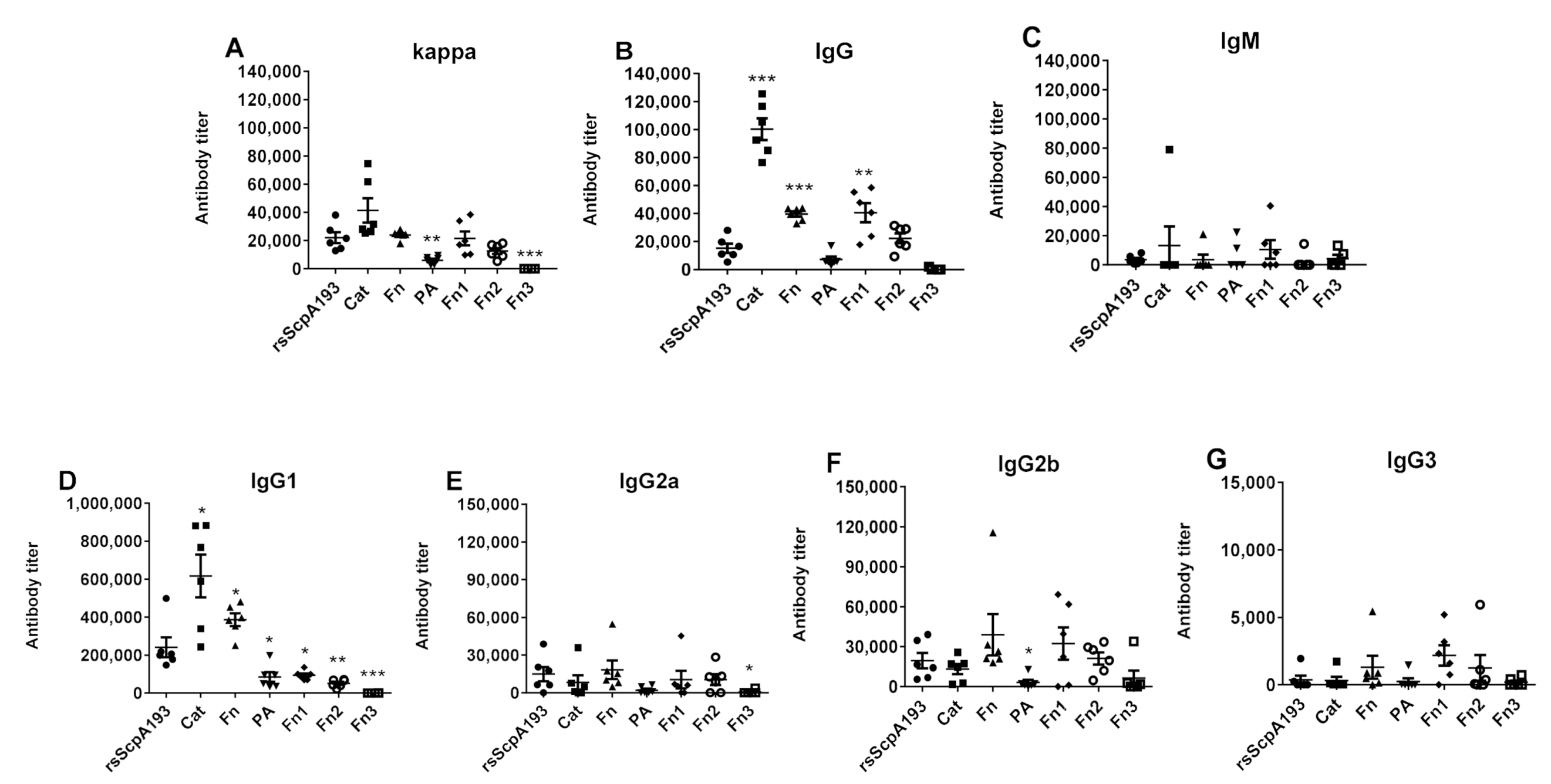
3.2. Immunological Evaluation of Segmented Domains of ScpA Protein
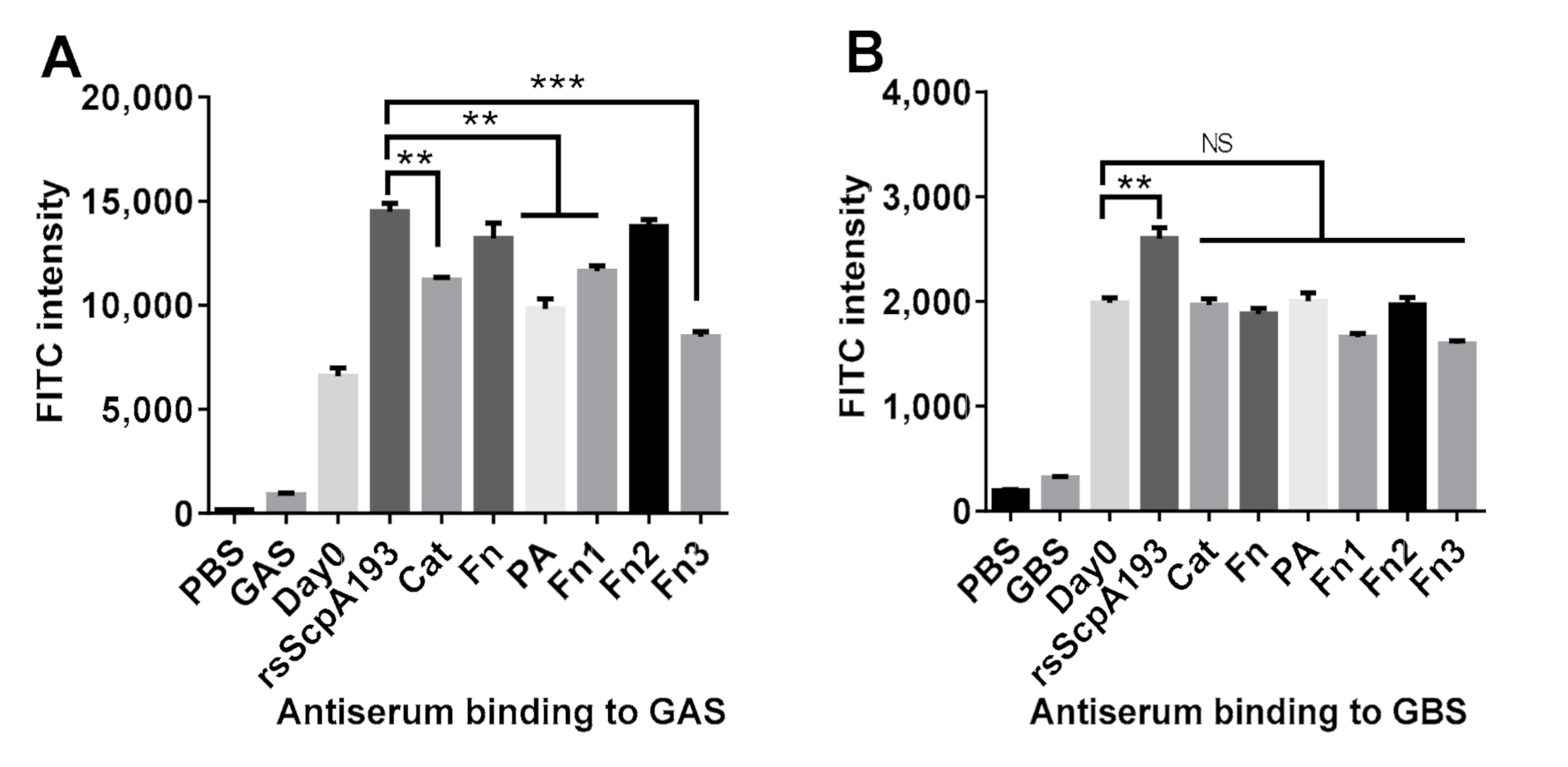
3.3. Antiserum Binding to GAS and GBS Bacterium
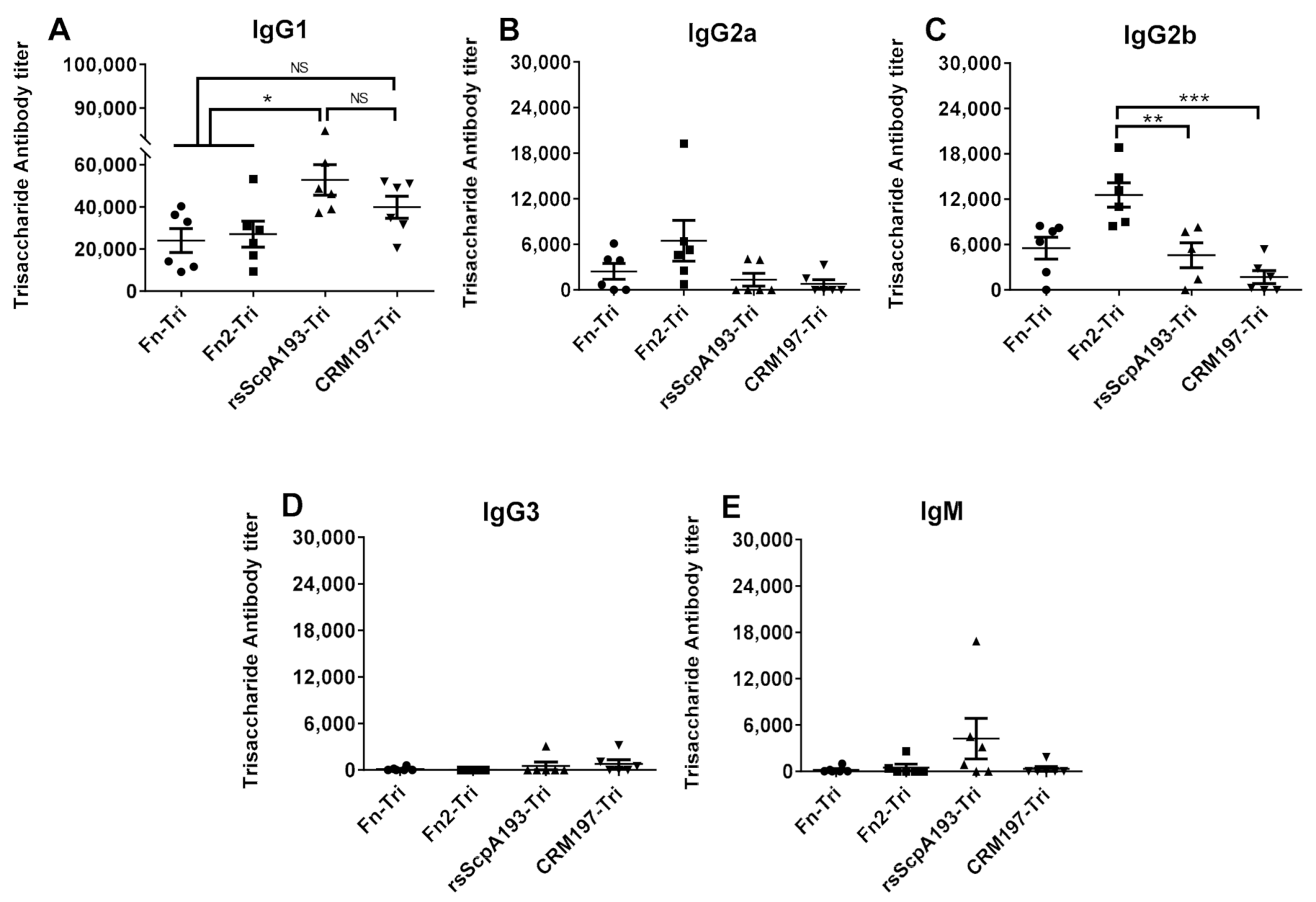
3.4. Preparation and Immunological Evaluation of GAC Trisaccharide-Protein Conjugates with Fn, Fn2 and rsScpA193 as Carrier Proteins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cunningham, M.W. Pathogenesis of group A streptococcal infections. Clin. Microbiol. Rev. 2000, 13, 470–511. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.J.; Barnett, T.C.; McArthur, J.D.; Cole, J.N.; Gillen, C.M.; Henningham, A.; Sriprakash, K.S.; Sanderson-Smith, M.L.; Nizet, V. Disease manifestations and pathogenic mechanisms of group A streptococcus. Clin. Microbiol. Rev. 2014, 27, 264–301. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, M.W. Pathogenesis of group a streptococcal infections and their sequelae. Adv. Exp. Med. Biol. 2008, 609, 29–42. [Google Scholar]
- Carapetis, J.R.; Steer, A.C.; Mulholland, E.K.; Weber, M. The global burden of group A streptococcal diseases. Lancet Infect. Dis. 2005, 5, 685–694. [Google Scholar] [CrossRef]
- Watkins, D.A.; Johnson, C.O.; Colquhoun, S.M.; Karthikeyan, G.; Beaton, A.; Bukhman, G.; Forouzanfar, M.H.; Longenecker, C.T.; Mayosi, B.M.; Mensah, G.A.; et al. Global, regional, and national burden of rheumatic heart disease, 1990–2015. N. Engl. J. Med. 2017, 377, 713–722. [Google Scholar] [CrossRef]
- McNeil, S.A.; Halperin, S.A.; Langley, J.M.; Smith, B.; Warren, A.; Sharratt, G.P.; Baxendale, D.M.; Reddish, M.A.; Hu, M.C.; Stroop, S.D.; et al. Safety and immunogenicity of 26-valent group A streptococcus vaccine in healthy adult volunteers. Clin. Infect. Dis. 2005, 41, 1114–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, J.B.; Penfound, T.A.; Chiang, E.Y.; Walton, W.J. New 30-valent M protein-based vaccine evokes cross-opsonic antibodies against non-vaccine serotypes of group A streptococci. Vaccine 2011, 29, 8175–8178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batzloff, M.R.; Hayman, W.A.; Davies, M.R.; Zeng, M.; Pruksakorn, S.; Brandt, E.R.; Good, M.F. Protection against group A streptococcus by immunization with J8-diphtheria toxoid: Contribution of J8-and diphtheria toxoid-specific antibodies to protection. J. Infect. Dis. 2003, 187, 1598–1608. [Google Scholar] [CrossRef] [Green Version]
- Guilherme, L.; Postol, E.; de Barros, S.F.; Higa, F.; Alencar, R.; Lastre, M.; Zayas, C.; Puschel, C.R.; Silva, W.R.; Sa-Rocha, L.C.; et al. Vaccine against S. pyogenes: Design and experimental immune response. Methods 2009, 49, 316–321. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Corretti, M.; Palmer, K.; Campbell, J.D.; Reddish, M.A.; Hu, M.C.; Wasserman, S.S.; Dale, J.B. Safety and immunogenicity of a recombinant multivalent group A streptococcal vaccine in healthy adults-phase 1 trial. JAMA-J. Am. Med. Assoc. 2004, 292, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Sabharwal, H.; Michon, F.; Nelson, D.; Dong, W.L.; Fuchs, K.; Manjarrez, R.C.; Sarkar, A.; Uitz, C.; Viteri-Jackson, A.; Suarez, R.S.R.; et al. Group A streptococcus (GAS) carbohydrate as an immunogen for protection against GAS infection. J. Infect. Dis. 2006, 193, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Kabanova, A.; Margarit, I.; Berti, F.; Romano, M.R.; Grandi, G.; Bensi, G.; Chiarot, E.; Proietti, D.; Swennen, E.; Cappelletti, E.; et al. Evaluation of a group A streptococcus synthetic oligosaccharide as vaccine candidate. Vaccine 2010, 29, 104–114. [Google Scholar] [CrossRef]
- Auzanneau, F.I.; Borrelli, S.; Pinto, B.M. Synthesis and immunological activity of an oligosaccharide-conjugate as a vaccine candidate against group A Streptococcus. Bioorg. Med. Chem. Lett. 2013, 23, 6038–6042. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, S.; Wang, G.; Li, H.; Guo, Z.; Gu, G. Synthesis and immunological studies of group A streptococcus cell-wall oligosaccharide-streptococcal C5a peptidase conjugates as bivalent vaccines. Org. Chem. Front. 2019, 6, 3589–3596. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, Y.; Wang, G.; Feng, S.; Guo, Z.; Gu, G. Group A streptococcus cell wall oligosaccharide-streptococcal C5a peptidase conjugates as effective antibacterial vaccines. ACS Infect. Dis. 2020, 6, 281–290. [Google Scholar] [CrossRef] [PubMed]
- van Sorge, N.M.; Cole, J.N.; Kuipers, K.; Henningham, A.; Aziz, R.K.; Kasirer-Friede, A.; Lin, L.; Berends, E.T.M.; Davies, M.R.; Dougan, G.; et al. The classical Lancefield antigen of group a streptococcus is a virulence determinant with implications for vaccine design. Cell. Host Microbe 2014, 15, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitirollo, O.; Micoli, F.; Necchi, F.; Mancini, F.; Carducci, M.; Adamo, R.; Evangelisti, C.; Morelli, L.; Polito, L.; Lay, L. Gold nanoparticles morphology does not affect the multivalent presentation and antibody recognition of group A Streptococcus synthetic oligorhamnans. Bioorg. Chem. 2020, 99, 103815. [Google Scholar] [CrossRef]
- Cleary, P.P.; Matsuka, Y.V.; Huynh, T.; Lam, H.; Olmsted, S.B. Immunization with C5a peptidase from either group A or B streptococci enhances clearance of group A streptococci from intranasally infected mice. Vaccine 2004, 22, 4332–4341. [Google Scholar] [CrossRef]
- Shet, A.; Kaplan, E.L.; Johnson, D.R.; Cleary, P.P. Immune response to group a streptococcal C5a peptidase in children: Implications for vaccine development. J. Infect. Dis. 2003, 188, 809–817. [Google Scholar] [CrossRef]
- Rodriguez-Ortega, M.J.; Norais, N.; Bensi, G.; Liberatori, S.; Capo, S.; Mora, M.; Scarselli, M.; Doro, F.; Ferrari, G.; Garaguso, I.; et al. Characterization and identification of vaccine candidate proteins through analysis of the group A streptococcus surface proteome. Nat. Biotechnol. 2006, 24, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Bensi, G.; Mora, M.; Tuscano, G.; Biagini, M.; Chiarot, E.; Bombaci, M.; Capo, S.; Falugi, F.; Manetti, A.G.O.; Donato, P.; et al. Multi high-throughput approach for highly selective identification of vaccine candidates: The group A streptococcus case. Mol. Cell. Proteom. 2012, 11, m111.015693. [Google Scholar] [CrossRef] [Green Version]
- Sheel, M.; Moreland, N.J.; Fraser, J.D.; Carapetis, J. Development of group A streptococcal vaccines: An unmet global health need. Expert Rev. Vaccines 2016, 15, 227–238. [Google Scholar] [CrossRef]
- Henningham, A.; Gillen, C.M.; Walker, M.J. Group A streptococcal vaccine candidates: Potential for the development of a human vaccine. Curr. Top. Microbiol. 2013, 368, 207–242. [Google Scholar]
- Steer, A.C.; Batzloff, M.R.; Mulholland, K.; Carapetis, J.R. Group A streptococcal vaccines: Facts versus fantasy. Curr. Opin. Infect. Dis. 2009, 22, 544–552. [Google Scholar] [CrossRef]
- Dale, J.B.; Fischetti, V.A.; Carapetis, J.R.; Steer, A.C.; Sow, S.; Kumar, R.; Mayosi, B.M.; Rubin, F.A.; Mulholland, K.; Hombach, J.M.; et al. Group A streptococcal vaccines: Paving a path for accelerated development. Vaccine 2013, 31, B216–B222. [Google Scholar] [CrossRef] [PubMed]
- Georgousakis, M.M.; McMillan, D.J.; Batzloff, M.R.; Sriprakash, K.S. Moving forward: A mucosal vaccine against group a streptococcus. Expert Rev. Vaccines 2009, 8, 747–760. [Google Scholar] [CrossRef] [PubMed]
- Steer, A.C.; Dale, J.B.; Carapetis, J.R. Progress toward a global group a streptococcal vaccine. Pediatr. Infect. Dis. J. 2013, 32, 180–182. [Google Scholar] [CrossRef] [PubMed]
- Dale, J.B.; Walker, M.J. Update on group a streptococcal vaccine development. Curr. Opin. Infect. Dis. 2020, 33, 244–250. [Google Scholar] [CrossRef]
- Cleary, P.P.; Prahbu, U.; Dale, J.B.; Wexler, D.E.; Handley, J. Streptococcal-C5a peptidase is a highly specific endopeptidase. Infect. Immun. 1992, 60, 5219–5223. [Google Scholar] [CrossRef] [Green Version]
- Manthey, H.D.; Woodruff, T.M.; Taylor, S.M.; Monk, P.N. Complement component 5a (C5a). Int. J. Biochem. Cell B 2009, 41, 2114–2117. [Google Scholar] [CrossRef]
- Ji, Y.D.; McLandsborough, L.; Kondagunta, A.; Cleary, P.P. C5a peptidase alters clearance and trafficking of group A streptococci by infected mice. Infect. Immun. 1996, 64, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.; Cleary, P.P. Complete nucleotide-sequence of the streptococcal C5a peptidase gene of Streptococcus pyogenes. J. Biol. Chem. 1990, 265, 3161–3167. [Google Scholar] [CrossRef]
- Ji, Y.D.; Carlson, B.; Kondagunta, A.; Cleary, P.P. Intranasal immunization with C5a peptidase prevents nasopharyngeal colonization of mice by the group A Streptococcus. Infect. Immun. 1997, 65, 2080–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oconnor, S.P.; Darip, D.; Fraley, K.; Nelson, C.M.; Kaplan, E.L.; Cleary, P.P. The human-antibody response to streptococcal C5a peptidase. J. Infect. Dis. 1991, 163, 109–116. [Google Scholar] [CrossRef]
- Xu, Z.; Rivera-Hernandez, T.; Moyle, P.M. Development of an enzyme-mediated, site-specific method to conjugate Toll-like receptor 2 agonists onto protein antigens: Toward a broadly protective, four components, group A streptococcal self-adjuvanting lipoprotein-fusion combination vaccine. ACS Infect. Dis. 2020, 6, 1770–1782. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Hernandez, T.; Carnathan, D.G.; Jones, S.; Cork, A.J.; Davies, M.R.; Moyle, P.M.; Toth, I.; Batzloff, M.R.; McCarthy, J.; Nizet, V.; et al. An experimental group a streptococcus vaccine that reduces pharyngitis and tonsillitis in a nonhuman primate model. mBio 2019, 10, e00693-19. [Google Scholar] [CrossRef] [Green Version]
- Bi, S.; Xu, M.Y.; Zhou, Y.; Xing, X.X.; Shen, A.; Wang, B.N. A multicomponent vaccine provides immunity against local and systemic infections by group a streptococcus across serotypes. mBio 2019, 10, e02600-19. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Hernandez, T.; Rhyme, M.S.; Cork, A.J.; Jones, S.; Segui-Perez, C.; Brunner, L.; Richter, J.; Petrovsky, N.; Lawrenz, M.; Goldblatt, D.; et al. Vaccine-induced Th1-type response protects against invasive group A streptococcus infection in the absence of opsonizing antibodies. mBio 2020, 11, e00122-20. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, S.; Zhao, Y.; Chen, Z.; Gu, G.; Guo, Z. Mutagenesis and immunological evaluation of group A streptococcal C5a peptidase as an antigen for vaccine development and as a carrier protein for glycoconjugate vaccine design. RSC Adv. 2017, 7, 42056–42063. [Google Scholar] [CrossRef] [Green Version]
- Kagawa, T.F.; O’Connell, M.R.; Mouat, P.; Paoli, M.; O’Toole, P.W.; Cooney, J.C. Model for substrate interactions in C5a peptidase from Streptococcus pyogenes: A 1.9 Å crystal structure of the active form of ScpA. J. Biol. Chem. 2009, 386, 754–772. [Google Scholar] [CrossRef]
- Stafslien, D.K.; Cleary, P.P. Characterization of the streptococcal C5a peptidase using a C5a-green fluorescent protein fusion protein substrate. J. Bacteriol. 2000, 182, 3254–3258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.T.; Wetherell, M.G.; Winter, L.A.; Olmsted, S.B.; Cleary, P.P.; Matsuka, Y.V. Processing, stability, and kinetic parameters of C5a peptidase from Streptococcus pyogenes. Eur. J. Biochem. 2002, 269, 4839–4851. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, T.; Anam, K.; Ali, N. A mixed Th1/Th2 response elicited by a liposomal formulation of Leishmania vaccine instructs Thl responses and resistance to Leishmania donovani in susceptible BALB/c mice. Vaccine 2004, 22, 1162–1171. [Google Scholar] [CrossRef]
- Mygind, T.; Vandahl, B.; Pedersen, A.S.; Christiansen, G.; Hollsberg, P.; Birkelund, S. Identification of an in vivo CD4(+) T cell-mediated response to polymorphic membrane proteins of Chlamydia pneumoniae during experimental infection. Fems Immunol. Med. Mic. 2004, 40, 129–137. [Google Scholar] [CrossRef]
- Chmouryguina, I.; Suvorov, A.; Ferrieri, P.; Cleary, P.P. Conservation of the C5a peptidase genes in group A and B streptococci. Infect. Immun. 1996, 64, 2387–2390. [Google Scholar] [CrossRef] [Green Version]
- Cleary, P.P.; Handley, J.; Suvorov, A.N.; Podbielski, A.; Ferrieri, P. Similarity between the group B and A streptococcal C5a peptidase genes. Infect. Immun. 1992, 60, 4239–4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.K.; Gu, Z.Y.; Matsuka, Y.V.; Purushothaman, S.S.; Winter, L.A.; Cleary, P.P.; Olmsted, S.B.; Ohlendorf, D.H.; Earhart, C.A. Structure of the streptococcal cell wall C5a peptidase. Proc. Natl. Acad. Sci. USA 2005, 102, 18391–18396. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Xiong, C.; Wang, G.; Wang, S.; Jin, G.; Gu, G. Exploration of recombinant fusion proteins YAPO and YAPL as carrier proteins for glycoconjugate vaccine design against Streptococcus pneumoniae infection. ACS Infect. Dis. 2020, 6, 2181–2191. [Google Scholar] [CrossRef]
- Bonam, S.R.; Bhunia, D.; Muller, S.; Nerella, S.G.; Alvala, M.; Halmuthur Mahabalarao, S.K. Novel trisaccharide based phospholipids as immunomodulators. Int. Immunopharmacol. 2019, 74, 105684. [Google Scholar] [CrossRef]
- Ravetch, J.V.; Kinet, J.P. Fc-receptors. Annu. Rev. Immunol. 1991, 9, 457–492. [Google Scholar] [CrossRef]
- Michaelsen, T.E.; Kolberg, J.; Aase, A.; Herstad, T.K.; Hoiby, E.A. The four mouse IgG isotypes differ extensively in bactericidal and opsonophagocytic activity when reacting with the P1.16 epitope on the outer membrane PorA protein of Neisseria meningitidis. Scand. J. Immunol. 2004, 59, 34–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefeber, D.J.; Benaissa-Trouw, B.; Vliegenthart, J.F.G.; Kamerling, J.P.; Jansen, W.T.M.; Kraaijeveld, K.; Snippe, H. Th1-directing adjuvants increase the immunogenicity of oligosaccharide-protein conjugate vaccines related to Streptococcus pneumoniae type 3. Infect. Immun. 2003, 71, 6915–6920. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glycoconjugate | Fn-Tri | Fn2-Tri | rsScpA193-Tri | CRM197-Tri | BSA-Tri |
|---|---|---|---|---|---|
| Oligosaccharide chains /carrier protein (m) | 4.3 | 2.8 | 16.3 | 5.5 | 8.3 |
| Carbohydrate loading (%) | 5.4 | 7.2 | 9.7 | 5.8 | 7.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Zhao, J.; Zhao, Y.; Wang, S.; Feng, S.; Gu, G. Immunogenicity Assessment of Different Segments and Domains of Group A Streptococcal C5a Peptidase and Their Application Potential as Carrier Protein for Glycoconjugate Vaccine Development. Vaccines 2021, 9, 139. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020139
Wang G, Zhao J, Zhao Y, Wang S, Feng S, Gu G. Immunogenicity Assessment of Different Segments and Domains of Group A Streptococcal C5a Peptidase and Their Application Potential as Carrier Protein for Glycoconjugate Vaccine Development. Vaccines. 2021; 9(2):139. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020139
Chicago/Turabian StyleWang, Guirong, Jielin Zhao, Yisheng Zhao, Subo Wang, Shaojie Feng, and Guofeng Gu. 2021. "Immunogenicity Assessment of Different Segments and Domains of Group A Streptococcal C5a Peptidase and Their Application Potential as Carrier Protein for Glycoconjugate Vaccine Development" Vaccines 9, no. 2: 139. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020139