
Efficacy of SAT2 Foot-and-Mouth Disease Vaccines Formulated with Montanide ISA 206B and Quil-A Saponin Adjuvants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Production of FMDV Antigen and Vaccine Formulation
2.3. Cattle Immunizations and Virus Challenge
2.4. Antibody Detection in Nguni Cattle
2.5. Non-Structural Protein (NSP) ELISA
2.6. Virus Neutralization Test (VNT)
2.7. IgG1 and IgG2 Isotyping ELISA
2.8. Whole Blood Re-Stimulation and Bovine Interferon Gamma (IFN-γ) ELISA
2.9. Virus Isolation
2.10. RNA Extraction, cDNA Synthesis and Real-Time Quantitative RT-PCR
2.11. Statistical Analysis
3. Results
3.1. Antibody Kinetics in Vaccinated Nguni Cattle with Stabilized SAT2 Antigen Formulated with Montanide ISA 206B or Quil-A Saponin Adjuvants
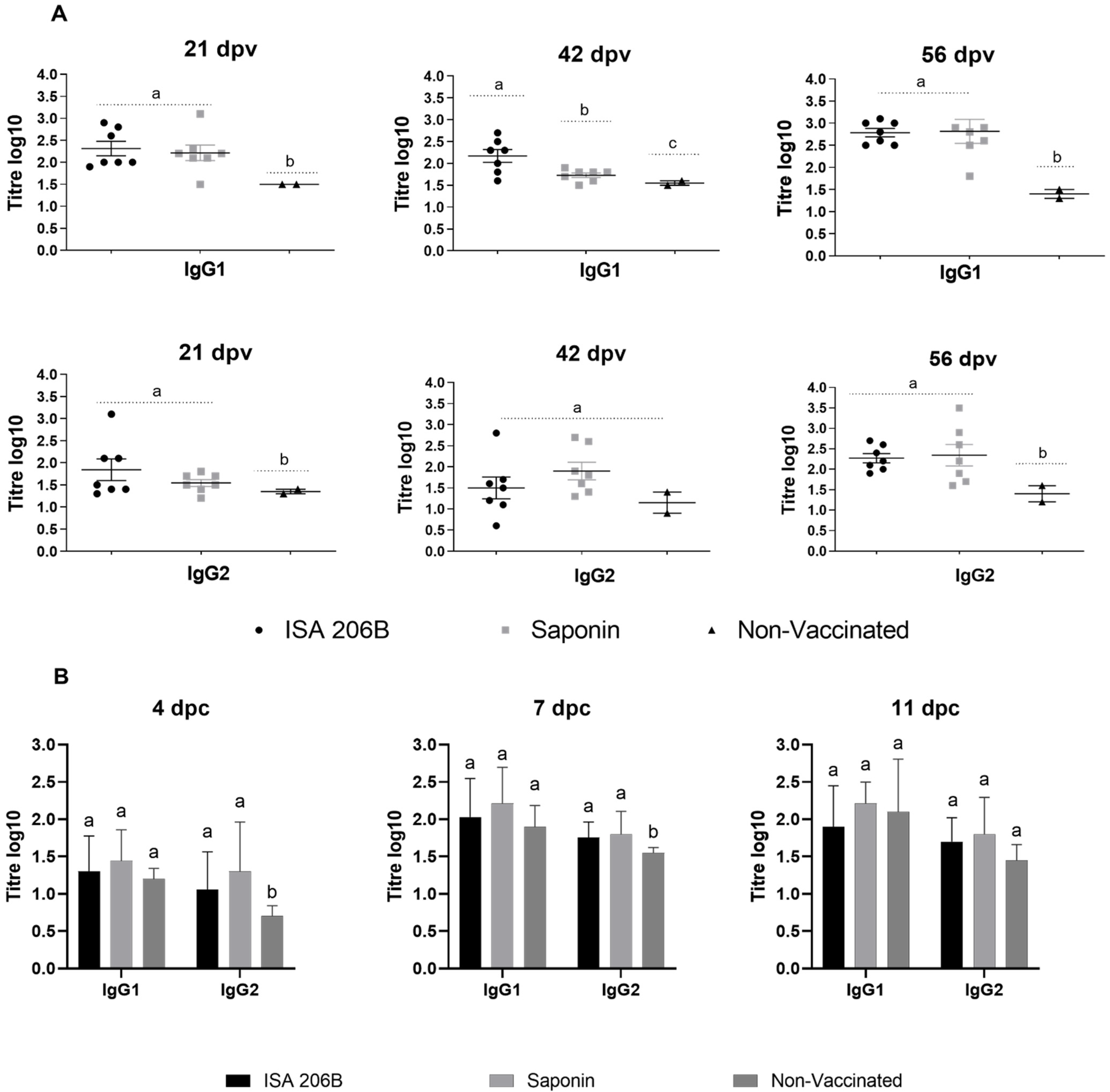
3.2. IgG Isotyping and IFN-γ Responses
3.3. Protection of Vaccinated Cattle against Live SAT2/ZIM/7/83 Virus Challenge
3.4. Virus Isolation and the Presence of Viral RNA Post Challenge
3.5. Variation in Response among Individuals to Antigenic Sites on the VP2 Protein Responsible for Antigenic Drift
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Organization for Animal Health (OIE). Terrestrial Animal Health Code; Office International des Epizooties: Paris, France, 2016. [Google Scholar]
- Bruckner, G.; Vosloo, W.; Du Plessis, B.; Kloeck, P.; Connoway, F.; Ekron, M.; Weaver, D.; Dickason, C.; Schreuder, F.; Marais, T.; et al. Foot and mouth disease: The experience of South Africa. OIE Rev. Sci. Tech. 2002, 21, 751–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, P. Vaccination as a means of control of foot-and-mouth disease in sub-saharan Africa. Vaccine 1998, 16, 261–264. [Google Scholar] [CrossRef]
- Ferguson, K.J.; Cleaveland, S.; Haydon, D.; Caron, A.; Kock, R.; Lembo, T.; Hopcraft, J.G.C.; Chardonnet, B.; Nyariki, T.; Keyyu, J.; et al. Evaluating the Potential for the Environmentally Sustainable Control of Foot and Mouth Disease in Sub-Saharan Africa. EcoHealth 2013, 10, 314–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jori, F.; Caron, A.; Thompson, P.N.; Dwarka, R.; Foggin, C.; De Garine-Wichatitsky, M.; Hofmeyr, M.; Van Heerden, J.; Heath, L.; Garine-Wichatitsky, M. Characteristics of Foot-and-Mouth Disease Viral Strains Circulating at the Wildlife/livestock Interface of the Great Limpopo Transfrontier Conservation Area. Transbound. Emerg. Dis. 2014, 63, e58–e70. [Google Scholar] [CrossRef] [Green Version]
- Bahnemann, H.G. Inactivation of Viral Antigens for Vaccine Preparation with Particular Reference to the Application of Binary Ethylenimine. Vaccine 1990, 8, 299–303. [Google Scholar] [CrossRef]
- Doel, T. FMD vaccines. Virus Res. 2003, 91, 81–99. [Google Scholar] [CrossRef]
- Sutmoller, P.; Barteling, S.S.; Olascoaga, R.C.; Sumption, K.J. Control and eradication of foot-and-mouth disease. Virus Res. 2003, 91, 101–144. [Google Scholar] [CrossRef]
- World Organization for Animal Health (OIE). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2018, 8th ed.; Office International des Epizooties: Paris, France, 2018. [Google Scholar]
- Doel, T.R.; Baccarini, P.J. Thermal stability of foot-and-mouth disease virus. Arch. Virol. 1981, 70, 21–32. [Google Scholar] [CrossRef]
- Kotecha, A.; Seago, J.; Scott, K.A.; Burman, A.; Loureiro, S.; Ren, J.; Porta, C.; Ginn, H.M.; Jackson, T.; Perez-Martin, E.; et al. Structure-based energetics of protein interfaces guides foot-and-mouth disease virus vaccine design. Nat. Struct. Mol. Biol. 2015, 22, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.A.; Kotecha, A.; Seago, J.; Ren, J.; Fry, E.E.; Stuart, D.I.; Charleston, B.; Maree, F.F. SAT2 Foot-and-Mouth Disease Virus Structurally Modified for Increased Thermostability. J. Virol. 2017, 91, e02312–e02316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieder, E.; Baxt, B.; Lubroth, J.; Mason, P.W. Vaccines prepared from chimeras of foot-and-mouth disease virus (FMDV) induce neutralizing antibodies and protective immunity to multiple serotypes of FMDV. J. Virol. 1994, 68, 7092–7098. [Google Scholar] [CrossRef] [Green Version]
- Blignaut, B.; Visser, N.; Theron, J.; Rieder, E.; Maree, F.F. Custom-engineered chimeric foot-and-mouth disease vaccine elicits protective immune responses in pigs. J. Gen. Virol. 2010, 92, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Maree, F.F.; Nsamba, P.; Mutowembwa, P.; Rotherham, L.S.; Esterhuysen, J.J.; Scott, K.A. Intra-serotype SAT2 chimeric foot-and-mouth disease vaccine protects cattle against FMDV challenge. Vaccine 2015, 33, 2909–2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maree, F.F.; Blignaut, B.; de Beer, T.A.; Visser, N.; Rieder, E.A. Mapping of amino acid residues responsible for adhesion of cell culture-adapted foot-and-mouth disease SAT type viruses. Virus Res. 2010, 153, 82–91. [Google Scholar] [CrossRef]
- Barnard, A.L.; Arriens, A.; Cox, S.; Barnett, P.; Kristensen, B.; Summerfield, A.; McCullough, K.C. Immune response characteristics following emergency vaccination of pigs against foot-and-mouth disease. Vaccine 2005, 23, 1037–1047. [Google Scholar] [CrossRef]
- Barnett, P.; Pullen, L.; Williams, L.; Doel, T. International bank for foot-and-mouth disease vaccine: Assessment of Montanide ISA 25 and ISA 206, two commercially available oil adjuvants. Vaccine 1996, 14, 1187–1198. [Google Scholar] [CrossRef]
- Patil, P.; Bayry, J.; Ramakrishna, C.; Hugar, B.; Misra, L.; Natarajan, C. Immune responses of goats against foot-and-mouth disease quadrivalent vaccine: Comparison of double oil emulsion and aluminium hydroxide gel vaccines in eliciting immunity. Vaccine 2002, 20, 2781–2789. [Google Scholar] [CrossRef]
- Cloete, M.; Dungu, B.; Van Staden, L.; Ismail-Cassim, N.; Vosloo, W. Evaluation of different adjuvants for foot-and-mouth disease vaccine containing all the SAT serotypes. Onderstepoort J. Vet. Res. 2008, 75, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Park, M.-E.; Lee, S.-Y.; Kim, R.-H.; Ko, M.-K.; Lee, K.-N.; Kim, S.-M.; Kim, B.-K.; Lee, J.-S.; Kim, B.; Park, J.-H. Enhanced immune responses of foot-and-mouth disease vaccine using new oil/gel adjuvant mixtures in pigs and goats. Vaccine 2014, 32, 5221–5227. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P. The performance of Southern African Territories serotypes of foot and mouth disease antigen in oil-adjuvanted vaccines. Rev. Sci. Tech. 1996, 15, 913–922. [Google Scholar] [CrossRef]
- Li, D.; Zhou, C.; She, D.; Li, P.; Sun, P.; Bai, X.; Chen, Y.; Xie, B.; Liu, Z. The comparison of the efficacy of swine FMD vaccine emulsified with oil adjuvant of ISA 201 VG or ISA 206 VG. J. Biosci. Med. 2013, 01, 22–25. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.C.; Coulter, A.R. Adjuvants—A classification and review of their modes of action. Vaccine 1997, 15, 248–256. [Google Scholar] [CrossRef]
- Dar, P.; Kalaivanan, R.; Sied, N.; Mamo, B.; Kishore, S.; Suryanarayana, V.; Kondabattula, G. Montanide ISA™ 201 adjuvanted FMD vaccine induces improved immune responses and protection in cattle. Vaccine 2013, 31, 3327–3332. [Google Scholar] [CrossRef]
- Bazid, A.-H.I.; El-Alfy, H.A.; El-Didamony, G.; Elfeil, W.K.; El-Sayed, M.M.; Fawzy, M. Adjuvant effect of saponin in an oil-based monovalent (serotype O) foot-and-mouth disease virus vaccine on the antibody response in guinea pigs and cattle. Arch. Virol. 2021, 166, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Dalsgaard, K.; Jensen, M.H. Saponin Adjuvants. Vi. The Adjuvant Activity of Quil a in Trivalent Vaccination of Cattle and Guinea Pigs Against Foot-And-Mouth Disease. Acta Vet. Scand. 1977, 18, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Doel, T.; Püllen, L. International bank for foot-and-mouth disease vaccine: Stability studies with virus concentrates and vaccines prepared from them. Vaccine 1990, 8, 473–478. [Google Scholar] [CrossRef]
- Storey, P.; Theron, J.; Maree, F.F.; O’Neill, H. A second RGD motif in the 1D capsid protein of a SAT1 type foot-and-mouth disease virus field isolate is not essential for attachment to target cells. Virus Res. 2007, 124, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maree, F.F.; Blignaut, B.; Esterhuysen, J.J.; de Beer, T.; Theron, J.; O’Neill, H.G.; Rieder, E. Predicting antigenic sites on the foot-and-mouth disease virus capsid of the South African Territories types using virus neutralization data. J. Gen. Virol. 2011, 92, 2297–2309. [Google Scholar] [CrossRef]
- Opperman, P.; Rotherham, L.S.; Esterhuysen, J.; Charleston, B.; Juleff, N.; Capozzo, A.V.; Theron, J.; Maree, F.F. Determining the Epitope Dominance on the Capsid of a Serotype SAT2 Foot-and-Mouth Disease Virus by Mutational Analyses. J. Virol. 2014, 88, 8307–8318. [Google Scholar] [CrossRef] [Green Version]
- Bahnemann, H.G. Binary ethylenimine as an inactivant for foot-and-mouth disease virus and its application for vaccine production. Arch. Virol. 1975, 47, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Barteling, S.J.; Meloen, R.H. A simple method for the quantification of 140 s particles of foot-and-mouth disease virus (FMDV). Arch. Gesamte Virusforsch. 1974, 45, 362–364. [Google Scholar] [CrossRef]
- Doel, T.; Mowat, G. An international collaborative study on foot and mouth disease virus assay methods. Quantification of 146S particles. J. Biol. Stand. 1985, 13, 335–344. [Google Scholar] [CrossRef]
- Scott, K.A.; Rathogwa, N.; Capozzo, A.; Maree, F.F. Evaluation of immune responses of stabilised SAT2 antigens of foot-and-mouth disease in cattle. Vaccine 2017, 35, 5426–5433. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, K.J.; Madsen, K.G.; Madsen, E.S.; Salt, J.S.; Nqindi, J.; Mackay, D.K.J. Differentiation of infection from vaccination in foot-and-mouth disease by the detection of antibodies to the non-structural proteins 3D, 3AB and 3ABC in ELISA using antigens expressed in baculovirus. Arch. Virol. 1998, 143, 1461–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn Schmiedeberg’s Arch. Pharmacol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Capozzo, A.; Periolo, O.; Robiolo, B.; Seki, C.; La Torre, J.; Grigera, P. Total and isotype humoral responses in cattle vaccinated with foot and mouth disease virus (FMDV) immunogen produced either in bovine tongue tissue or in BHK-21 cell suspension cultures. Vaccine 1997, 15, 624–630. [Google Scholar] [CrossRef]
- Bucafusco, D.; Di Giacomo, S.; Pega, J.; Schammas, J.M.; Cardoso, N.; Capozzo, A.V.; Perez-Filgueira, M. Foot-and-mouth disease vaccination induces cross-reactive IFN-γ responses in cattle that are dependent on the integrity of the 140S particles. Virology 2015, 476, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Doherty, R.; O’Farrelly, C.; Meade, K. Epigenetic regulation of the innate immune response to LPS in bovine peripheral blood mononuclear cells (PBMC). Vet. Immunol. Immunopathol. 2013, 154, 102–110. [Google Scholar] [CrossRef]
- Ferris, N.; Donaldson, A. The world reference laboratory for Foot and Mouth Disease: A review of thirty-three years of activity (1958–1991). Rev. Sci. Tech. 1992, 11, 657–684. [Google Scholar] [CrossRef]
- Roeder, P.; Smith, P.L.B. Detection and typing of foot-and-mouth disease virus by enzyme-linked immunosorbent assay: A sensitive, rapid and reliable technique for primary diagnosis. Res. Vet. Sci. 1987, 43, 225–232. [Google Scholar] [CrossRef]
- Callahan, J.D.; Brown, F.; Osorio, F.A.; Sur, J.-H.; Kramer, E.; Long, G.W.; Lubroth, J.; Ellis, S.J.; Shoulars, K.S.; Gaffney, K.L.; et al. Use of a portable real-time reverse transcriptasepolymerase chain reaction assay for rapid detection of foot-and-mouth disease virus. J. Am. Vet. Med. Assoc. 2002, 220, 1636–1642. [Google Scholar] [CrossRef] [PubMed]
- Cox, S. Emergency vaccination of sheep against foot-and-mouth disease: Protection against disease and reduction in contact transmission. Vaccine 1999, 17, 1858–1868. [Google Scholar] [CrossRef]
- Ibrahim, E.E.-S.; Gamal, W.M.; Hassan, A.I.; Mahdy, S.E.-D.; Hegazy, A.Z.; Abdel-Atty, M.M. Comparative study on the immunopotentiator effect of ISA 201, ISA 61, ISA 50, ISA 206 used in trivalent foot and mouth disease vaccine. Vet. World 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Coulter, A.; Wong, T.-Y.; Drane, D.; Bates, J.; Macfarlan, R.; Cox, J. Studies on experimental adjuvanted influenza vaccines: Comparison of immune stimulating complexes (Iscoms™) and oil-in-water vaccines. Vaccine 1998, 16, 1243–1253. [Google Scholar] [CrossRef]
- Pay, T.; Hingley, P. Correlation of 140S antigen dose with the serum neutralizing antibody response and the level of protection induced in cattle by foot-and-mouth disease vaccines. Vaccine 1987, 5, 60–64. [Google Scholar] [CrossRef]
- McCullough, K.C.; De Simone, F.; Brocchi, E.; Capucci, L.; Crowther, J.R.; Kihm, U. Protective immune response against foot-and-mouth disease. J. Virol. 1992, 66, 1835–1840. [Google Scholar] [CrossRef] [Green Version]
- Salt, J. The carrier state in foot and mouth disease—An immunological review. Br. Vet. J. 1993, 149, 207–223. [Google Scholar] [CrossRef]
- Barnett, P.V.; Statham, R.J.; Vosloo, W.; Haydon, D. Foot-and-mouth disease vaccine potency testing: Determination and statistical validation of a model using a serological approach. Vaccine 2003, 21, 3240–3248. [Google Scholar] [CrossRef]
- Patch, J.R.; Pedersen, L.E.; Toka, F.N.; Moraes, M.; Grubman, M.J.; Nielsen, M.; Jungersen, G.; Buus, S.; Golde, W.T. Induction of Foot-and-Mouth Disease Virus-Specific Cytotoxic T Cell Killing by Vaccination. Clin. Vaccine Immunol. 2010, 18, 280–288. [Google Scholar] [CrossRef] [Green Version]
- Schiller, I.; Waters, W.R.; Vordermeier, H.M.; Nonnecke, B.; Welsh, M.; Keck, N.; Whelan, A.; Sigafoose, T.; Stamm, C.; Palmer, M.; et al. Optimization of a Whole-Blood Gamma Interferon Assay for Detection of Mycobacterium bovis-infected cattle. Clin. Vaccine Immunol. 2009, 16, 1196–1202. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.; Fleming, L.; Statham, B.; Hamblin, P.; Barnett, P.; Paton, D.J.; Park, J.-H.; Joo, Y.S.; Parida, S. Interferon-γ Induced by In Vitro Re-Stimulation of CD4+ T-Cells Correlates with In Vivo FMD Vaccine Induced Protection of Cattle against Disease and Persistent Infection. PLoS ONE 2012, 7, e44365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, B.V.; Lefevre, E.A.; Windsor, M.A.; Inghese, C.; Gubbins, S.; Prentice, H.; Juleff, N.D.; Charleston, B. CD4+ T-cell responses to foot-and-mouth disease virus in vaccinated cattle. J. Gen. Virol. 2013, 94, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Vosloo, W.; Bastos, A.; Sangare, O.; Hargreaves, S.; Thomson, G. Review of the status and control of foot and mouth disease in sub-Saharan Africa. OIE Rev. Sci. Tech. 2002, 21, 437–449. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rathogwa, N.M.; Scott, K.A.; Opperman, P.; Theron, J.; Maree, F.F. Efficacy of SAT2 Foot-and-Mouth Disease Vaccines Formulated with Montanide ISA 206B and Quil-A Saponin Adjuvants. Vaccines 2021, 9, 996. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9090996
Rathogwa NM, Scott KA, Opperman P, Theron J, Maree FF. Efficacy of SAT2 Foot-and-Mouth Disease Vaccines Formulated with Montanide ISA 206B and Quil-A Saponin Adjuvants. Vaccines. 2021; 9(9):996. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9090996
Chicago/Turabian StyleRathogwa, Ntungufhadzeni M., Katherine A. Scott, Pamela Opperman, Jacques Theron, and Francois F. Maree. 2021. "Efficacy of SAT2 Foot-and-Mouth Disease Vaccines Formulated with Montanide ISA 206B and Quil-A Saponin Adjuvants" Vaccines 9, no. 9: 996. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9090996