
Production and Immunogenicity of a Tag-Free Recombinant Chimera Based on PfMSP-1 and PfMSP-3 Using Alhydrogel and Dipeptide-Based Hydrogels

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tag-Free rFu24 Cloning in pET-24b Vector
2.2. Expression and Purification of the Full-Length rFu24
2.3. Characterization of Recombinant rFu24
2.4. Synthesis of Dipeptides
2.5. Preparation of Hydrogel and Entrapment of Antigen
2.6. Characterization of F∆F and L∆F Hydrogels
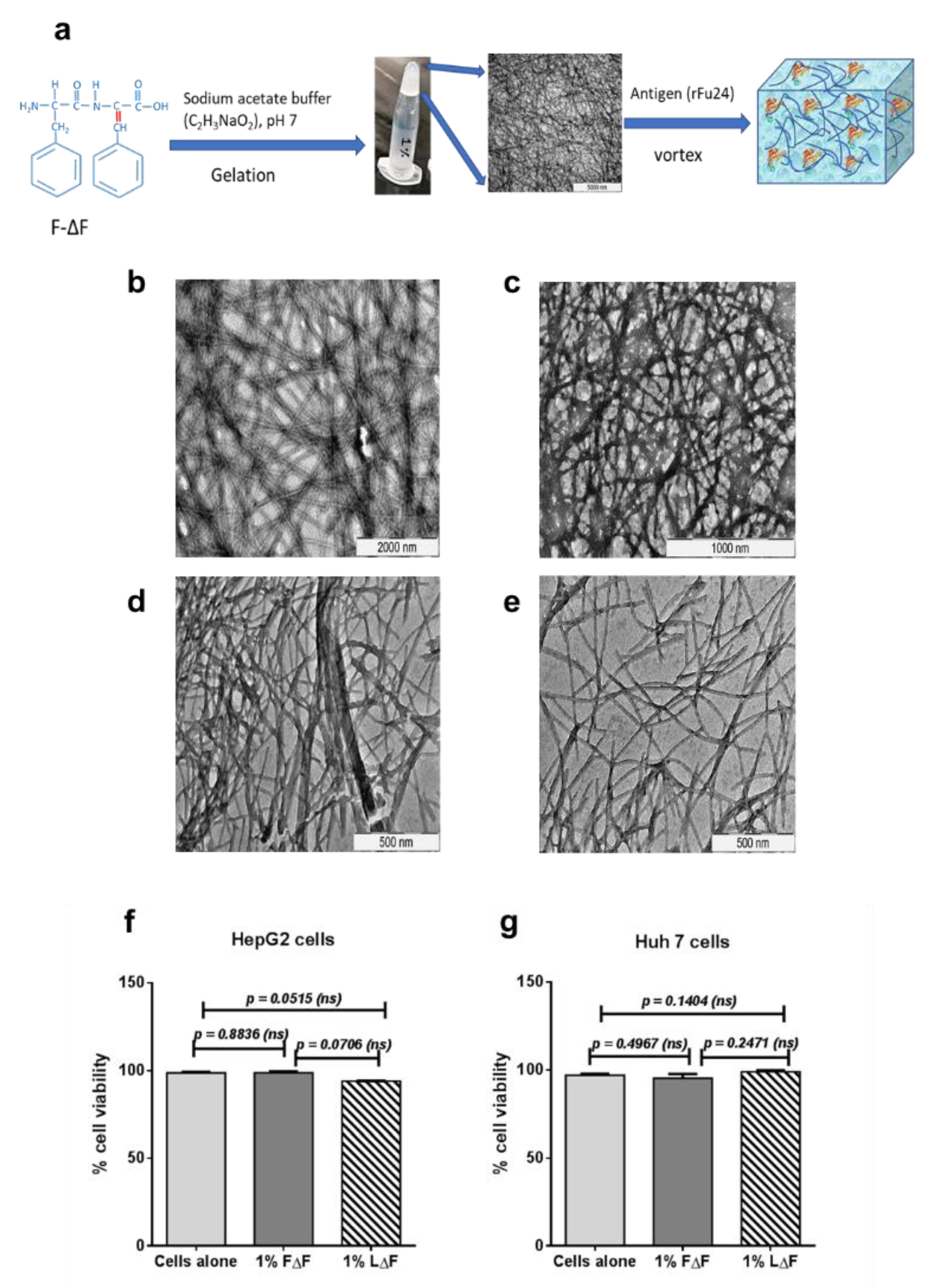
2.7. Cell Viability Experiments
2.8. Immunization of Mice with rFu24
2.9. Ethics Statement
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Invasion Inhibition Assay
2.12. Statistics
3. Results
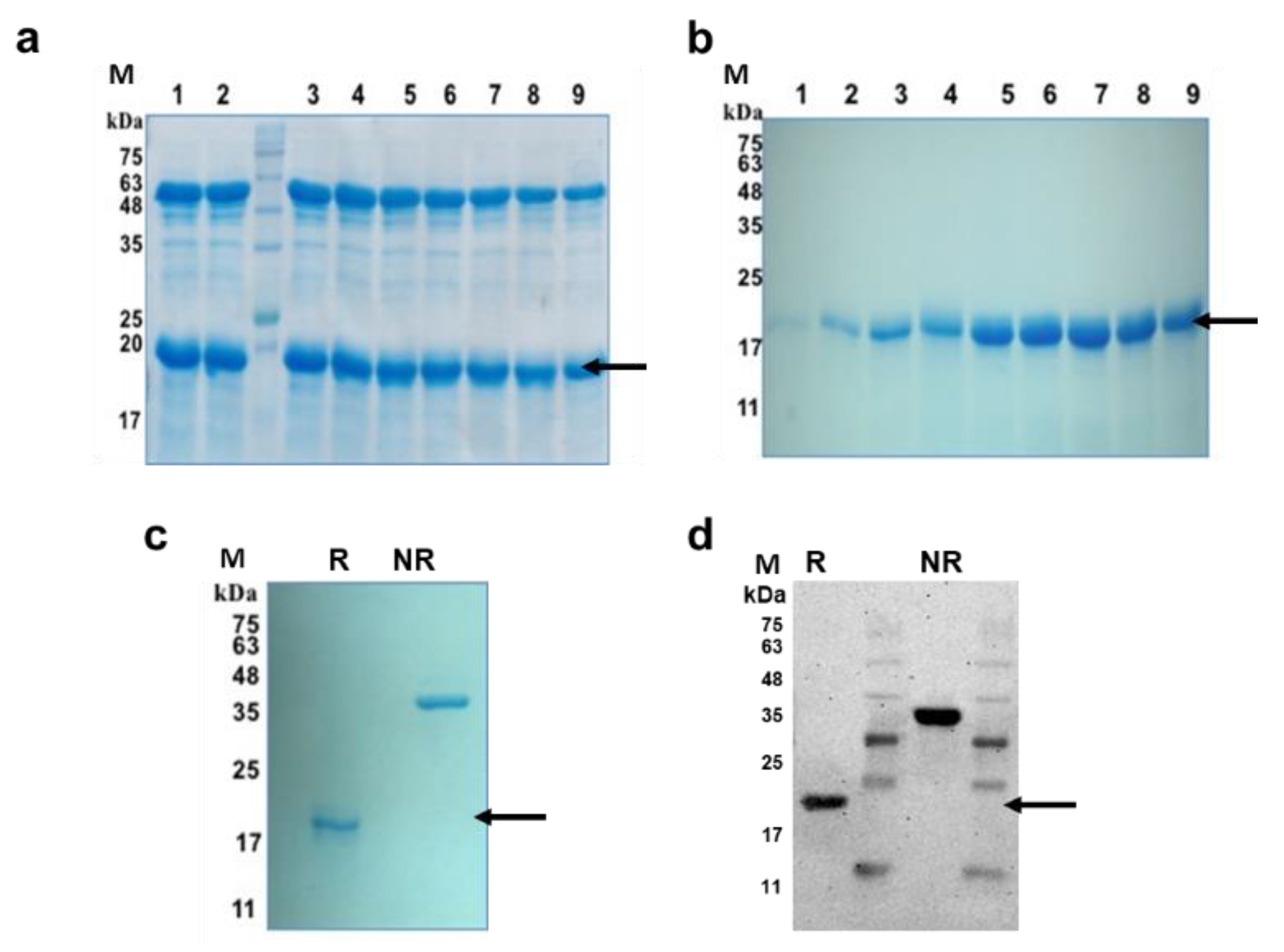
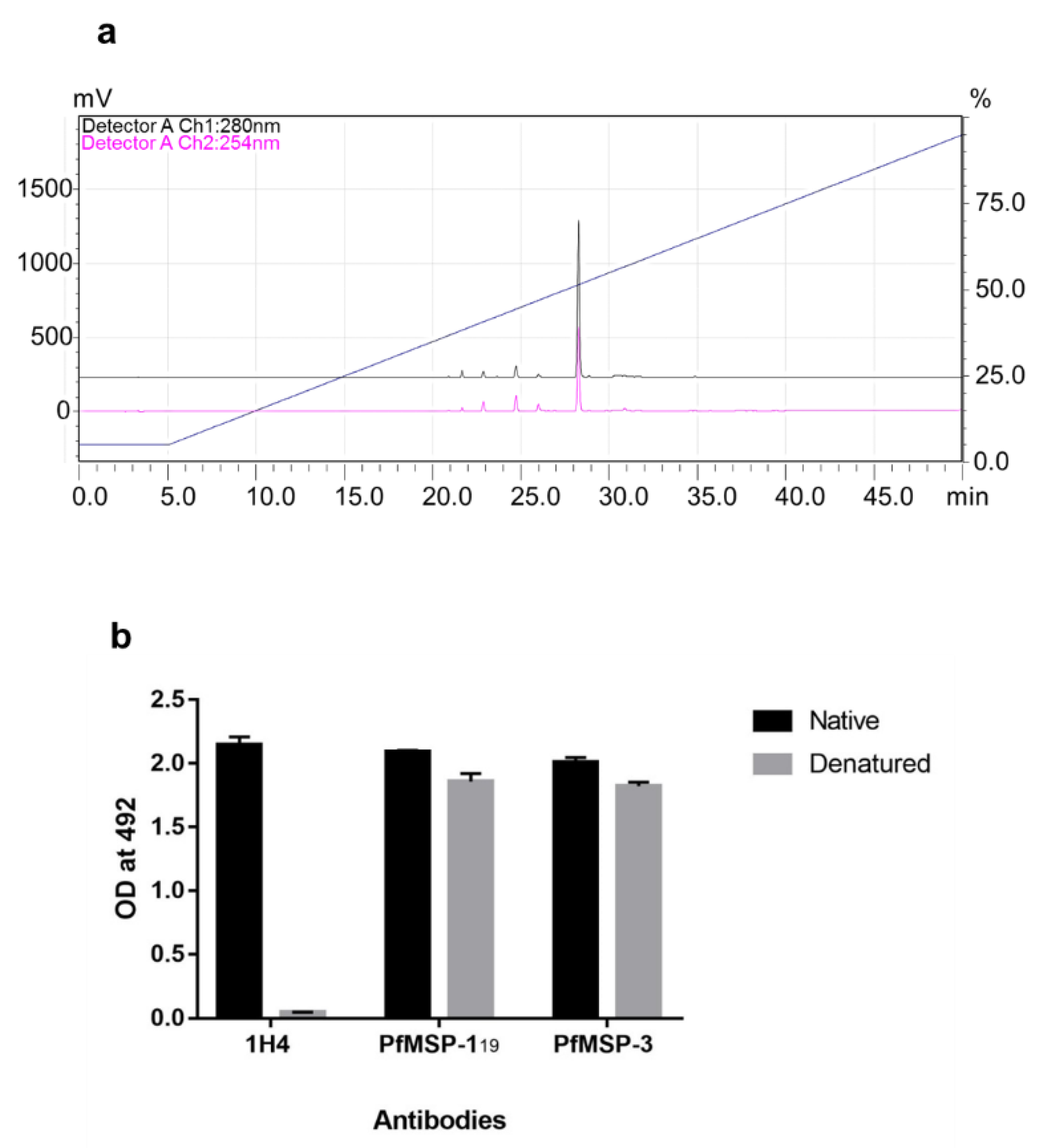
3.1. Expression, Purification and Characterization of 6-His Tag Free PfMSPFu24 (rFu24)
3.2. Synthesis and Characterization Dipeptide F∆F and L∆F Hydrogel
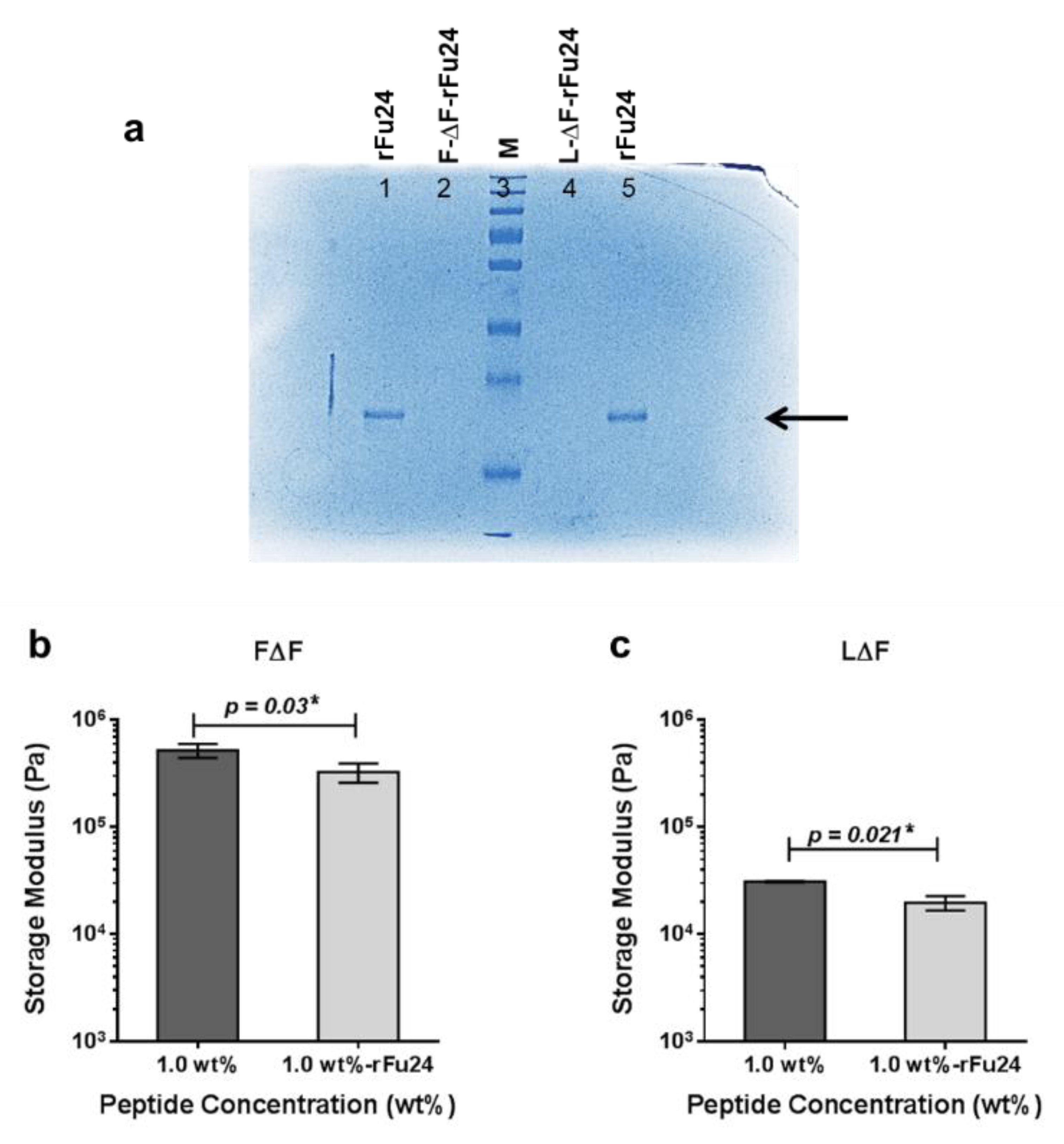
3.3. Entrapment of rFu24 and Its Rheological Characterization
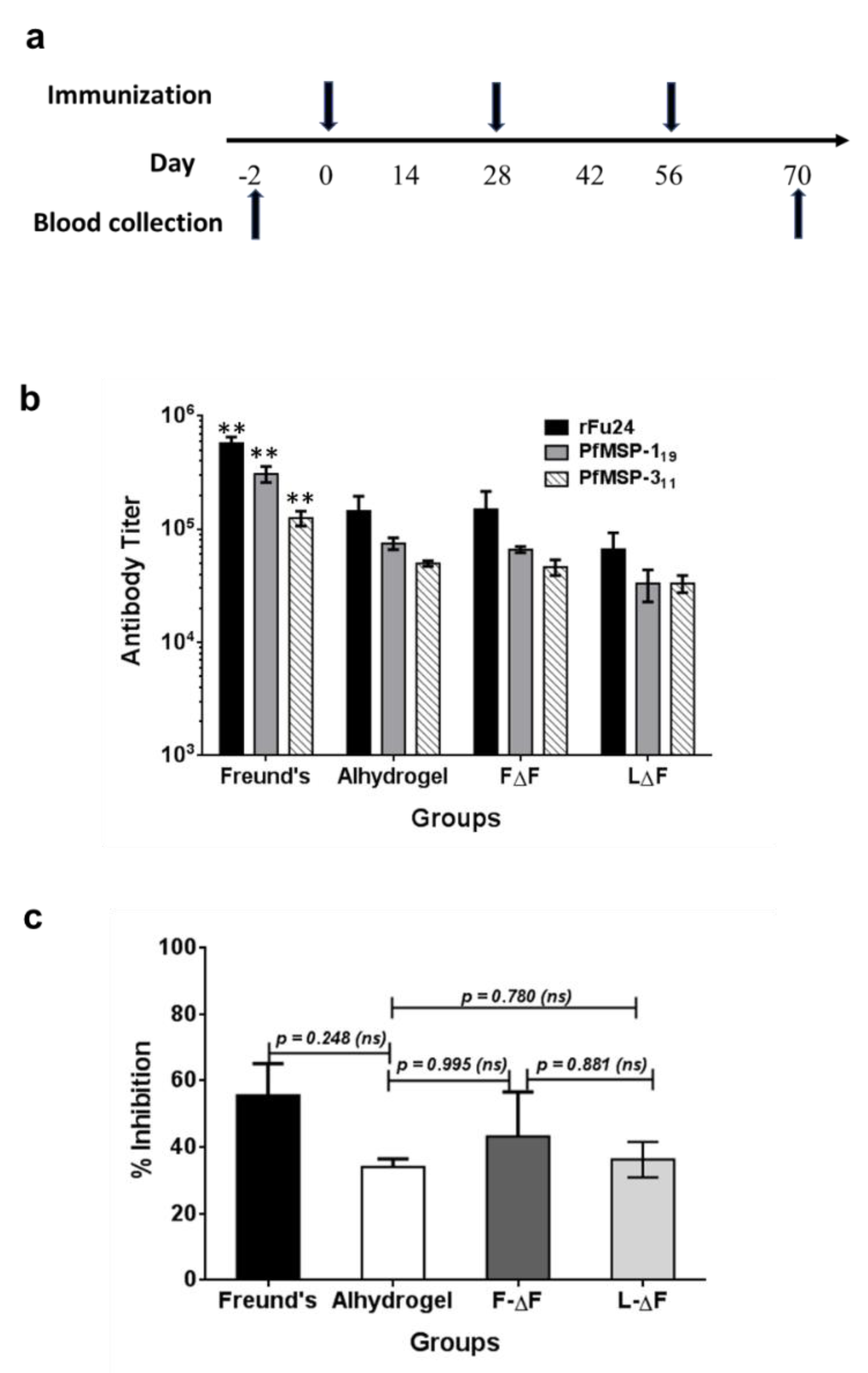
3.4. Immunogenicity of rFu24
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Alonso, P.L.; Brown, G.; Arevalo-Herrera, M.; Binka, F.; Chitnis, C.; Collins, F.; Doumbo, O.K.; Greenwood, B.; Hall, B.F.; Levine, M.M.; et al. A research agenda to underpin malaria eradication. PLoS Med. 2011, 8, e1000406. [Google Scholar] [CrossRef] [PubMed]
- Healer, J.; Cowman, A.F.; Kaslow, D.C.; Birkett, A.J. Vaccines to accelerate malaria elimination and eventual eradication. Cold Spring Harb. Perspect. Med. 2017, 7, a025627. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, M.; Ogutu, B.; Sewankambo, N.K.; Biller-Andorno, N.; Tanner, M. RTS,S malaria vaccine pilot studies: Addressing the human realities in large-scale clinical trials. Trials 2019, 20, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanner, M.; Greenwood, B.; Whitty, C.J.M.; Ansah, E.K.; Price, R.N.; Dondorp, A.M.; Von Seidlein, L.; Baird, J.K.; Beeson, J.G.; Fowkes, F.J.; et al. Malaria eradication and elimination: Views on how to translate a vision into reality. BMC Med. 2015, 13, 167. [Google Scholar] [CrossRef] [PubMed]
- Goodman, A.L.; Draper, S.J. Blood-stage malaria vaccines—Recent progress and future challenges. Ann. Trop. Med. Parasitol. 2010, 104, 189–211. [Google Scholar] [CrossRef] [PubMed]
- Takala, S.L.; Coulibaly, D.; Thera, A.H.; Batchelor, A.H.; Cummings, M.P.; Escalante, A.A.; Ouattara, A.; Traoré, K.; Niangaly, A.; Djimdé, A.A.; et al. Extreme polymorphism in a vaccine antigen and risk of clinical malaria: Implications for vaccine development. Sci. Transl. Med. 2009, 1, 2ra5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, G.J.; Rayner, J.C. Plasmodium falciparum erythrocyte invasion: Combining function with immune evasion. PLoS Pathog. 2014, 10, e1003943. [Google Scholar] [CrossRef] [Green Version]
- Miura, K. Progress and prospects for blood-stage malaria vaccines. Expert Rev. Vaccines 2016, 15, 765–781. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, L.; Brown, G.V.; Genton, B.; Moorthy, V.S. A review of malaria vaccine clinical projects based on the WHO rainbow table. Malar. J. 2012, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Ogutu, B.R.; Apollo, O.J.; McKinney, D.; Okoth, W.; Siangla, J.; Dubovsky, F.; Tucker, K.; Waitumbi, J.N.; Diggs, C.; Wittes, J.; et al. Blood stage malaria vaccine eliciting high antigen-specific antibody concentrations confers no protection to young children in Western Kenya. PLoS ONE 2009, 4, e4708. [Google Scholar] [CrossRef] [PubMed]
- Dent, A.E.; Chelimo, K.; O Sumba, P.; Spring, M.D.; Crabb, B.S.; Moormann, A.M.; Tisch, D.J.; Kazura, J.W. Temporal stability of naturally acquired immunity to Merozoite Surface Protein-1 in Kenyan adults. Malar. J. 2009, 8, 162. [Google Scholar] [CrossRef] [Green Version]
- Crompton, P.D.; Kayala, M.A.; Traore, B.; Kayentao, K.; Ongoiba, A.; Weiss, G.E.; Molina, D.M.; Curk, C.R.; Waisberg, M.; Jasinskas, A.; et al. A prospective analysis of the Ab response to Plasmodium falciparum before and after a malaria season by protein microarray. Proc. Natl. Acad. Sci. USA 2010, 107, 6958–6963. [Google Scholar] [CrossRef] [Green Version]
- Thera, M.A.; Doumbo, O.K.; Coulibaly, D.; Laurens, M.B.; Ouattara, A.; Kone, A.K.; Guindo, A.B.; Traore, K.; Traore, I.; Kouriba, B.; et al. A field trial to assess a blood-stage malaria vaccine. N. Engl. J. Med. 2011, 365, 1004–1013. [Google Scholar] [CrossRef] [Green Version]
- Chitnis, N.; Hyman, J.M.; Cushing, J.M. Determining important parameters in the spread of malaria through the sensitivity analysis of a mathematical model. Bull. Math. Biol. 2008, 70, 1272–1296. [Google Scholar] [CrossRef]
- Dondorp, A.M.; Yeung, S.; White, L.; Nguon, C.; Day, N.P.; Socheat, D.; Von Seidlein, L. Artemisinin resistance: Current status and scenarios for containment. Nat. Rev. Microbiol. 2010, 8, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Draper, S.J.; Angov, E.; Horii, T.; Miller, L.H.; Srinivasan, P.; Theisen, M.; Biswas, S. Recent advances in recombinant protein-based malaria vaccines. Vaccine 2015, 33, 7433–7443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, R.A.; Hackett, F.; Howell, S.A.; Treeck, M.; Struck, N.; Krnajski, Z.; Withers-Martinez, C.; Gilberger, T.-W.; Blackman, M.J. Intramembrane proteolysis mediates shedding of a key adhesin during erythrocyte invasion by the malaria parasite. J. Cell Biol. 2006, 174, 1023–1033. [Google Scholar] [CrossRef]
- Baldwin, M.R.; Li, X.; Hanada, T.; Liu, S.C.; Chishti, A.H. Merozoite surface protein 1 recognition of host glycophorin A mediates malaria parasite invasion of red blood cells. Blood 2015, 125, 2704–2711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, M.J.; Wilson, D.W.; Richards, J.S.; Riglar, D.T.; Tetteh, K.K.A.; Conway, D.; Ralph, S.A.; Baum, J.; Beeson, J.G. Isolation of viable Plasmodium falciparum merozoites to define erythrocyte invasion events and advance vaccine and drug development. Proc. Natl. Acad. Sci. USA 2010, 107, 14378–14383. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Hertrich, N.; Perrin, A.J.; Withers-Martinez, C.; Collins, C.R.; Jones, M.L.; Watermeyer, J.M.; Fobes, E.T.; Martin, S.R.; Saibil, H.R.; et al. Processing of Plasmodium falciparum Merozoite Surface Protein MSP1 activates a Spectrin-binding function enabling parasite egress from RBCs. Cell Host Microbe 2015, 18, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Blackman, M.J.; Heidrich, H.G.; Donachie, S.; McBride, J.S.; Holder, A.A. A single fragment of a malaria merozoite surface protein remains on the parasite during red cell invasion and is the target of invasion-inhibiting antibodies. J. Exp. Med. 1990, 172, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Eksi, S.; Czesny, B.; van Gemert, G.-J.; Sauerwein, R.W.; Eling, W.; Williamson, K.C. Malaria transmission-blocking antigen, Pfs230, mediates human red blood cell binding to exflagellating male parasites and oocyst production. Mol. Microbiol. 2006, 61, 991–998. [Google Scholar] [CrossRef]
- Egan, A.F.; Morris, J.; Barnish, G.; Allen, S.; Greenwood, B.M.; Kaslow, D.C.; Holder, A.A.; Riley, E.M. Clinical immunity to Plasmodium falciparum malaria is associated with serum antibodies to the 19-kDa C-terminal fragment of the merozoite surface antigen, PfMSP-1. J. Infect. Dis. 1996, 173, 765–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holder, A.A. The carboxy-terminus of merozoite surface protein 1: Structure, specific antibodies and immunity to malaria. Parasitology 2009, 136, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Tiendrebeogo, R.W.; Adu, B.; Singh, S.K.; Dziegiel, M.H.; Nébié, I.; Sirima, S.B.; Christiansen, M.; Dodoo, D.; Thesien, M. Antibody-dependent cellular inhibition is associated with reduced risk against febrile malaria in a longitudinal cohort study involving Ghanaian children. Open Forum Infect. Dis. 2015, 2, ofv044. [Google Scholar] [CrossRef] [Green Version]
- Oeuvray, C.; Theisen, M.; Rogier, C.; Trape, J.-F.; Jepsen, S.; Druilhe, P. Cytophilic immunoglobulin responses to Plasmodium falciparum glutamate-rich protein are correlated with protection against clinical malaria in Dielmo, Senegal. Infect. Immun. 2000, 68, 2617–2620. [Google Scholar] [CrossRef] [Green Version]
- Courtin, D.; Oesterholt, M.; Huismans, H.; Kusi, K.; Milet, J.; Badaut, C.; Gaye, O.; Roeffen, W.; Remarque, E.J.; Sauerwein, R.; et al. The quantity and quality of African children’s IgG responses to merozoite surface antigens reflect protection against Plasmodium falciparum malaria. PLoS ONE 2009, 4, e7590. [Google Scholar] [CrossRef] [Green Version]
- Sirima, S.B.; Tiono, A.B.; Ouédraogo, A.; Diarra, A.; Lin Ouédraogo, A.; Yaro, J.B.; Ouédraogo, E.; Gansané, A.; Bougouma, E.C.; Konaté, A.T.; et al. Safety and immunogenicity of the malaria vaccine candidate MSP3 long synthetic peptide in 12–24 months-old Burkinabe children. PLoS ONE 2009, 4, e7549. [Google Scholar] [CrossRef]
- Mazumdar, S.; Mukherjee, P.; Yazdani, S.S.; Jain, S.K.; Mohmmed, A.; Chauhan, V.S. Plasmodium falciparum merozoite surface protein 1 (MSP-1)-MSP-3 chimeric protein: Immunogenicity determined with human-compatible adjuvants and induction of protective immune response. Infect. Immun. 2010, 78, 872–883. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Mukherjee, P.; Dhawan, S.; Pandey, A.K.; Mazumdar, S.; Gaur, D.; Jain, S.K.; Chauhan, V.S. Production and preclinical evaluation of Plasmodium falciparum MSP-119 and MSP-311 chimeric protein, PfMSP-Fu24. Clin. Vaccine Immunol. 2014, 21, 886–897. [Google Scholar] [CrossRef] [Green Version]
- Coffman, R.L.; Sher, A.; Seder, R.A. Vaccine adjuvants: Putting innate immunity to work. Immunity 2010, 33, 492–503. [Google Scholar] [CrossRef] [Green Version]
- Magiri, R.; Mutwiri, G.; Wilson, H.L. Recent advances in experimental polyphosphazene adjuvants and their mechanisms of action. Cell Tissue Res. 2018, 374, 465–471. [Google Scholar] [CrossRef]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Miyaji, E.N.; Carvalho, E.; Oliveira, M.L.; Raw, I.; Ho, P.L. Trends in adjuvant development for vaccines: DAMPs and PAMPs as potential new adjuvants. Braz. J. Med. Biol. Res. 2011, 44, 500–513. [Google Scholar] [CrossRef]
- Liu, F.; Sun, X.; Fairman, J.; Lewis, D.B.; Katz, J.M.; Levine, M.; Tumpey, T.M.; Lu, X. A cationic liposome-DNA complexes adjuvant (JVRS-100) enhances the immunogenicity and cross-protective efficacy of pre-pandemic influenza A (H5N1) vaccine in ferrets. Virology 2016, 492, 197–203. [Google Scholar] [CrossRef]
- Rosenkrands, I.; Vingsbo-Lundberg, C.; Bundgaard, T.J.; Lindenstrøm, T.; Enouf, V.; van der Werf, S.; Andersen, P.; Agger, E.M. Enhanced humoral and cell-mediated immune responses after immunization with trivalent influenza vaccine adjuvanted with cationic liposomes. Vaccine 2011, 29, 6283–6291. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Kool, M.; Willart, M.A.; Hammad, H. Mechanism of action of clinically approved adjuvants. Curr. Opin. Immunol. 2009, 21, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Marrack, P.; McKee, A.S.; Munks, M.W. Towards an understanding of the adjuvant action of aluminium. Nat. Rev. Immunol. 2009, 9, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenbarth, S.C.; Colegio, O.R.; O’Connor, W.; Sutterwala, F.S.; Flavell, R.A. Crucial role for the Nalp3 inflammasome in the immunostimulatory properties of aluminium adjuvants. Nature 2008, 453, 1122–1126. [Google Scholar] [CrossRef]
- Black, K.A.; Lin, B.F.; Wonder, E.A.; Desai, S.S.; Chung, E.J.; Ulery, B.D.; Katari, R.S.; Tirrell, M.V. Biocompatibility and characterization of a peptide amphiphile hydrogel for applications in peripheral nerve regeneration. Tissue Eng. Part A 2015, 21, 1333–1342. [Google Scholar] [CrossRef]
- Azmi, F.; Ahmad Fuaad, A.A.; Skwarczynski, M.; Toth, I. Recent progress in adjuvant discovery for peptide-based subunit vaccines. Hum. Vaccines Immunother. 2014, 10, 778–796. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Cai, L.; Jia, Y.G.; Liu, S.; Chen, Y.; Ren, L. Progress in self-healing hydrogels assembled by host-guest interactions: Preparation and biomedical applications. J. Mater. Chem. B 2019, 7, 1637–1651. [Google Scholar] [CrossRef]
- Kobsa, S.; Saltzman, W.M. Bioengineering approaches to controlled protein delivery. Pediatr. Res. 2008, 63, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Panda, J.J.; Mishra, A.; Basu, A.; Chauhan, V.S. Stimuli responsive self-assembled hydrogel of a low molecular weight free dipeptide with potential for tunable drug delivery. Biomacromolecules 2008, 9, 2244–2250. [Google Scholar] [CrossRef] [PubMed]
- Panda, J.J.; Kaul, A.; Kumar, S.; Alam, S.; Mishra, A.K.; Kundu, G.C.; Chauhan, V.S. Modified dipeptide-based nanoparticles: Vehicles for targeted tumor drug delivery. Nanomedicine 2013, 8, 1927–1942. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Panda, J.J.; Chauhan, V.S. Novel dipeptide nanoparticles for effective curcumin delivery. Int. J. Nanomed. 2012, 7, 4207–4222. [Google Scholar]
- Khatri, A.; Mishra, A.; Chauhan, V.S. Characterization of DNA condensation by conformationally restricted dipeptides and gene delivery. J. Biomed. Nanotechnol. 2017, 13, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Bagaria, A.; Mishra, A.; Mathur, P.; Basu, A.; Ramakumar, S.; Chauhan, V.S. Self-assembly of a dipeptide-containing conformationally restricted dehydrophenylalanine residue to form ordered nanotubes. Adv. Mater. 2007, 19, 858–861. [Google Scholar] [CrossRef]
- Panda, J.J.; Dua, R.; Mishra, A.; Mittra, B.; Chauhan, V.S. 3D cell growth and proliferation on a RGD functionalized nanofibrillar hydrogel based on a conformationally restricted residue containing dipeptide. ACS Appl. Mater. Interfaces 2010, 2, 2839–2848. [Google Scholar] [CrossRef]
- Thota, C.K.; Yadav, N.; Chauhan, V.S. A novel highly stable and injectable hydrogel based on a conformationally restricted ultrashort peptide. Sci. Rep. 2016, 6, 31167. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, A.; Hayakawa, T.; Yamamoto, Y.; Hamori, M.; Tabata, K.; Seto, K.; Shibata, N. Controlled release of insulin from self-assembling nanofiber hydrogel, PuraMatrix™: Application for the subcutaneous injection in rats. Eur. J. Pharm. Sci. 2012, 45, 1–7. [Google Scholar] [CrossRef]
- Mazumdar, S.; Sachdeva, S.; Chauhan, V.S.; Yazdani, S.S. Identification of cultivation condition to produce correctly folded form of a malaria vaccine based on Plasmodium falciparum merozoite surface protein-1 in Escherichia coli. Bioprocess Biosyst. Eng. 2010, 33, 719–730. [Google Scholar] [CrossRef]
- Pattnaik, P.; Shakri, A.R.; Singh, S.; Goel, S.; Mukherjee, P.; Chitnis, C.E. Immunogenicity of a recombinant malaria vaccine based on receptor binding domain of Plasmodium falciparum EBA-175. Vaccine 2007, 25, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Trager, W.; Jensen, J.B. Human malaria parasites in continuous culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef]
- Pizarro, J.; Chitarra, V.; Verger, D.; Holm, I.; Pêtres, S.; Dartevelle, S.; Nato, F.; Longacre, S.; Bentley, G. Crystal structure of a Fab complex formed with PfMSP1–19, the C-terminal fragment of merozoite surface protein 1 from Plasmodium falciparum: A malaria vaccine candidate. J. Mol. Biol. 2003, 328, 1091–1103. [Google Scholar] [CrossRef]
- Chappel, J.A.; Holder, A.A. Monoclonal antibodies that inhibit Plasmodium falciparum invasion in vitro recognise the first growth factor-like domain of merozoite surface protein-1. Mol. Biochem. Parasitol. 1993, 60, 303–311. [Google Scholar] [CrossRef]
- Rudra, J.S.; Tian, Y.F.; Jung, J.P.; Collier, J.H. A self-assembling peptide acting as an immune adjuvant. Proc. Natl. Acad. Sci. USA 2010, 10, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Yang, P.; Huang, P.; Zhang, C.; Kong, D.; Wang, W. Injectable polypeptide hydrogel-based co-delivery of vaccine and immune checkpoint inhibitors improves tumor immunotherapy. Theranostics 2019, 9, 2299–2314. [Google Scholar] [CrossRef]
- Friedrich, B.M.; Beasley, D.W.; Rudra, J.S. Supramolecular peptide hydrogel adjuvanted subunit vaccine elicits protective antibody responses against West Nile virus. Vaccine 2016, 34, 5479–5482. [Google Scholar] [CrossRef]
- Karavasili, C.; Panteris, E.; Vizirianakis, I.S.; Koutsopoulos, S.; Fatouros, D.G. Chemotherapeutic delivery from a self-assembling peptide nanofiber hydrogel for the management of Glioblastoma. Pharm. Res. 2018, 35, 166. [Google Scholar] [CrossRef]
- Lee, A.L.Z.; Yang, C.; Gao, S.; Hedrick, J.L.; Yang, Y.Y. Subcutaneous vaccination using injectable biodegradable hydrogels for long-term immune response. Nanomedicine 2019, 21, 102056. [Google Scholar] [CrossRef]
- Zhang, H.; Park, J.; Jiang, Y.; Woodrow, K.A. Rational design of charged peptides that self-assemble into robust nanofibers as immune-functional scaffolds. Acta Biomater. 2017, 55, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Banquy, X.; Suarez, F.; Argaw, A.; Rabanel, J.-M.; Grutter, P.; Bouchard, J.-F.; Hildgen, P.; Giasson, S. Effect of mechanical properties of hydrogel nanoparticles on macrophage cell Uptake. Soft Matter 2009, 5, 3984–3991. [Google Scholar] [CrossRef]
- Egan, A.; Waterfall, M.; Pinder, M.; Holder, A.; Riley, E. Characterization of human T- and B-cell epitopes in the C terminus of Plasmodium falciparum merozoite surface protein 1: Evidence for poor T-cell recognition of polypeptides with numerous disulfide bonds. Infect. Immun. 1997, 65, 3024–3031. [Google Scholar] [CrossRef] [Green Version]
- Roth, G.A.; Gale, E.C.; Alcántara-Hernández, M.; Luo, W.; Axpe, E.; Verma, R.; Yin, Q.; Yu, A.C.; Hernandez, H.L.; Maikawa, C.L.; et al. Injectable hydrogels for sustained codelivery of subunit vaccines enhance humoral immunity. ACS Cent. Sci. 2020, 6, 1800–1812. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Liang, X.; Li, L.; Gong, S.; Li, X.; Zhang, M.; Zhu, S.; Xiao, H.; Wu, Q.; Gong, C. A spontaneously formed and self-adjuvanted hydrogel vaccine triggers strong immune responses. Mater. Des. 2021, 197, 109232. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anand, G.; Biswas, S.; Yadav, N.; Mukherjee, P.; Chauhan, V.S. Production and Immunogenicity of a Tag-Free Recombinant Chimera Based on PfMSP-1 and PfMSP-3 Using Alhydrogel and Dipeptide-Based Hydrogels. Vaccines 2021, 9, 782. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9070782
Anand G, Biswas S, Yadav N, Mukherjee P, Chauhan VS. Production and Immunogenicity of a Tag-Free Recombinant Chimera Based on PfMSP-1 and PfMSP-3 Using Alhydrogel and Dipeptide-Based Hydrogels. Vaccines. 2021; 9(7):782. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9070782
Chicago/Turabian StyleAnand, Gaurav, Saikat Biswas, Nitin Yadav, Paushali Mukherjee, and Virander Singh Chauhan. 2021. "Production and Immunogenicity of a Tag-Free Recombinant Chimera Based on PfMSP-1 and PfMSP-3 Using Alhydrogel and Dipeptide-Based Hydrogels" Vaccines 9, no. 7: 782. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9070782