
Challenge of Naïve and Vaccinated Pigs with a Vaccine-Derived Recombinant Porcine Reproductive and Respiratory Syndrome Virus 1 Strain (Horsens Strain)

 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Design
2.2. Challenge Viruses and Inoculum
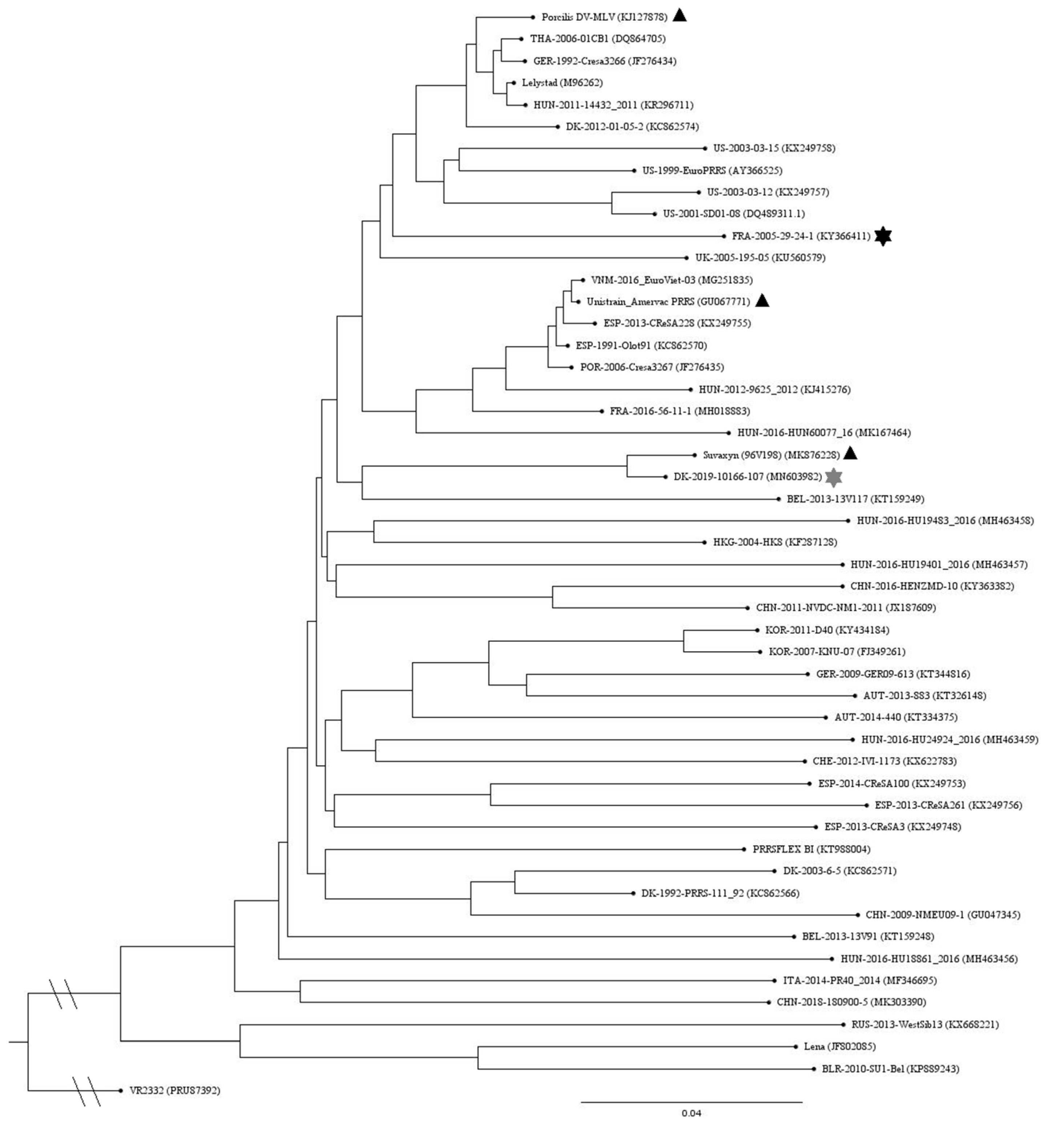
2.3. Genetic Comparisons of the Vaccine and Challenge Strains
2.4. Sampling
2.5. Clinical Observation
2.6. Necropsy
2.7. RNA Extraction and RT-qPCR Assays
SYBR Green Real-Time RT-PCR Assay Specific for the Horsens Strain
2.8. Antibody ELISA
2.9. Laboratory Analyses
2.10. Statistical Analysis
3. Results
3.1. Genetic and Antigenic Analysis of Vaccine and Challenge Strains
3.2. Clinical Signs, Rectal Temperature and Body Weight Gains
3.3. Macroscopic Changes
3.4. PRRS Virus in Serum and OF
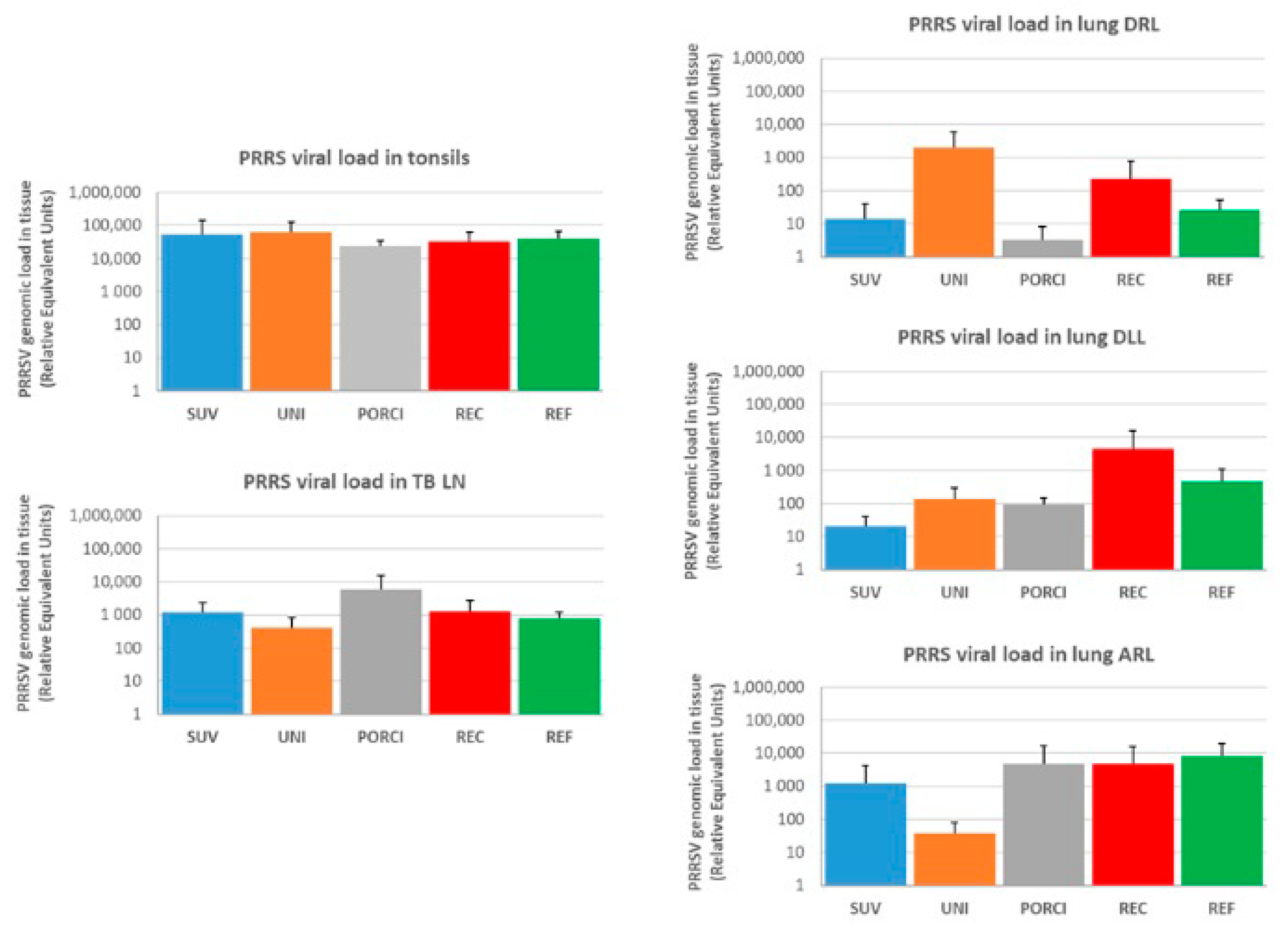
3.5. PRRS Viral Load in Tissue
3.6. PRRSV Antibodies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Afonso, C.L.; Amarasinghe, G.K.; Banyai, K.; Bao, Y.; Basler, C.F.; Bavari, S.; Bejerman, N.; Blasdell, K.R.; Briand, F.-X.; Briese, T.; et al. Taxonomy of the order Mononegavirales: Update 2016. Arch. Virol. 2016, 161, 2351–2360. [Google Scholar] [CrossRef]
- Kvisgaard, L.K.; Kristensen, C.S.; Ryt-Hansen, P.; Pedersen, K.; Stadejek, T.; Trebbien, R.; Andresen, L.O.; Larsen, L.E. A recombination between two Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-1) vaccine strains has caused severe outbreaks in Danish pigs. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristensen, C.S.; Christiansen, M.G.; Pedersen, K.; Larsen, L.E. Production losses five months after outbreak with a recombinant of two PRRSV vaccine strains in 13 Danish sow herds. Porc. Health Manag. 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Genís, S.; Kvisgaard, L.K.; Larsen, L.E.; Taylor, L.P.; Calvert, J.G.; Balasch, M. Assessment of the impact of the recombinant porcine reproductive and respiratory syndrome virus horsens strain on the reproductive performance in pregnant sows. Pathogens 2020, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Weesendorp, E.; Morgan, S.; Stockhofe-Zurwieden, N.; Popma-De Graaf, D.J.; Graham, S.P.; Rebel, J.M. Comparative analysis of immune responses following experimental infection of pigs with European porcine reproductive and respiratory syndrome virus strains of differing virulence. Vet. Microbiol. 2013, 163, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kvisgaard, L.K.; Hjulsager, C.K.; Kristensen, C.S.; Lauritsen, K.T.; Larsen, L.E. Genetic and antigenic characterization of complete genomes of Type 1 Porcine Reproductive and Respiratory Syndrome viruses (PRRSV) isolated in Denmark over a period of 10 years. Virus Res. 2013, 178, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wernike, K.; Bonilauri, P.; Dauber, M.; Errington, J.; LeBlanc, N.; Revilla-Fernández, S.; Hjulsager, C.; Isaksson, M.; Stadejek, T.; Beer, M.; et al. Porcine reproductive and respiratory syndrome virus: Interlaboratory ring trial to evaluate real-time reverse transcription polymerase chain reaction detection methods. J. Vet. Diagn. Investig. 2012, 24, 855–866. [Google Scholar] [CrossRef] [Green Version]
- Rose, N.; Renson, P.; Andraud, M.; Paboeuf, F.; Le Potier, M.F.; Bourry, O. Porcine reproductive and respiratory syndrome virus (PRRSv) modified-live vaccine reduces virus transmission in experimental conditions. Vaccine 2015, 33, 2493–2499. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.P.; Genzow, M. Immunological solutions for treatment and prevention of porcine reproductive and respiratory syndrome (PRRS). Vaccine 2011, 29, 8192–8204. [Google Scholar] [CrossRef]
- Stadejek, T.; Larsen, L.E.; Podgórska, K.; Bøtner, A.; Botti, S.; Dolka, I.; Fabisiak, M.; Heegaard, P.M.H.; Hjulsager, C.K.; Huć, T.; et al. Pathogenicity of three genetically diverse strains of PRRSV Type 1 in specific pathogen free pigs. Vet. Microbiol. 2017, 209, 1–7. [Google Scholar] [CrossRef]
- Frydas, I.S.; Trus, I.; Kvisgaard, L.K.; Bonckaert, C.; Reddy, V.R.A.P.; Li, Y.; Larsen, L.E.; Nauwynck, H.J. Different clinical, virological, serological and tissue tropism outcomes of two new and one old Belgian type 1 subtype 1 porcine reproductive and respiratory virus (PRRSV) isolates. Vet. Res. 2015, 46, 37. [Google Scholar] [CrossRef] [Green Version]
- Sinn, L.J.; Klingler, E.; Lamp, B.; Brunthaler, R.; Weissenböck, H.; Rümenapf, T.; Ladinig, A. Emergence of a virulent porcine reproductive and respiratory syndrome virus (PRRSV) 1 strain in Lower Austria. Porc. Health Manag. 2016, 2, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvisgaard, L.K.; Larsen, L.E.; Hjulsager, C.K.; Botner, A.; Rathkjen, P.H.; Heegaard, P.M.H.; Bisgaard, N.P.; Nielsen, J.; Hansen, M.S. Genetic and biological characterization of a Porcine Reproductive and Respiratory Syndrome Virus 2 (PRRSV-2) causing significant clinical disease in the field. Vet. Microbiol. 2017, 211, 74–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canelli, E.; Catella, A.; Borghetti, P.; Ferrari, L.; Ogno, G.; De Angelis, E.; Bonilauri, P.; Guazzetti, S.; Nardini, R.; Martelli, P. Efficacy of a modified-live virus vaccine in pigs experimentally infected with a highly pathogenic porcine reproductive and respiratory syndrome virus type 1 (HP-PRRSV-1). Vet. Microbiol. 2018, 226, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Eclercy, J.; Renson, P.; Lebret, A.; Hirchaud, E.; Normand, V.; Andraud, M.; Paboeuf, F.; Blanchard, Y.; Rose, N.; Bourry, O. A field recombinant strain derived from two type 1 porcine reproductive and respiratory syndrome virus (PRRSV-1) modified live vaccines shows increased viremia and transmission in spf pigs. Viruses 2019, 11, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karniychuk, U.U.; Geldhof, M.; Vanhee, M.; Van Doorsselaere, J.; Saveleva, T.A.; Nauwynck, H.J. Pathogenesis and antigenic characterization of a new East European subtype 3 porcine reproductive and respiratory syndrome virus isolate. BMC Vet. Res. 2010, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Sinn, L.J.; Zieglowski, L.; Koinig, H.; Lamp, B.; Jansko, B.; Mößlacher, G.; Riedel, C.; Hennig-Pauka, I.; Rümenapf, T. Characterization of two Austrian porcine reproductive and respiratory syndrome virus (PRRSV) field isolates reveals relationship to East Asian strains. Vet. Res. 2016, 47. [Google Scholar] [CrossRef] [Green Version]
- Renson, P.; Rose, N.; Le Dimna, M.; Mahé, S.; Keranflec’h, A.; Paboeuf, F.; Belloc, C.; Le Potier, M.F.; Bourry, O. Dynamic changes in bronchoalveolar macrophages and cytokines during infection of pigs with a highly or low pathogenic genotype 1 PRRSV strain. Vet. Res. 2017, 48, 15. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Han, Q.; Zhang, L.; Zhang, Z.; Wu, Y.; Shen, H.; Jiang, P. Emergence of mosaic recombinant strains potentially associated with vaccine JXA1-R and predominant circulating strains of porcine reproductive and respiratory syndrome virus in different provinces of China. Virol. J. 2017, 14, 1–10. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, X.; Zhai, J.; Wei, C.; Dai, A.; Yang, X.; Luo, M. Recombination in JXA1-R vaccine and NADC30-like strain of porcine reproductive and respiratory syndrome viruses. Vet. Microbiol. 2017, 204, 110–120. [Google Scholar] [CrossRef]
- Dortmans, J.C.F.M.; Buter, G.J.; Dijkman, R.; Houben, M.; Duinhof, T.F. Molecular characterization of type 1 porcine reproductive and respiratory syndrome viruses (PRRSV) isolated in the Netherlands from 2014 to 2016. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Martín-Valls, G.E.; Kvisgaard, L.K.; Tello, M.; Darwich, L.; Cortey, M.; Burgara-Estrella, A.J.; Hernández, J.; Larsen, L.E.; Mateu, E. Analysis of ORF5 and full-length genome sequences of porcine reproductive and respiratory syndrome virus isolates of genotypes 1 and 2 retrieved worldwide provides evidences supporting that recombination is a common phenomenon and may produce mosaic isola. J. Virol. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marton, S.; Szalay, D.; Kecskeméti, S.; Forró, B.; Olasz, F.; Zádori, Z.; Szabó, I.; Molnár, T.; Bányai, K.; Bálint, Á. Coding-complete sequence of a vaccine-derived recombinant porcine reproductive and respiratory syndrome virus strain isolated in Hungary. Arch. Virol. 2019, 164, 2605–2608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frossard, J.-P.; Hughes, G.J.; Westcott, D.G.; Naidu, B.; Williamson, S.; Woodger, N.G.A.; Steinbach, F.; Drew, T.W. Porcine reproductive and respiratory syndrome virus: Genetic diversity of recent British isolates. Vet. Microbiol. 2013, 162, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Renson, P.; Touzain, F.; Lebret, A.; Le Dimna, M.; Quenault, H.; Normand, V.; Claude, J.-B.; Pez, F.; Rose, N.; Blanchard, Y.; et al. Complete Genome Sequence of a Recombinant Porcine Reproductive and Respiratory Syndrome Virus Strain from Two Genotype 1 Modified Live Virus Vaccine Strains. Genome Announc. 2017, 5, e00454-17. [Google Scholar] [CrossRef] [Green Version]
- Van Geelen, A.G.M.; Anderson, T.K.; Lager, K.M.; Das, P.B.; Otis, N.J.; Montiel, N.A.; Miller, L.C.; Kulshreshtha, V.; Buckley, A.C.; Brockmeier, S.L.; et al. Porcine reproductive and respiratory disease virus: Evolution and recombination yields distinct ORF5 RFLP 1-7-4 viruses with individual pathogenicity. Virology 2018, 513, 168–179. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | No. of Pigs | Vaccination (dpv 0) | Challenge (dpv 28) |
|---|---|---|---|
| 1 (SUV) | 6 | Suvaxyn PRRS MLV | Horsens |
| 2 (UNI) | 6 | Unistrain®PRRS | Horsens |
| 3 (PORCI) | 6 | Porcilis®PRRS | Horsens |
| 4 (REC) | 6 | None | Horsens |
| 5 (REF) | 6 | None | Finistere |
| 6 (CTRL) | 6 | None | None |
| Days Post-Challenge | ||||||
|---|---|---|---|---|---|---|
| Group | −4 | 3 | 8 | 14 | 21 | 28 |
| SUV | ||||||
| UNI | ||||||
| PORCI | ||||||
| REC | ||||||
| REF | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kvisgaard, L.K.; Larsen, L.E.; Kristensen, C.S.; Paboeuf, F.; Renson, P.; Bourry, O. Challenge of Naïve and Vaccinated Pigs with a Vaccine-Derived Recombinant Porcine Reproductive and Respiratory Syndrome Virus 1 Strain (Horsens Strain). Vaccines 2021, 9, 417. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9050417
Kvisgaard LK, Larsen LE, Kristensen CS, Paboeuf F, Renson P, Bourry O. Challenge of Naïve and Vaccinated Pigs with a Vaccine-Derived Recombinant Porcine Reproductive and Respiratory Syndrome Virus 1 Strain (Horsens Strain). Vaccines. 2021; 9(5):417. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9050417
Chicago/Turabian StyleKvisgaard, Lise K., Lars E. Larsen, Charlotte S. Kristensen, Frédéric Paboeuf, Patricia Renson, and Olivier Bourry. 2021. "Challenge of Naïve and Vaccinated Pigs with a Vaccine-Derived Recombinant Porcine Reproductive and Respiratory Syndrome Virus 1 Strain (Horsens Strain)" Vaccines 9, no. 5: 417. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9050417