
Baker’s Yeast-Based Microbial Fuel Cell Mediated by 2-Methyl-1,4-Naphthoquinone

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. The Preparation of Yeast Samples
2.3. Graphite Electrode Preparation
2.4. The Preparation of Cells for Viability Assays
2.5. Electrochemical Measurements
2.6. Calculations
3. Results and Discussion
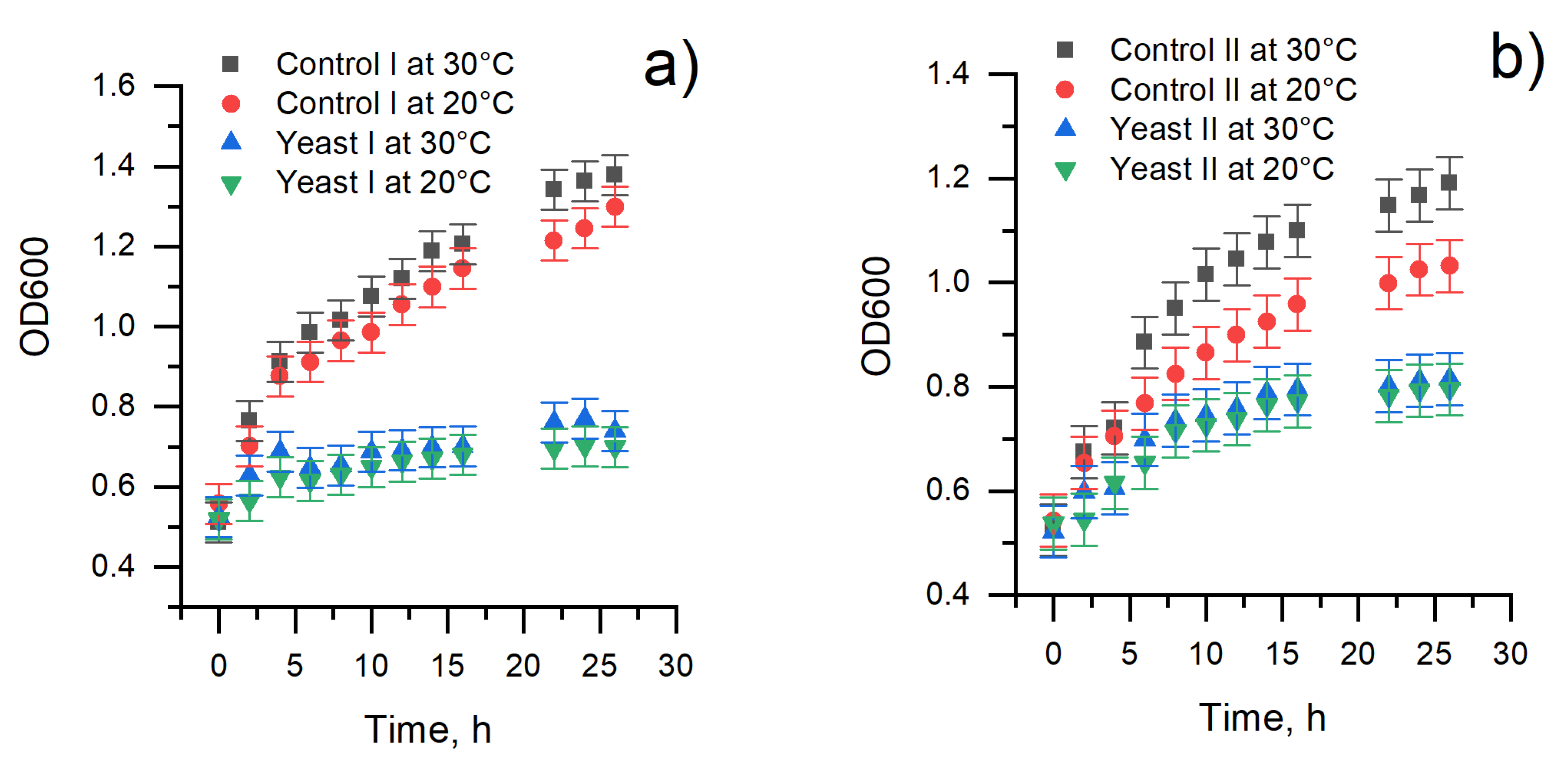
3.1. The Assessment of Yeast Cells’ Viability
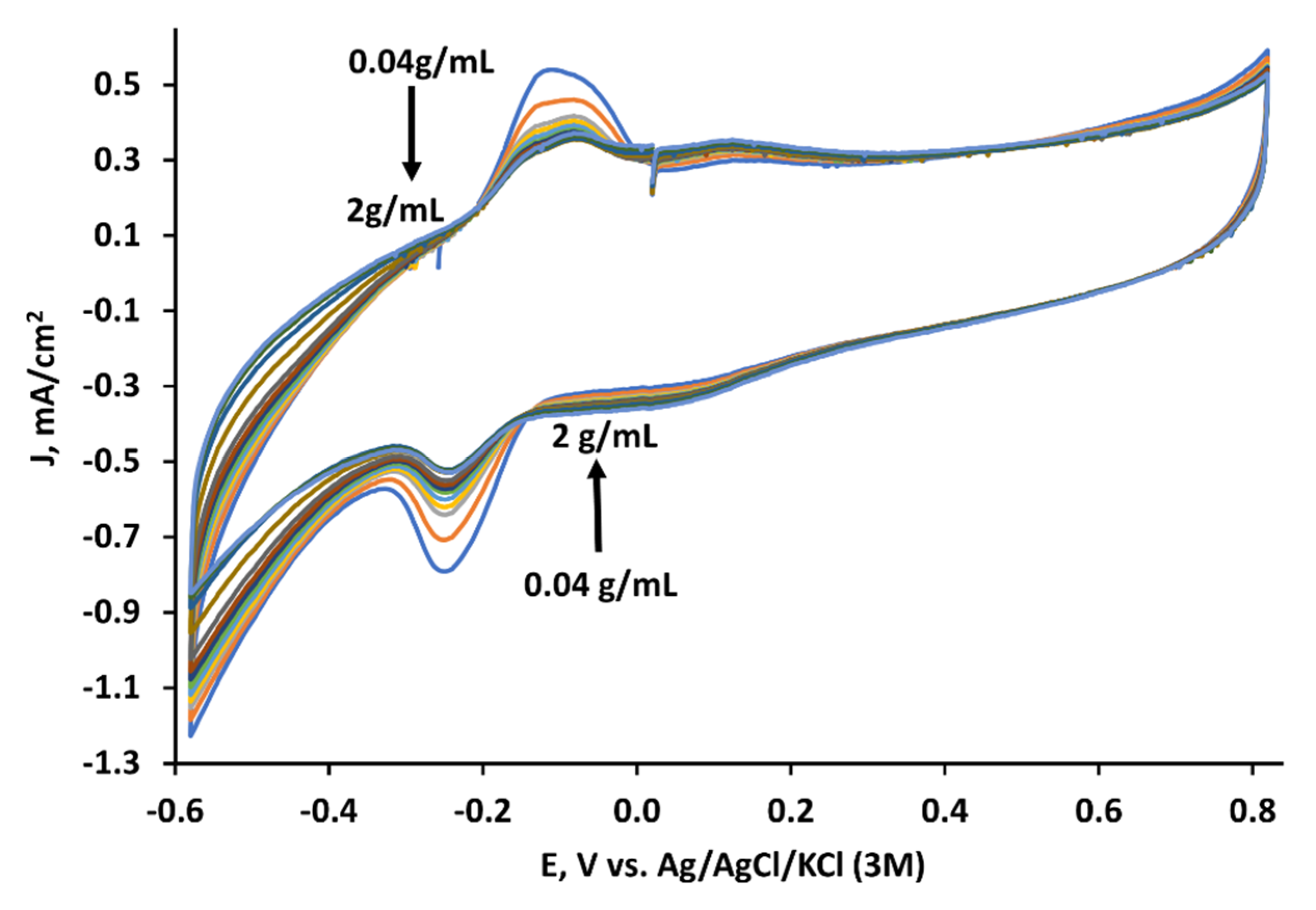
3.2. The Evaluation of the MD-Modified Graphite Electrode at Different Yeast Concentrations
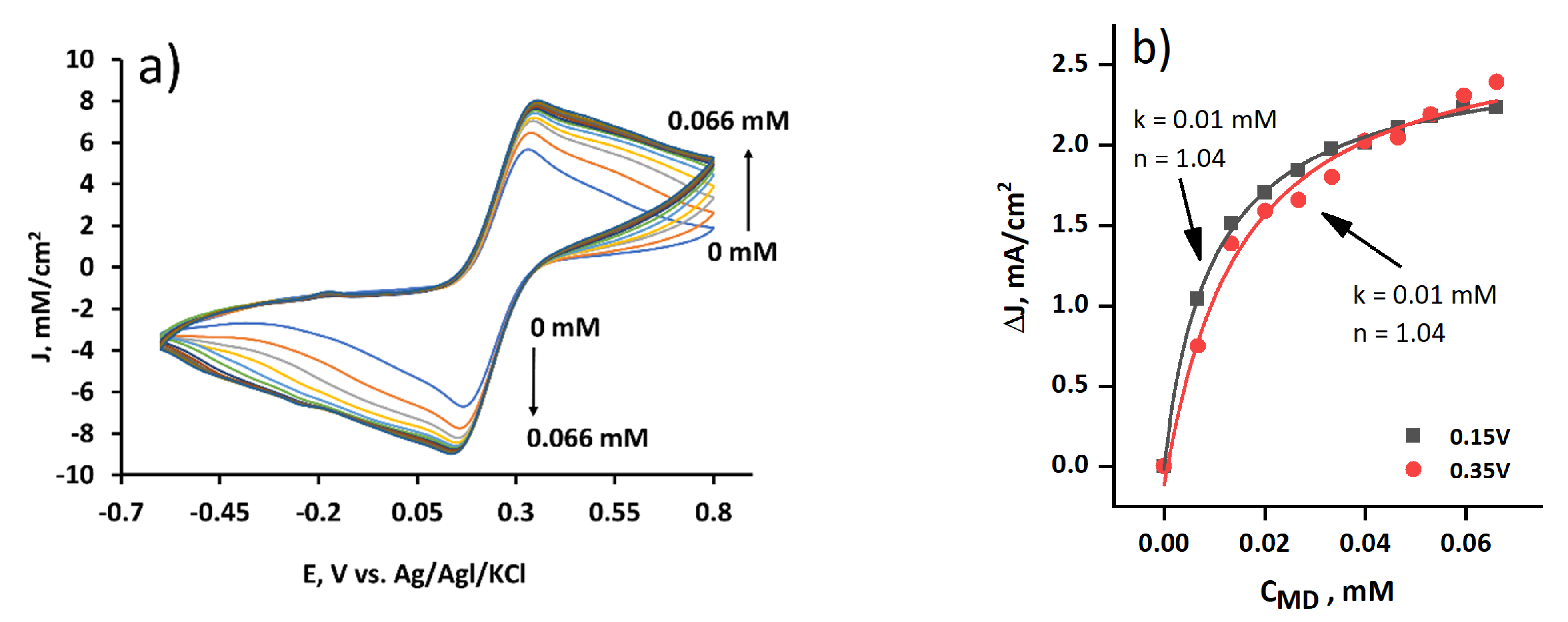
3.3. The Evaluation of the Yeast-Modified Graphite Electrode at Different MD Concentrations
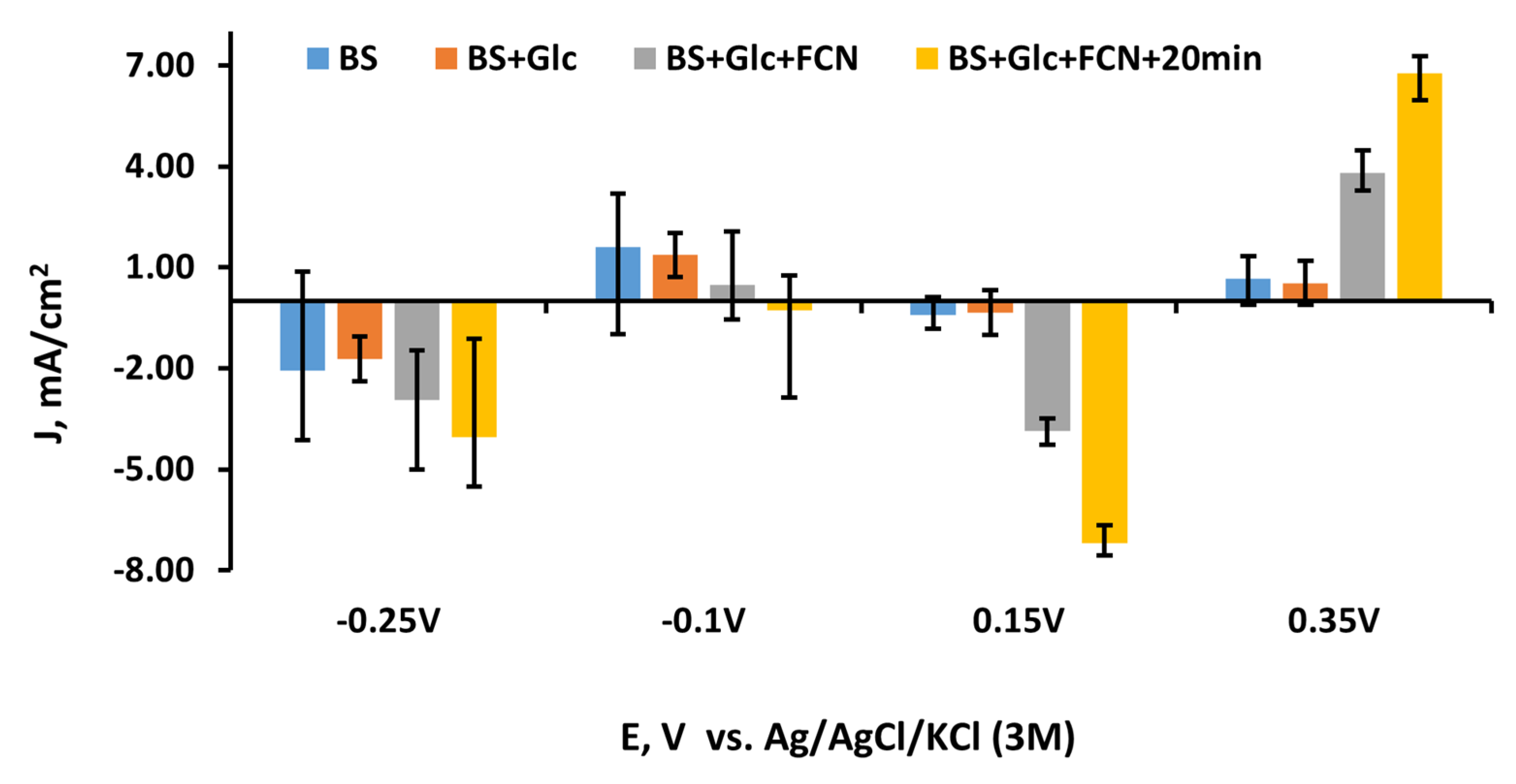
3.4. The Evaluation of the Yeast and MD-Modified Graphite Electrode at Different MD Concentrations
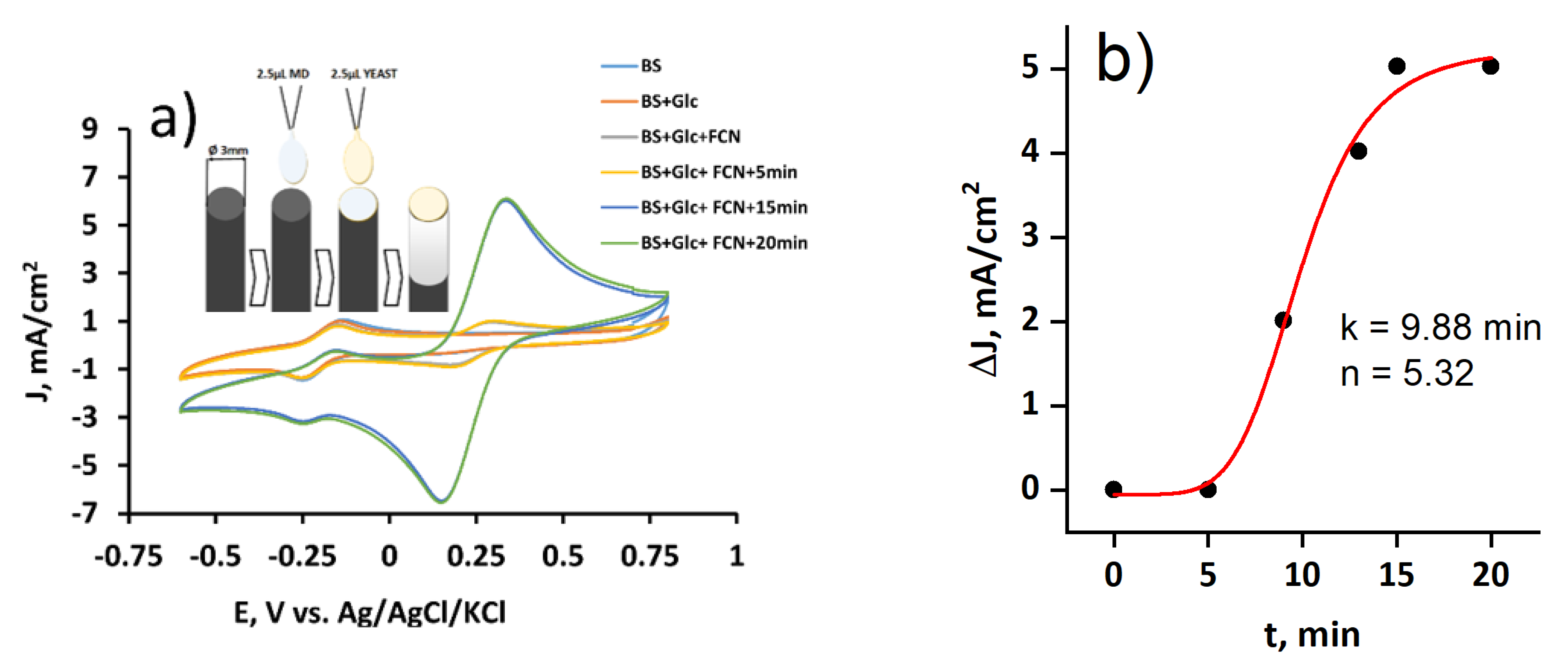
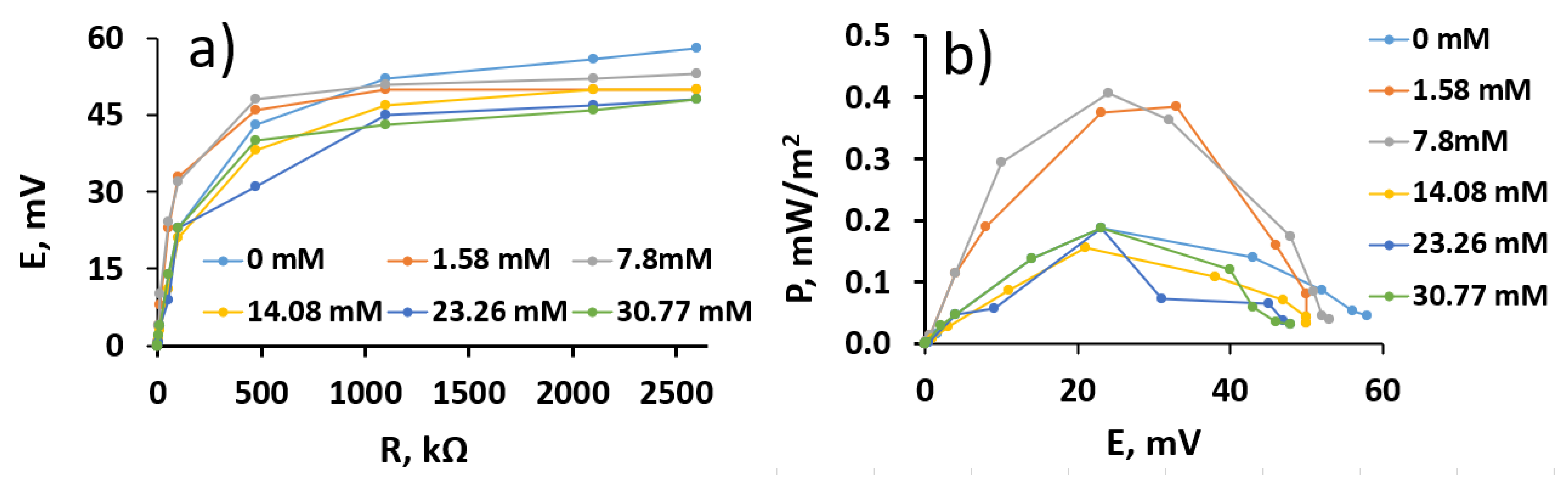
3.5. The Performance of the Microbial Fuel Cell
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahimnejad, M.; Adhami, A.; Darvari, S.; Zirepour, A.; Oh, S.E. Microbial fuel cell as new technology for bioelectricity generation: A review. Alex. Eng. J. 2015, 54, 745–756. [Google Scholar] [CrossRef] [Green Version]
- Rahimnejad, M.; Ghoreyshi, A.A.; Najafpour, G.; Jafary, T. Power generation from organic substrate in batch and continuous flow microbial fuel cell operations. Appl. Energy 2011. [Google Scholar] [CrossRef]
- Davis, F.; Higson, S.P.J. Biofuel cells-Recent advances and applications. Biosens. Bioelectron. 2007. [Google Scholar] [CrossRef] [Green Version]
- Slate, A.J.; Whitehead, K.A.; Brownson, D.A.C.; Banks, C.E. Microbial fuel cells: An overview of current technology. Renew. Sustain. Energy Rev. 2019, 101, 60–81. [Google Scholar] [CrossRef]
- Bruzaite, I.; Rozene, J.; Morkvenaite-Vilkonciene, I.; Ramanavicius, A. Towards microorganism-based biofuel cells: The viability of saccharomyces cerevisiae modified by multiwalled carbon nanotubes. Nanomaterials 2020, 10, 954. [Google Scholar] [CrossRef]
- Rozene, J.; Morkvenaite-Vilkonciene, I.; Bruzaite, I.; Dzedzickis, A.; Ramanavicius, A. Yeast-based microbial biofuel cell mediated by 9,10-phenantrenequinone. Electrochim. Acta 2021, 137918. [Google Scholar] [CrossRef]
- Pandey, P.; Shinde, V.N.; Deopurkar, R.L.; Kale, S.P.; Patil, S.A.; Pant, D. Recent advances in the use of different substrates in microbial fuel cells toward wastewater treatment and simultaneous energy recovery. Appl. Energy 2016, 168, 706–723. [Google Scholar] [CrossRef]
- Babanova, S.; Hubenova, Y.; Mitov, M. Influence of artificial mediators on yeast-based fuel cell performance. J. Biosci. Bioeng. 2011, 112, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Sharma, Y.; Li, B. The variation of power generation with organic substrates in single-chamber microbial fuel cells (SCMFCs). Bioresour. Technol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Li, C.M.; Bao, S.J.; Bao, Q.L. Carbon nanotube/polyaniline composite as anode material for microbial fuel cells. J. Power Sources 2007. [Google Scholar] [CrossRef]
- Hao Yu, E.; Scott, K. Enzymatic Biofuel Cells—Fabrication of Enzyme Electrodes. Energies 2010, 3, 23–42. [Google Scholar] [CrossRef] [Green Version]
- Scott, K.; Rimbu, G.A.; Katuri, K.P.; Prasad, K.K.; Head, I.M. Application of modified carbon anodes in microbial fuel cells. Process Saf. Environ. Prot. 2007. [Google Scholar] [CrossRef]
- Morkvenaite-Vilkonciene, I.; Ramanaviciene, A.; Ramanavicius, A. 9,10-Phenanthrenequinone as a redox mediator for the imaging of yeast cells by scanning electrochemical microscopy. Sens. Actuators B Chem. 2016, 228. [Google Scholar] [CrossRef]
- Kisieliute, A.; Popov, A.; Apetrei, R.-M.; Cârâc, G.; Morkvenaite-Vilkonciene, I.; Ramanaviciene, A.; Ramanavicius, A. Towards microbial biofuel cells: Improvement of charge transfer by self-modification of microoganisms with conducting polymer – Polypyrrole. Chem. Eng. J. 2019, 356, 1014–1021. [Google Scholar] [CrossRef]
- Petroniene, J.; Morkvenaite-Vilkonciene, I.; Miksiunas, R.; Bironaite, D.; Ramanaviciene, A.; Rucinskas, K.; Janusauskas, V.; Ramanavicius, A. Scanning electrochemical microscopy for the investigation of redox potential of human myocardium-derived mesenchymal stem cells grown at 2D and 3D conditions. Electrochim. Acta 2020, 360. [Google Scholar] [CrossRef]
- Petroniene, J.; Morkvenaite-Vilkonciene, I.; Miksiunas, R.; Bironaite, D.; Ramanaviciene, A.; Mikoliunaite, L.; Kisieliute, A.; Rucinskas, K.; Janusauskas, V.; Plikusiene, I.; et al. Evaluation of redox activity of human myocardium-derived mesenchymal stem cells by scanning electrochemical microscopy. Electroanalysis 2020, elan.201900723. [Google Scholar] [CrossRef]
- Rodriguez, C.E.; Sobol, Z.; Schiestl, R.H. 9,10-Phenanthrenequinone induces DNA deletions and forward mutations via oxidative mechanisms in the yeast Saccharomyces cerevisiae. Toxicol. In Vitro 2008, 22, 296–300. [Google Scholar] [CrossRef]
- Yamashoji, S. Different characteristics between menadione and menadione sodium bisulfite as redox mediator in yeast cell suspension. Biochem. Biophys. Rep. 2016, 6, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Hubenova, Y.V.; Rashkov, R.S.; Buchvarov, V.D.; Arnaudova, M.H.; Babanova, S.M.; Mitov, M.Y. Improvement of Yeast− Biofuel Cell Output by Electrode Modifications. Ind. Eng. Chem. Res. 2010, 50, 557–564. [Google Scholar] [CrossRef]
- O’Brien, P.J. Molecular mechanisms of quinone cytotoxicity. Chem. Biol. Interact. 1991, 80, 1–41. [Google Scholar] [CrossRef]
- Brunmark, A.; Cadenas, E. Redox and addition chemistry of quinoid compounds and its biological implications. Free Radic. Biol. Med. 1989, 7, 435–477. [Google Scholar] [CrossRef]
- Monks, T.J.; Lau, S.S. Toxicology of quinone-thioethers. Crit. Rev. Toxicol. 1992, 22, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Bolton, J.L.; Trush, M.A.; Penning, T.M.; Dryhurst, G.; Monks, T.J. Role of quinones in toxicology. Chem. Res. Toxicol. 2000, 13, 135–160. [Google Scholar] [CrossRef]
- Kumagai, Y.; Koide, S.; Taguchi, K.; Endo, A.; Nakai, Y.; Yoshikawa, T.; Shimojo, N. Oxidation of proximal protein sulfhydryls by phenanthraquinone, a component of diesel exhaust particles. Chem. Res. Toxicol. 2002, 15, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, O.V.; Bayliak, M.M.; Korobova, O.V.; Levine, R.L.; Lushchak, V.I. Buffer modulation of menadione-induced oxidative stress in Saccharomyces cerevisiae. Redox Rep. 2009, 14, 214–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, F.A.V.; Mariani, D.; Panek, A.D.; Eleutherio, E.C.A.; Pereira, M.D. Cytotoxicity Mechanism of Two Naphthoquinones (Menadione and Plumbagin) in Saccharomyces cerevisiae. PLoS ONE 2008, 3, e3999. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rozene, J.; Morkvenaite-Vilkonciene, I.; Bruzaite, I.; Zinovicius, A.; Ramanavicius, A. Baker’s Yeast-Based Microbial Fuel Cell Mediated by 2-Methyl-1,4-Naphthoquinone. Membranes 2021, 11, 182. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11030182
Rozene J, Morkvenaite-Vilkonciene I, Bruzaite I, Zinovicius A, Ramanavicius A. Baker’s Yeast-Based Microbial Fuel Cell Mediated by 2-Methyl-1,4-Naphthoquinone. Membranes. 2021; 11(3):182. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11030182
Chicago/Turabian StyleRozene, Juste, Inga Morkvenaite-Vilkonciene, Ingrida Bruzaite, Antanas Zinovicius, and Arunas Ramanavicius. 2021. "Baker’s Yeast-Based Microbial Fuel Cell Mediated by 2-Methyl-1,4-Naphthoquinone" Membranes 11, no. 3: 182. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11030182