
Association of Alpha-Crystallin with Fiber Cell Plasma Membrane of the Eye Lens Accompanied by Light Scattering and Cataract Formation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Binding of α-Crystallin to Lipids (Phospholipids (PLs) and Sphingolipids) of the Eye Lens Fiber Cell Plasma Membrane
2.1. Nature of α-Crystallin Binding to Lipids (PLs and Sphingolipids) of the Lens Membrane
2.2. Binding of α-Crystallin to the Lens Lipid Membrane Changes the Membrane’s Physical Properties
3. Role of Cholesterol (Chol) on the α-Crystallin Binding to the Lens Membrane
3.1. Extremely High Chol Content Forming Cholesterol Bilayer Domains (CBDs) within the Lens Membrane
3.2. Chol and CBDs Inhibit the Binding of α-Crystallin to the Membranes Made of Eye Lens Lipids (PLs and Sphingolipids)
3.3. Binding of α-Crystallin to the Chol/Lipid Membrane Changes the Membrane’s Physical Properties
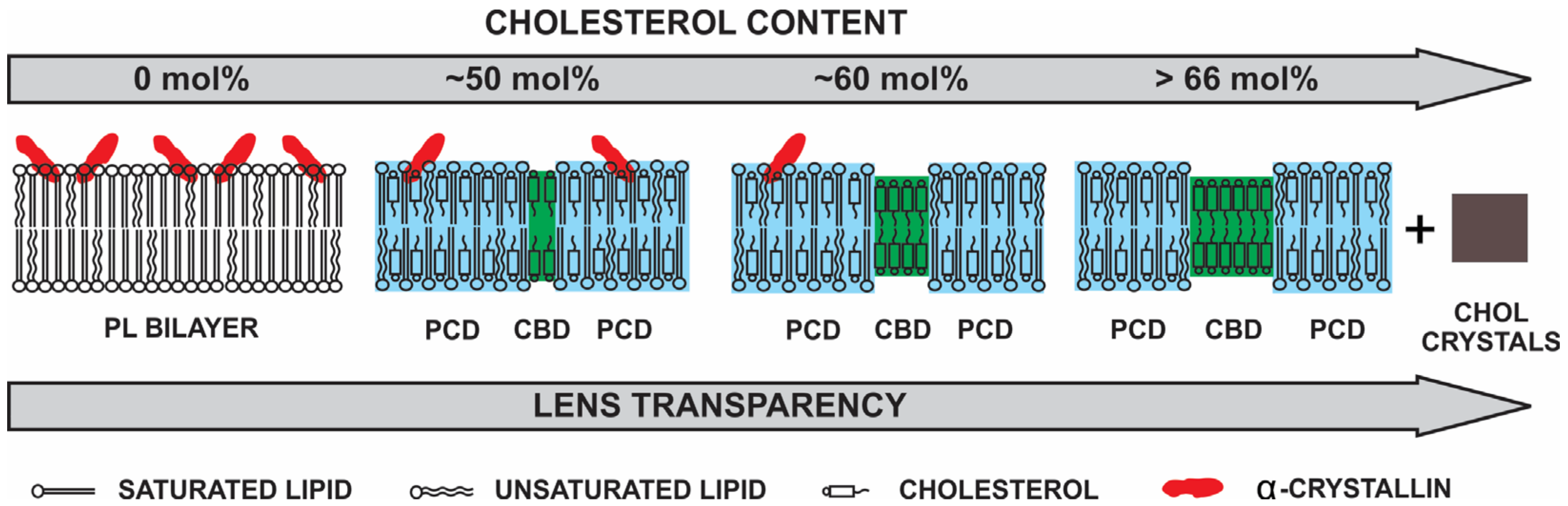
3.4. Hight Chol Content and Lens Transparency
3.5. Lipid Peroxidation and Cataract Formation
4. Interaction of α-Crystallin with the Lens Integral Membrane Proteins
5. Mutations and Post-Translational Modifications (PTMs) of α-Crystallin, Resulting in Association of α-Crystallin to the Fiber Cell Plasma Membranes and Cataract Formation
5.1. Mutations in α-Crystallin That Cause Association of α-Crystallin to Lens Membranes
5.2. Post-Translations Modifications (PTMs) in α-Crystallin
5.2.1. Deamidation
5.2.2. Phosphorylation
5.2.3. Isomerization
5.2.4. Acetylation
5.2.5. Glycation
5.2.6. Proteolytic Cleavage
5.2.7. Oxidation
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barbazetto, I.; Liang, J.; Chang, S.; Zheng, L.; Spector, A.; Dillon, J.P. Oxygen tension in the rabbit lens and vitreous before and after vitrectomy. Exp. Eye Res. 2004, 78, 917–924. [Google Scholar] [CrossRef]
- Borchman, D.; Yappert, M.C. Lipids and the ocular lens. J. Lipid Res. 2010, 51, 2473–2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassnett, S.; Shi, Y.; Vrensen, G.F.J.M. Biological glass: Structural determinants of eye lens transparency. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1250–1264. [Google Scholar] [CrossRef] [Green Version]
- Borchman, D.; Delamere, N.; McCauley, L.; Paterson, C. Studies on the distribution of cholesterol, phospholipid, and protein in the human and bovine lens. Lens Eye Toxic. Res. 1989, 6, 703–724. [Google Scholar] [PubMed]
- Cooper, G.M. The Cell: A Molecular Approach. 2nd edition. Sunderland (MA): Sinauer Associates 2000. Structure of the Plasma Membrane. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK9898/ (accessed on 8 April 2021).
- Broekhuyse, R.; Kuhlmann, E.; Stols, A. Lens membranes II. Isolation and characterization of the main intrinsic polypeptide (MIP) of bovine lens fiber membranes. Exp. Eye Res. 1976, 23, 365–371. [Google Scholar] [CrossRef]
- Broekhuyse, R.; Kuhlmann, E.; Winkens, H. Lens membranes VII. MIP is an immunologically specific component of lens fiber membranes and is identical with 26K band protein. Exp. Eye Res. 1979, 29, 303–313. [Google Scholar] [CrossRef]
- Gonen, T.; Cheng, Y.; Sliz, P.; Hiroaki, Y.; Fujiyoshi, Y.; Harrison, S.C.; Walz, T. Lipid–protein interactions in double-layered two-dimensional AQP0 crystals. Nature 2005, 438, 633–638. [Google Scholar] [CrossRef]
- Kar, R.; Batra, N.; Riquelme, M.A.; Jiang, J.X. Biological role of connexin intercellular channels and hemichannels. Arch. Biochem. Biophys. 2012, 524, 2–15. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, J. Alpha-crystallin. Exp. Eye Res. 2003, 76, 145–153. [Google Scholar] [CrossRef]
- Horwitz, J.; Huang, Q.-L.; Ding, L.; Bova, M.P. Lens α-crystallin: Chaperone-like properties. Biomembr. Part K 1998, 290, 365–383. [Google Scholar] [CrossRef]
- Horwitz, J.; Bova, M.P.; Ding, L.-L.; Haley, D.; Stewart, P.L. Lens α-crystallin: Function and structure. Eye 1999, 13, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Lynnerup, N.; Kjeldsen, H.; Heegaard, S.; Jacobsen, C.; Heinemeier, J. Radiocarbon Dating of the Human Eye Lens Crystallines Reveal Proteins without Carbon Turnover throughout Life. PLoS ONE 2008, 3, e1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloemendal, H.; de Jong, W.; Jaenicke, R.; Lubsen, N.H.; Slingsby, C.; Tardieu, A. Ageing and vision: Structure, stability and function of lens crystallins. Prog. Biophys. Mol. Biol. 2004, 86, 407–485. [Google Scholar] [CrossRef] [PubMed]
- De Jong, W. Molecular and Cellular Biology of the Eye Lens. Available online: https://www.abebooks.com/9780471051718/Molecular-Cellular-Biology-Eye-Lens-0471051713/plp (accessed on 1 March 2021).
- Srinivas, P.; Narahari, A.; Petrash, J.M.; Swamy, M.J.; Reddy, G.B. Importance of eye lens α-crystallin heteropolymer with 3:1 αA to αB ratio: Stability, aggregation, and modifications. IUBMB Life 2010, 62, 693–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oertel, M.F.; May, C.A.; Bloemendal, H.; Lütjen-Drecoll, E. Alpha-B-Crystallin Expression in Tissues Derived from Different Species in Different Age Groups. Ophthalmologica 2000, 214, 13–23. [Google Scholar] [CrossRef]
- Moreau, K.L.; King, J.A. Protein misfolding and aggregation in cataract disease and prospects for prevention. Trends Mol. Med. 2012, 18, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Ryazantsev, S.N.; Poliansky, N.B.; Chebotareva, N.A.; Muranov, K.O. 3D structure of the native α-crystallin from bovine eye lens. Int. J. Biol. Macromol. 2018, 117, 1289–1298. [Google Scholar] [CrossRef]
- Kaiser, C.J.O.; Peters, C.; Schmid, P.W.N.; Stavropoulou, M.; Zou, J.; Dahiya, V.; Mymrikov, E.V.; Rockel, B.; Asami, S.; Haslbeck, M.; et al. The structure and oxidation of the eye lens chaperone αA-crystallin. Nat. Struct. Mol. Biol. 2019, 26, 1141–1150. [Google Scholar] [CrossRef]
- Jehle, S.; Vollmar, B.S.; Bardiaux, B.; Dove, K.K.; Rajagopal, P.; Gonen, T.; Oschkinat, H.; Klevit, R.E. N-terminal domain of B-crystallin provides a conformational switch for multimerization and structural heterogeneity. Proc. Natl. Acad. Sci. USA 2011, 108, 6409–6414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagerholm, P.P.; Philipson, B.T.; Lindström, B. Normal human lens—The distribution of protein. Exp. Eye Res. 1981, 33, 615–620. [Google Scholar] [CrossRef]
- Horwitz, J. Alpha-crystallin can function as a molecular chaperone. Proc. Natl. Acad. Sci. USA 1992, 89, 10449–10453. [Google Scholar] [CrossRef] [Green Version]
- Jakob, U.; Gaestel, M.; Engel, K.; Buchner, J. Small heat shock proteins are molecular chaperones. J. Biol. Chem. 1993, 268, 1517–1520. [Google Scholar] [CrossRef]
- Augusteyn, R.C. α-crystallin: A review of its structure and function. Clin. Exp. Optom. 2004, 87, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.A.; Sprague-Piercy, M.A.; Kwok, A.O.; Roskamp, K.W.; Martin, R.W. Chemical Properties Determine Solubility and Stability in βγ-Crystallins of the Eye Lens. ChemBioChem 2021, 22, 1329–1346. [Google Scholar] [CrossRef] [PubMed]
- Maulucci, G.; Papi, M.; Arcovito, G.; De Spirito, M. The Thermal Structural Transition of α-Crystallin Inhibits the Heat Induced Self-Aggregation. PLoS ONE 2011, 6, e18906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehnal, D.; Bittrich, S.; Deshpande, M.; Svobodová, R.; Berka, K.; Bazgier, V.; Velankar, S.; Burley, S.K.; Koča, J.; Rose, A.S. Mol Viewer: Modern web app for 3D visualization and analysis of large biomolecular structures. Nucleic Acids Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Bron, A.; Vrensen, G.; Koretz, J.; Maraini, G.; Harding, J. The Ageing Lens. Ophthalmologica 2000, 214, 86–104. [Google Scholar] [CrossRef]
- Truscott, R.J. Age-related nuclear cataract—Oxidation is the key. Exp. Eye Res. 2005, 80, 709–725. [Google Scholar] [CrossRef]
- Khatry, S.K.; Lewis, A.E.; Schein, O.D.; Thapa, M.D.; Pradhan, E.K.; Katz, J. The epidemiology of ocular trauma in rural Nepal. Br. J. Ophthalmol. 2004, 88, 456–460. [Google Scholar] [CrossRef]
- Négrel, A.-D.; Thylefors, B. The global impact of eye injuries. Ophthalmic Epidemiol. 1998, 5, 143–169. [Google Scholar] [CrossRef]
- Hejtmancik, J.; Kantorow, M. Molecular genetics of age-related cataract. Exp. Eye Res. 2004, 79, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Shiels, A. Genetic Origins of Cataract. Arch. Ophthalmol. 2007, 125, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Jacob, S.; Michel, M.; Brézin, A.P.; Laurier, D.; Bernier, M.O. Ionizing Radiation as a Risk Factor for Cataract: What about Low-Dose Effects? J. Clin. Exp. Ophthalmol. 2013, S1, 005. [Google Scholar] [CrossRef] [Green Version]
- Lipman, R.M.; Tripathi, B.J.; Tripathi, R.C. Cataracts induced by microwave and ionizing radiation. Surv. Ophthalmol. 1988, 33, 200–210. [Google Scholar] [CrossRef]
- Uwineza, A.; Kalligeraki, A.A.; Hamada, N.; Jarrin, M.; Quinlan, R.A. Cataractogenic load—A concept to study the contribution of ionizing radiation to accelerated aging in the eye lens. Mutat. Res. Mutat. Res. 2019, 779, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Hoffer, K.J. Axial Dimension of the Human Cataractous Lens. Arch. Ophthalmol. 1993, 111, 914–918. [Google Scholar] [CrossRef] [PubMed]
- Ignjatović, Z. Secondary cataracts in extreme myopia. Srp. Arh. Celok. Lek. 1998, 126, 239–241. [Google Scholar] [PubMed]
- Metge, P.; Donnadieu, M. Myopia and cataract. Rev. Prat. 1993, 43, 1784–1786. [Google Scholar]
- Solberg, Y.; Rosner, M.; Belkin, M. The Association Between Cigarette Smoking and Ocular Diseases. Surv. Ophthalmol. 1998, 42, 535–547. [Google Scholar] [CrossRef]
- Erie, J.C.; Pueringer, M.R.; Brue, S.M.; Chamberlain, A.M.; Hodge, D.O. Statin Use and Incident Cataract Surgery: A Case-Control Study. Ophthalmic Epidemiol. 2016, 23, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.-L.; Shau, W.-Y.; Chang, C.-H.; Chen, M.-F.; Lai, M.-S. Statin Use and Cataract Surgery: A Nationwide Retrospective Cohort Study in Elderly Ethnic Chinese Patients. Drug Saf. 2013, 36, 1017–1024. [Google Scholar] [CrossRef]
- Wang, J.J.; Rochtchina, E.; Tan, A.G.; Cumming, R.; Leeder, S.R.; Mitchell, P. Use of Inhaled and Oral Corticosteroids and the Long-term Risk of Cataract. Ophthalmology 2009, 116, 652–657. [Google Scholar] [CrossRef]
- Chylack, L.T. Cataracts and Inhaled Corticosteroids. N. Engl. J. Med. 1997, 337, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Cumming, R.G.; Mitchell, P. Inhaled Corticosteroids and Cataract. Drug Saf. 1999, 20, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Hippisley-Cox, J.; Coupland, C. Unintended effects of statins in men and women in England and Wales: Population based cohort study using the QResearch database. BMJ 2010, 340, c2197. [Google Scholar] [CrossRef] [Green Version]
- Leuschen, J.; Mortensen, E.M.; Frei, C.R.; Mansi, E.A.; Panday, V.; Mansi, I. Association of Statin Use with Cataracts. JAMA Ophthalmol. 2013, 131, 1427–1434. [Google Scholar] [CrossRef] [Green Version]
- Wise, S.J.; Nathoo, N.A.; Etminan, M.; Mikelberg, F.S.; Mancini, G.J. Statin Use and Risk for Cataract: A Nested Case-Control Study of 2 Populations in Canada and the United States. Can. J. Cardiol. 2014, 30, 1613–1619. [Google Scholar] [CrossRef]
- Gong, Y.; Feng, K.; Yan, N.; Xu, Y.; Pan, C.-W. Different Amounts of Alcohol Consumption and Cataract. Optom. Vis. Sci. 2015, 92, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, Y.; Li, G. Alcohol and eye diseases: A review of epidemiologic studies. J. Stud. Alcohol 2001, 62, 397–402. [Google Scholar] [CrossRef]
- Kanthan, G.L.; Mitchell, P.; Burlutsky, G.; Wang, J.J. Alcohol Consumption and the Long-Term Incidence of Cataract and Cataract Surgery: The Blue Mountains Eye Study. Am. J. Ophthalmol. 2010, 150, 434–440. [Google Scholar] [CrossRef]
- Lindblad, B.E.; Håkansson, N.; Philipson, B.; Wolk, A. Alcohol Consumption and Risk of Cataract Extraction. Ophthalmology 2007, 114, 680–685. [Google Scholar] [CrossRef]
- Kuang, T.-M.; Tsai, S.-Y.; Hsu, W.-M.; Cheng, C.-Y.; Liu, J.-H.; Chou, P. Body Mass Index and Age-Related Cataract. Arch. Ophthalmol. 2005, 123, 1109–1114. [Google Scholar] [CrossRef] [Green Version]
- Weintraub, J.; Willett, W.; Rosner, B.; Colditz, G.; Seddon, J.; Hankinson, S. A prospective study of the relationship between body mass index and cataract extraction among US women and men. Int. J. Obes. 2002, 26, 1588–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leske, M.; Wu, S.-Y.; Hennis, A.; Connell, A.M.; Hyman, L.; Schachat, A. Diabetes, hypertension, and central obesity as cataract risk factors in a black population: The Barbados Eye Study. Ophthalmology 1999, 106, 35–41. [Google Scholar] [CrossRef]
- Klein, R.; Klein, B.E. Diabetic eye disease. Lancet 1997, 350, 197–204. [Google Scholar] [CrossRef]
- Pollreisz, A.; Schmidt-Erfurth, U. Diabetic Cataract—Pathogenesis, Epidemiology and Treatment. Available online: https://www.hindawi.com/journals/joph/2010/608751/ (accessed on 26 May 2020).
- Boyle, D.L.; Takemoto, L. EM immunolocalization of α-crystallins: Association with the plasma membrane from normal and cataractous human lenses. Curr. Eye Res. 1996, 15, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Cenedella, R.J.; Fleschner, C.R. Selective association of crystallins with lens ‘native’ membrane during dynamic cataractogenesis. Curr. Eye Res. 1992, 11, 801–815. [Google Scholar] [CrossRef]
- Chandrasekher, G.; Cenedella, R.J. Protein associated with human lens ‘native’ membrane during aging and cataract formation. Exp. Eye Res. 1995, 60, 707–717. [Google Scholar] [CrossRef]
- Datiles, M.B.; Ansari, R.R.; Yoshida, J.; Brown, H.; Zambrano, A.I.; Tian, J.; Vitale, S.; Zigler, J.S.; Ferris, F.L.; West, S.K.; et al. Longitudinal Study of Age-Related Cataract Using Dynamic Light Scattering. Ophthalmology 2016, 123, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, M.G.; Truscott, R.J.W. Membrane Association of Proteins in the Aging Human Lens: Profound Changes Take Place in the Fifth Decade of Life. Investig. Opthalmol. Vis. Sci. 2009, 50, 4786–4793. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, M.G.; Truscott, R.J.W. Large-Scale Binding of α-Crystallin to Cell Membranes of Aged Normal Human Lenses: A Phenomenon That Can Be Induced by Mild Thermal Stress. Investig. Opthalmol. Vis. Sci. 2010, 51, 5145–5152. [Google Scholar] [CrossRef] [Green Version]
- Su, S.-P.; McArthur, J.D.; Friedrich, M.G.; Truscott, R.J.; Aquilina, J.A. Understanding the α-crystallin cell membrane conjunction. Mol. Vis. 2011, 17, 2798–2807. [Google Scholar] [PubMed]
- Cobb, B.A.; Petrash, J.M. α-Crystallin Chaperone-like Activity and Membrane Binding in Age-Related Cataracts. Biochemistry 2002, 41, 483–490. [Google Scholar] [CrossRef]
- Truscott, R.J. Presbyopia. Emerging from a blur towards an understanding of the molecular basis for this most common eye condition. Exp. Eye Res. 2009, 88, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Heys, K.R.; Friedrich, M.G.; Truscott, R.J.W. Presbyopia and heat: Changes associated with aging of the human lens suggest a functional role for the small heat shock protein, α-crystallin, in maintaining lens flexibility. Aging Cell 2007, 6, 807–815. [Google Scholar] [CrossRef]
- Schmid, P.W.N.; Lim, N.C.H.; Peters, C.; Back, K.C.; Bourgeois, B.; Pirolt, F.; Richter, B.; Peschek, J.; Puk, O.; Amarie, O.V.; et al. Imbalances in the eye lens proteome are linked to cataract formation. Nat. Struct. Mol. Biol. 2021, 28, 143–151. [Google Scholar] [CrossRef]
- Bessems, G.J.; De Man, B.M.; Bours, J.; Hoenders, H.J. Age-related variations in the distribution of crystallins within the bovine lens. Exp. Eye Res. 1986, 43, 1019–1030. [Google Scholar] [CrossRef]
- Babizhayev, M.A.; Bours, J.; Utikal, K.J. Isoelectric Focusing of Crystallins in Microsections of Calf and Adult Bovine Lens. Ophthalmic Res. 1996, 28, 365–374. [Google Scholar] [CrossRef]
- Bessems, G.J.; Hoenders, H.J.; Wollensak, J. Variation in proportion and molecular weight of native crystallins from single human lenses upon aging and formation of nuclear cataract. Exp. Eye Res. 1983, 37, 627–637. [Google Scholar] [CrossRef]
- Bindels, J.G.; Bours, J.; Hoenders, H.J. Age-dependent variations in the distribution of rat lens water-soluble crystallins, size fractionation and molecular weight determination. Mech. Ageing Dev. 1983, 21, 1–13. [Google Scholar] [CrossRef]
- Bours, J.; Födisch, H.; Hockwin, O. Age-Related Changes in Water and Crystallin Content of the Fetal and Adult Human Lens, Demonstrated by a Microsectioning Technique. Ophthalmic Res. 1987, 19, 235–239. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, X.-J.; Zhu, J.; Xiangjun, C.; Yang, X.; Hu, L.-D.; Ouyang, H.; Patel, S.H.; Jin, X.; Lin, D.; et al. Lanosterol reverses protein aggregation in cataracts. Nature 2015, 523, 607–611. [Google Scholar] [CrossRef]
- Bova, M.P.; Mchaourab, H.S.; Han, Y.; Fung, B.K.-K. Subunit Exchange of Small Heat Shock Proteins. J. Biol. Chem. 2000, 275, 1035–1042. [Google Scholar] [CrossRef] [Green Version]
- Bova, M.P.; Ding, L.-L.; Horwitz, J.; Fung, B.K.-K. Subunit Exchange of αA-Crystallin. J. Biol. Chem. 1997, 272, 29511–29517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oetelaar, P.J.M.V.D.; Van Someren, P.F.H.M.; Thomson, J.A.; Siezen, R.J.; Hoenders, H.J. A dynamic quaternary structure of bovine α-crystallin as indicated from intermolecular exchange of subunits. Biochemistry 1990, 29, 3488–3493. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Borchman, D.; Yappert, M.C. Alpha-Crystallin/Lens Lipid Interactions Using Resonance Energy Transfer. Ophthalmic Res. 1999, 31, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Makley, L.N.; McMenimen, K.A.; DeVree, B.; Goldman, J.; McGlasson, B.N.; Rajagopal, P.; Dunyak, B.M.; McQuade, T.J.; Thompson, A.; Sunahara, R.K.; et al. Pharmacological chaperone for -crystallin partially restores transparency in cataract models. Science 2015, 350, 674–677. [Google Scholar] [CrossRef] [Green Version]
- Deeley, J.M.; Mitchell, T.W.; Wei, X.; Korth, J.; Nealon, J.R.; Blanksby, S.J.; Truscott, R.J. Human lens lipids differ markedly from those of commonly used experimental animals. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2008, 1781, 288–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yappert, M.; Rujoi, M.; Borchman, D.; Vorobyov, I.; Estrada, R. Glycero- versus sphingo-phospholipids: Correlations with human and non-human mammalian lens growth. Exp. Eye Res. 2003, 76, 725–734. [Google Scholar] [CrossRef]
- Li, L.K.; So, L.; Spector, A. Membrane cholesterol and phospholipid in consecutive concentric sections of human lenses. J. Lipid Res. 1985, 26, 600–609. [Google Scholar] [CrossRef]
- Li, L.-K.; So, L.; Spector, A. Age-dependent changes in the distribution and concentration of human lens cholesterol and phospholipids. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1987, 917, 112–120. [Google Scholar] [CrossRef]
- Zelenka, P.S. Lens lipids. Curr. Eye Res. 1984, 3, 1337–1359. [Google Scholar] [CrossRef]
- Iwata, J.L.; Bardygula-Nonn, L.G.; Glonek, T.; Greiner, J.V. Interspecies comparisons of lens phospholipids. Curr. Eye Res. 1995, 14, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Broekhuyse, R. Lipids in tissues of the eye. IV. Influence of age and -species differences on the phospholipid composition of the lens. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1970, 218, 546–548. [Google Scholar] [CrossRef]
- Feldman, G.L.; Culp, T.W.; Feldman, L.S.; Grantham, C.K.; Jonsson, H.T. Phospholipids of the Bovine, Rabbit, and Human Lens. Investig. Ophthalmol. 1964, 3, 194–197. [Google Scholar]
- Borchman, D.; Tang, D. Binding Capacity of α-Crystallin to Bovine Lens Lipids. Exp. Eye Res. 1996, 63, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekher, G.; Cenedella, R.J. Properties of α-crystallin Bound to Lens Membrane: Probing Organization at the Membrane Surface. Exp. Eye Res. 1997, 64, 423–430. [Google Scholar] [CrossRef]
- Ifeanyi, F.; Takemoto, L. Interaction of lens crystallins with lipid vesicles. Exp. Eye Res. 1991, 52, 535–538. [Google Scholar] [CrossRef]
- Cobb, B.A.; Petrash, J.M. Characterization of α-Crystallin-Plasma Membrane Binding. J. Biol. Chem. 2000, 275, 6664–6672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ifeanyi, F.; Takemoto, L. Differential binding of α-crystallins to bovine lens membrane. Exp. Eye Res. 1989, 49, 143–147. [Google Scholar] [CrossRef]
- Ifeanyi, F.; Takemoto, L. Specificity of alpha crystallin binding to the lens membrane. Curr. Eye Res. 1990, 9, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Ifeanyi, F.; Takemoto, L. Alpha crystallin from human cataractous vs. normal lenses: Change in binding to lens membrane. Exp. Eye Res. 1990, 50, 113–116. [Google Scholar] [CrossRef]
- Mulders, J.W.M.; Stokkermans, J.P.W.G.; Leunissen, J.A.M.; Benedetti, E.L.; Bloemendal, H.; Jong, W.W. Interaction of Alpha-crystallin with lens plasma membranes. Affinity for MP26. JBIC J. Biol. Inorg. Chem. 1985, 152, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Z.; Augusteyn, R.C. On the interaction of α-crystallin with membranes. Curr. Eye Res. 1994, 13, 225–230. [Google Scholar] [CrossRef]
- Cobb, B.A.; Petrash, J.M. Factors Influencing α-Crystallin Association with Phospholipid Vesicles. Mol. Vis. 2002, 8, 85–93. [Google Scholar]
- Mainali, L.; O’Brien, W.J.; Timsina, R. Interaction of Alpha-Crystallin with Phospholipid Membranes. Curr. Eye Res. 2021, 46, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Borchman, D. Temperature Induced Structural Changes of β-Crystallin and Sphingomyelin Binding. Exp. Eye Res. 1998, 67, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Timsina, R.; Khadka, N.K.; Maldonado, D.; Mainali, L. Interaction of alpha-crystallin with four major phospholipids of eye lens membranes. Exp. Eye Res. 2021, 202, 108337. [Google Scholar] [CrossRef]
- Tang, D.; Borchman, D.; Yappert, M.C.; Cenedella, R.J. Influence of Cholesterol on the Interaction of α-Crystallin with Phospholipids. Exp. Eye Res. 1998, 66, 559–567. [Google Scholar] [CrossRef]
- Mulders, J.W.; Wojcik, E.; Bloemendal, H.; De Jong, W.W. Loss of high-affinity membrane binding of bovine nuclear α-crystallin. Exp. Eye Res. 1989, 49, 149–152. [Google Scholar] [CrossRef]
- Zhang, Z.Z.; Zeng, J.J.; Yin, H.H.; Tang, D.D.; Borchman, D.D.; Paterson, C.C. Membrane lipid a-crystallin interaction and membrane Ca2+-ATPase activities. Curr. Eye Res. 1999, 18, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Borchman, D.; Ozaki, Y.; Lamba, O.P.; Byrdwell, W.; Yappert, M.; Paterson, C. Lipid-protein Interactions in Human and Bovine Lens Membranes by Fourier Transform Raman and Infrared Spectroscopies. Exp. Eye Res. 1996, 62, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.B.; Kumar, P.A.; Kumar, M.S. Chaperone-like activity and hydrophobicity of α-crystallin. IUBMB Life 2006, 58, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Cenedella, R.J.; Chandrasekher, G. High capacity binding of alpha crystallins to various bovine lens membrane preparations. Curr. Eye Res. 1993, 12, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Timsina, R.; Trossi-Torres, G.; O’Dell, M.; Khadka, N.K.; Mainali, L. Cholesterol and cholesterol bilayer domains inhibit binding of alpha-crystallin to the membranes made of the major phospholipids of eye lens fiber cell plasma membranes. Exp. Eye Res. 2021, 206, 108544. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkova, N.M.; Horvath, I.; Torok, Z.; Wolkers, W.F.; Balogi, Z.; Shigapova, N.; Crowe, L.M.; Tablin, F.; Vierling, E.; Crowe, J.H.; et al. Small heat-shock proteins regulate membrane lipid polymorphism. Proc. Natl. Acad. Sci. USA 2002, 99, 13504–13509. [Google Scholar] [CrossRef] [Green Version]
- Borchman, D. Lipid conformational order and the etiology of cataract and dry eye. J. Lipid Res. 2021, 62, 100039. [Google Scholar] [CrossRef]
- Borchman, D.; Stimmelmayr, R.; George, J.C. Whales, lifespan, phospholipids, and cataracts. J. Lipid Res. 2017, 58, 2289–2298. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Grami, V.; Marrero, Y.; Tang, D.; Yappert, M.C.; Rasi, V.; Borchman, D. Human Lens Phospholipid Changes with Age and Cataract. Investig. Opthalmol. Vis. Sci. 2005, 46, 1682–1689. [Google Scholar] [CrossRef]
- Mainali, L.; Raguz, M.; O’Brien, W.J.; Subczynski, W.K. Changes in the Properties and Organization of Human Lens Lipid Membranes Occurring with Age. Curr. Eye Res. 2017, 42, 721–731. [Google Scholar] [CrossRef]
- Paterson, C.A.; Zeng, J.; Husseini, Z.; Borchman, D.; Delamere, N.A.; Garland, D.; Jimenez-Asensio, J. Calcium ATPase activity and membrane structure in clear and cataractous human lenses. Curr. Eye Res. 1997, 16, 333–338. [Google Scholar] [CrossRef]
- Truscott, R.J. Age-Related Nuclear Cataract: A Lens Transport Problem. Ophthalmic Res. 2000, 32, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Stimmelmayr, R.; Borchman, D. Lens Lipidomes Among Phocidae and Odobenidae. Aquat. Mamm. 2018, 43, 506–518. [Google Scholar] [CrossRef]
- Borchman, D.; Yappert, M.C.; Bin Afzal, M.H. Lens lipids and maximum lifespan. Exp. Eye Res. 2004, 79, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Raguz, M.; Widomska, J.; Dillon, J.; Gaillard, E.R.; Subczynski, W.K. Physical properties of the lipid bilayer membrane made of cortical and nuclear bovine lens lipids: EPR spin-labeling studies. Biochim. Biophys. Acta Biomembr. 2009, 1788, 2380–2388. [Google Scholar] [CrossRef] [Green Version]
- Schreier, S.; Polnaszek, C.F.; Smith, I.C. Spin labels in membranes problems in practice. Biochim. Biophys. Acta Rev. Biomembr. 1978, 515, 395–436. [Google Scholar] [CrossRef]
- Kusumi, A.; Subczynski, W.K.; Pasenkiewicz-Gierula, M.; Hyde, J.S.; Merkle, H. Spin-label studies on phosphatidylcholine-cholesterol membranes: Effects of alkyl chain length and unsaturation in the fluid phase. Biochim. Biophys. Acta Biomembr. 1986, 854, 307–317. [Google Scholar] [CrossRef]
- Mainali, L.; Raguz, M.; Subczynski, W.K. Phases and domains in sphingomyelin–cholesterol membranes: Structure and properties using EPR spin-labeling methods. Eur. Biophys. J. 2011, 41, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Raguz, M.; Mainali, L.; Widomska, J.; Subczynski, W.K. Using spin-label electron paramagnetic resonance (EPR) to discriminate and characterize the cholesterol bilayer domain. Chem. Phys. Lipids 2011, 164, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Borchman, D.; Byrdwell, W.C.; Yappert, M.C. Regional and age-dependent differences in the phospholipid composition of human lens membranes. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3938–3942. [Google Scholar]
- Borchman, D.; Tang, D.; Yappert, M.C. Lipid composition, membrane structure relationships in lens and muscle sarcoplasmic reticulum membranes. Biospectroscopy 1999, 5, 151–167. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Mainali, L.; Raguz, M.; O’Brien, W.J. Organization of lipids in fiber-cell plasma membranes of the eye lens. Exp. Eye Res. 2017, 156, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widomska, J.; Subczynski, W.K. Why Is Very High Cholesterol Content Beneficial for the Eye Lens but Negative for Other Organs? Nutrients 2019, 11, 1083. [Google Scholar] [CrossRef] [Green Version]
- Mainali, L.; Raguz, M.; O’Brien, W.J.; Subczynski, W.K. Properties of membranes derived from the total lipids extracted from clear and cataractous lenses of 61–70-year-old human donors. Eur. Biophys. J. 2014, 44, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Mainali, L.; Raguz, M.; O’Brien, W.J.; Subczynski, W.K. Properties of membranes derived from the total lipids extracted from the human lens cortex and nucleus. Biochim. Biophys. Acta Biomembr. 2013, 1828, 1432–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mainali, L.; Raguz, M.; O’Brien, W.J.; Subczynski, W.K. Properties of fiber cell plasma membranes isolated from the cortex and nucleus of the porcine eye lens. Exp. Eye Res. 2012, 97, 117–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mainali, L.; Pasenkiewicz-Gierula, M.; Subczynski, W.K. Formation of cholesterol Bilayer Domains Precedes Formation of Cholesterol Crystals in Membranes Made of the Major Phospholipids of Human Eye Lens Fiber Cell Plasma Membranes. Curr. Eye Res. 2020, 45, 162–172. [Google Scholar] [CrossRef]
- Mainali, L.; Raguz, M.; Subczynski, W.K. Phase-Separation and Domain-Formation in Cholesterol-Sphingomyelin Mixture: Pulse-EPR Oxygen Probing. Biophys. J. 2011, 101, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Raguz, M.; Mainali, L.; Widomska, J.; Subczynski, W.K. The immiscible cholesterol bilayer domain exists as an integral part of phospholipid bilayer membranes. Biochim. Biophys. Acta Biomembr. 2011, 1808, 1072–1080. [Google Scholar] [CrossRef] [Green Version]
- Raguz, M.; Kumar, S.N.; Zareba, M.; Ilic, N.; Mainali, L.; Subczynski, W.K. Confocal Microscopy Confirmed that in Phosphatidylcholine Giant Unilamellar Vesicles with very High Cholesterol Content Pure Cholesterol Bilayer Domains Form. Cell Biophys. 2019, 77, 309–317. [Google Scholar] [CrossRef]
- Khadka, N.K.; Timsina, R.; Rowe, E.; O’Dell, M.; Mainali, L. Mechanical properties of the high cholesterol-containing membrane: An AFM study. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183625. [Google Scholar] [CrossRef]
- Mainali, L.; O’Brien, W.J.; Subczynski, W.K. Detection of cholesterol bilayer domains in intact biological membranes: Methodology development and its application to studies of eye lens fiber cell plasma membranes. Exp. Eye Res. 2019, 178, 72–81. [Google Scholar] [CrossRef]
- Widomska, J.; Subczynski, W.K.; Mainali, L.; Raguz, M. Cholesterol Bilayer Domains in the Eye Lens Health: A Review. Cell Biophys. 2017, 75, 387–398. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Raguz, M.; Widomska, J.; Mainali, L.; Konovalov, A. Functions of Cholesterol and the Cholesterol Bilayer Domain Specific to the Fiber-Cell Plasma Membrane of the Eye Lens. J. Membr. Biol. 2012, 245, 51–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada, R.; Puppato, A.; Borchman, D.; Yappert, M.C. Reevaluation of the phospholipid composition in membranes of adult human lenses by 31P NMR and MALDI MS. Biochim. Biophys. Acta Biomembr. 2010, 1798, 303–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subczynski, W.K.; Wisniewska, A.; Yin, J.-J.; Hyde, J.S.; Kusumi, A. Hydrophobic Barriers of Lipid Bilayer Membranes Formed by Reduction of Water Penetration by Alkyl Chain Unsaturation and Cholesterol. Biochemistry 1994, 33, 7670–7681. [Google Scholar] [CrossRef] [PubMed]
- Mainali, L.; Feix, J.B.; Hyde, J.S.; Subczynski, W.K. Membrane fluidity profiles as deduced by saturation-recovery EPR measurements of spin-lattice relaxation times of spin labels. J. Magn. Reson. 2011, 212, 418–425. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Buboltz, J.T.; Feigenson, G.W. Maximum solubility of cholesterol in phosphatidylcholine and phosphatidylethanolamine bilayers. Biochim. Biophys. Acta Biomembr. 1999, 1417, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Jacob, R.F.; Cenedella, R.J.; Mason, R.P. Evidence for Distinct Cholesterol Domains in Fiber Cell Membranes from Cataractous Human Lenses. J. Biol. Chem. 2001, 276, 13573–13578. [Google Scholar] [CrossRef] [Green Version]
- Bhuyan, K.C.; Bhuyan, D.K.; Podos, S.M. Lipid peroxidation in cataract of the human. Life Sci. 1986, 38, 1463–1471. [Google Scholar] [CrossRef]
- Babizhayev, M.A. Lipid fluorophores of the human crystalline lens with cataract. Graefe Arch. Clin. Exp. Ophthalmol. 1989, 227, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Borchman, D.; Yappert, M.C.; Rubini, R.Q.; Paterson, C. Distribution of phospholipid-malondialdehyde-adduct in the human lens. Curr. Eye Res. 1989, 8, 939–946. [Google Scholar] [PubMed]
- Simonelli, F.; Nesti, A.; Pensa, M.; Romano, L.; Savastano, S.; Rinaldi, E.; Auricchio, G. Lipid peroxidation and human cataractogenesis in diabetes and severe myopia. Exp. Eye Res. 1989, 49, 181–187. [Google Scholar] [CrossRef]
- Borchman, D.; Yappert, M.C. Age-related lipid oxidation in human lenses. Investig. Ophthalmol. Vis. Sci. 1998, 39, 1053–1058. [Google Scholar]
- Babizhayev, M.A.; Deyev, A.I.; Linberg, L.F. Lipid peroxidation as a possible cause of cataract. Mech. Ageing Dev. 1988, 44, 69–89. [Google Scholar] [CrossRef]
- Bhuyan, K.C.; Master, R.W.; Coles, R.S.; Bhuyan, D.K. Molecular mechanisms of cataractogenesis: IV. Evidence of phospholipid · malondialdehyde adduct in human senile cataract. Mech. Ageing Dev. 1986, 34, 289–296. [Google Scholar] [CrossRef]
- Bhuyan, K.C.; Bhuyan, D.K. Molecular mechanism of cataractogenesis: III. Toxic metabolites of oxygen as initiators of lipid peroxidation and cataract. Curr. Eye Res. 1984, 3, 67–82. [Google Scholar] [CrossRef]
- Micelli-Ferrari, T.; Vendemiale, G.; Grattagliano, I.; Boscia, F.; Arnese, L.; Altomare, E.; Cardia, L. Role of lipid peroxidation in the pathogenesis of myopic and senile cataract. Br. J. Ophthalmol. 1996, 80, 840–843. [Google Scholar] [CrossRef] [Green Version]
- Varma, S.D.; Chand, D.; Sharma, Y.R.; Kuck, J.F.; Richards, R.D. Oxidative stress on lens and cataract formation: Role of light and oxygen. Curr. Eye Res. 1984, 3, 35–58. [Google Scholar] [CrossRef]
- Babizhayev, M.A. Accumulation of lipid peroxidation products in human cataracts. Acta Ophthalmol. 2009, 67, 281–287. [Google Scholar] [CrossRef]
- Hughes, J.R.; Levchenko, V.; Blanksby, S.J.; Mitchell, T.W.; Williams, A.; Truscott, R.J.W. No turnover in lens lipids for the entire human lifespan. eLife 2015, 4, e06003. [Google Scholar] [CrossRef] [PubMed]
- Witting, L.A. Lipid peroxidationin vivo. J. Am. Oil Chem. Soc. 1965, 42, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Byrdwell, C.; Borchman, D. Liquid Chromatography/Mass-Spectrometric Characterization of Sphingomyelin and Dihydrosphingomyelin of Human Lens Membranes. Ophthalmic Res. 1997, 29, 191–206. [Google Scholar] [CrossRef]
- Deeley, J.M.; Hankin, J.A.; Friedrich, M.G.; Murphy, R.C.; Truscott, R.J.; Mitchell, T.W.; Blanksby, S.J. Sphingolipid distribution changes with age in the human lens. J. Lipid Res. 2010, 51, 2753–2760. [Google Scholar] [CrossRef] [Green Version]
- Oborina, E.M.; Yappert, M. Effect of sphingomyelin versus dipalmitoylphosphatidylcholine on the extent of lipid oxidation. Chem. Phys. Lipids 2003, 123, 223–232. [Google Scholar] [CrossRef]
- Huang, L.; Tang, D.; Yappert, M.C.; Borchman, D. Oxidation-induced changes in human lens epithelial cells 2. Mitochondria and the generation of reactive oxygen species. Free Radic. Biol. Med. 2006, 41, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Yappert, M.C.; Miller, J.J.; Borchman, D. Thyroxine Ameliorates Oxidative Stress by Inducing Lipid Compositional Changes in Human Lens Epithelial Cells. Investig. Opthalmol. Vis. Sci. 2007, 48, 3698–3704. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Estrada, R.; Yappert, M.C.; Borchman, D. Oxidation-induced changes in human lens epithelial cells: 1. Phospholipids. Free Radic. Biol. Med. 2006, 41, 1425–1432. [Google Scholar] [CrossRef]
- Huang, L.; Yappert, M.C.; Jumblatt, M.M.; Borchman, D. Hyperoxia and Thyroxine Treatment and the Relationships between Reactive Oxygen Species Generation, Mitochondrial Membrane Potential, and Cardiolipin in Human Lens Epithelial Cell Cultures. Curr. Eye Res. 2008, 33, 575–586. [Google Scholar] [CrossRef]
- Jacob, R.F.; Mason, R.P. Lipid Peroxidation Induces Cholesterol Domain Formation in Model Membranes. J. Biol. Chem. 2005, 280, 39380–39387. [Google Scholar] [CrossRef] [Green Version]
- Jacob, R.F.; Aleo, M.D.; Self-Medlin, Y.; Doshna, C.M.; Mason, R.P. 1,2-Naphthoquinone Stimulates Lipid Peroxidation and Cholesterol Domain Formation in Model Membranes. Investig. Opthalmol. Vis. Sci. 2013, 54, 7189–7197. [Google Scholar] [CrossRef] [Green Version]
- Mainali, L.; Zareba, M.; Subczynski, W.K. Oxidation of Polyunsaturated Phospholipid Decreases the Cholesterol Content at which Cholesterol Bilayer Domains Start to form in Phospholipid-Cholesterol Membranes. Biophys. J. 2017, 112, 375a. [Google Scholar] [CrossRef] [Green Version]
- Choe, M.; Jackson, C.; Yu, B.P. Lipid peroxidation contributes to age-related membrane rigidity. Free Radic. Biol. Med. 1995, 18, 977–984. [Google Scholar] [CrossRef]
- Borchman, D.; Lamba, O.P.; Salmassi, S.; Lou, M.; Yappert, M.C. The dual effect of oxidation on lipid bilayer structure. Lipids 1992, 27, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Galeotti, T.; Borrello, S.; Palombini, G.; Masotti, L.; Ferrari, M.; Cavatorta, P.; Arcioni, A.; Stremmenos, C.; Zannoni, C. Lipid peroxidation and fluidity of plasma membranes from rat liver and Morris hepatoma 3924A. FEBS Lett. 1984, 169, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Dobretsov, G.; Borschevskaya, T.; Petrov, V.; Vladimirov, Y. The increase of phospholipid bilayer rigidity after lipid peroxidation. FEBS Lett. 1977, 84, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Delmelle, M. Retinal Sensitized Photodynamic Damage to Liposomes. Photochem. Photobiol. 1978, 28, 357–360. [Google Scholar] [CrossRef]
- Fukuzawa, K.; Chida, H.; Tokumura, A.; Tsukatani, H. Antioxidative effect of α-tocopherol incorporation into lecithin liposomes on ascorbic acid-Fe2+-induced lipid peroxidation. Arch. Biochem. Biophys. 1981, 206, 173–180. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Hochstein, P. Alterations in erythrocyte membrane fluidity by phenylhydrazine-induced peroxidation of lipids. Biochem. Biophys. Res. Commun. 1981, 100, 1537–1542. [Google Scholar] [CrossRef]
- Eichenberger, K.; Böhni, P.; Winterhalter, K.H.; Kawato, S.; Richter, C. Microsomal lipid peroxidation causes an increase in the order of the membrane lipid domain. FEBS Lett. 1982, 142, 59–62. [Google Scholar] [CrossRef] [Green Version]
- Pauls, K.P.; Thompson, J.E. In vitro simulation of senescence-related membrane damage by ozone-induced lipid peroxidation. Nature 1980, 283, 504–506. [Google Scholar] [CrossRef]
- Coolbear, K.; Keough, K. Lipid oxidation and gel to liquid-crystalline transition temperatures of synthetic polyunsaturated mixed-acid phosphatidylcholines. Biochim. Biophys. Acta Biomembr. 1983, 732, 531–540. [Google Scholar] [CrossRef]
- Braun, A.G.; Buckner, C.A. Evidence of far ultraviolet light-mediated changes in plasma membrane structure and function. Biochim. Biophys. Acta Biomembr. 1981, 648, 263–266. [Google Scholar] [CrossRef]
- Goodenough, D.A. The crystalline lens. A system networked by gap junctional intercellular communication. Semin. Cell Biol. 1992, 3, 49–58. [Google Scholar] [CrossRef]
- Hall, J.E.; Freites, J.A.; Tobias, D.J. Experimental and Simulation Studies of Aquaporin 0 Water Permeability and Regulation. Chem. Rev. 2019, 119, 6015–6039. [Google Scholar] [CrossRef] [PubMed]
- Varadaraj, K.; Kushmerick, C.; Baldo, G.; Bassnett, S.; Shiels, A.; Mathias, R. The role of MIP in lens fiber cell membrane transport. J. Membr. Biol. 1999, 170, 191–203. [Google Scholar] [CrossRef]
- Buzhynskyy, N.; Sens, P.; Behar-Cohen, F.; Scheuring, S. Eye lens membrane junctional microdomains: A comparison between healthy and pathological cases. New J. Phys. 2011, 13, 085016. [Google Scholar] [CrossRef] [Green Version]
- Agre, P. Aquaporin Water Channels. Biosci. Rep. 2004, 24, 127–163. [Google Scholar] [CrossRef] [PubMed]
- Francis, P.; Chung, J.-J.; Yasui, M.; Berry, V.; Moore, A.; Wyatt, M.K.; Wistow, G.; Bhattacharya, S.S.; Agre, P. Functional impairment of lens aquaporin in two families with dominantly inherited cataracts. Hum. Mol. Genet. 2000, 9, 2329–2334. [Google Scholar] [CrossRef] [Green Version]
- Varadaraj, K.; Kumari, S.; Patil, R.; Wax, M.; Mathias, R. Functional characterization of a human aquaporin 0 mutation that leads to a congenital dominant lens cataract. Exp. Eye Res. 2008, 87, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Verkman, A. Role of aquaporin water channels in eye function. Exp. Eye Res. 2003, 76, 137–143. [Google Scholar] [CrossRef]
- Mathias, R.T.; White, T.; Gong, X. Lens Gap Junctions in Growth, Differentiation, and Homeostasis. Physiol. Rev. 2010, 90, 179–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, J.; Berthoud, V.M.; Novak, L.; Zoltoski, R.; Heilbrunn, B.; Minogue, P.J.; Liu, X.; Ebihara, L.; Kuszak, J.; Beyer, E.C. Transgenic overexpression of connexin50 induces cataracts. Exp. Eye Res. 2007, 84, 513–528. [Google Scholar] [CrossRef] [Green Version]
- Tenbroek, E.; Arneson, M.; Jarvis, L.; Louis, C. The distribution of the fiber cell intrinsic membrane proteins MP20 and connexin46 in the bovine lens. J. Cell Sci. 1992, 103, 245–257. [Google Scholar] [CrossRef]
- White, T.; Goodenough, D.A.; Paul, D.L. Targeted Ablation of Connexin50 in Mice Results in Microphthalmia and Zonular Pulverulent Cataracts. J. Cell Biol. 1998, 143, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.-H.; Cheng, C.; Huang, Q.; Cheung, D.; Li, L.; Dunia, I.; Benedetti, L.E.; Horwitz, J.; Gong, X. Absence of α3 (Cx46) and α8 (Cx50) connexins leads to cataracts by affecting lens inner fiber cells. Exp. Eye Res. 2006, 83, 688–696. [Google Scholar] [CrossRef]
- Liang, J.J.; Li, X.-Y. Spectroscopic studies on the interaction of calf lens membranes with crystallins. Exp. Eye Res. 1992, 54, 719–724. [Google Scholar] [CrossRef]
- Liu, B.-F.; Liang, J.J. Confocal fluorescence microscopy study of interaction between lens MIP26/AQP0 and crystallins in living cells. J. Cell. Biochem. 2008, 104, 51–58. [Google Scholar] [CrossRef]
- Gorin, M.B.; Yancey, S.; Cline, J.; Revel, J.-P.; Horwitz, J. The major intrinsic protein (MIP) of the bovine lens fiber membrane: Characterization and structure based on cDNA cloning. Cell 1984, 39, 49–59. [Google Scholar] [CrossRef]
- Bloemendal, H.; Hermsen, T.; Dunia, I.; Benedetti, E.L. Association of crystallins with the plasma membrane. Exp. Eye Res. 1982, 35, 61–67. [Google Scholar] [CrossRef]
- Liang, J.J.; Sun, T.X.; Akhtar, N.J. Heat-Induced Conformational Change of Human Lens Recombinant AlphaA- and AlphaB-Cryst. Mol. Vis. 2000, 6, 10–14. [Google Scholar]
- Ball, L.E.; Garland, D.L.; Crouch, R.K.; Schey, K.L. Post-translational Modifications of Aquaporin 0 (AQP0) in the Normal Human Lens: Spatial and Temporal Occurrence. Biochemistry 2004, 43, 9856–9865. [Google Scholar] [CrossRef]
- Raguz, M.; Mainali, L.; O’Brien, W.J.; Subczynski, W.K. Lipid domains in intact fiber-cell plasma membranes isolated from cortical and nuclear regions of human eye lenses of donors from different age groups. Exp. Eye Res. 2015, 132, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raguz, M.; Mainali, L.; O’Brien, W.J.; Subczynski, W.K. Lipid-protein interactions in plasma membranes of fiber cells isolated from the human eye lens. Exp. Eye Res. 2014, 120, 138–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichow, S.L.; Gonen, T. Lipid–protein interactions probed by electron crystallography. Curr. Opin. Struct. Biol. 2009, 19, 560–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, J.; Canty, J.; Briggs, M.M.; McIntosh, T.J. The water permeability of lens aquaporin-0 depends on its lipid bilayer environment. Exp. Eye Res. 2013, 113, 32–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieri, V.G.; Wallach, D.F.H. Variations of lipid-protein interactions in erythrocyte ghosts as a function of temperature and pH in physiological and non-physiological ranges: A study using paramagnetic quenching of protein fluorescence by nitroxide lipid analogues. Biochim. Biophys. Acta Biomembr. 1975, 406, 415–423. [Google Scholar] [CrossRef]
- Warren, G.B.; Houslay, M.D.; Metcalfe, J.C.; Birdsall, N.J.M. Cholesterol is excluded from the phospholipid annulus surrounding an active calcium transport protein. Nature 1975, 255, 684–687. [Google Scholar] [CrossRef] [PubMed]
- Borchman, D.; Cenedella, R.J.; Lamba, O.P. Role of Cholesterol in the Structural Order of Lens Membrane Lipids. Exp. Eye Res. 1996, 62, 191–198. [Google Scholar] [CrossRef]
- Yappert, M.; Borchman, U. Sphingolipids in human lens membranes: An update on their composition and possible biological implications. Chem. Phys. Lipids 2004, 129, 1–20. [Google Scholar] [CrossRef]
- Zigman, S.; Paxhia, T.; Marinetti, G.; Girsch, S. Lipids of human lens fiber cell membranes. Curr. Eye Res. 1984, 3, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Derham, B.K.; Van Boekel, M.A.M.; Muchowski, P.J.; Clark, J.I.; Horwitz, J.; Hepburne-Scott, H.W.; De Jong, W.W.; Crabbe, M.J.C.; Harding, J.J. Chaperone function of mutant versions of αA- and αB-crystallin prepared to pinpoint chaperone binding sites. JBIC J. Biol. Inorg. Chem. 2001, 268, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Derham, B.K. Alpha-Crystallin as a molecular chaperone. Prog. Retin. Eye Res. 1999, 18, 463–509. [Google Scholar] [CrossRef]
- Andley, U.P. αA-crystallin R49Cneomutation influences the architecture of lens fiber cell membranes and causes posterior and nuclear cataracts in mice. BMC Ophthalmol. 2009, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Cobb, B.A.; Petrash, J.M. Structural and Functional Changes in the αA-Crystallin R116C Mutant in Hereditary Cataracts. Biochemistry 2000, 39, 15791–15798. [Google Scholar] [CrossRef]
- Brown, Z.; Ponce, A.; Lampi, K.; Hancock, L.; Takemoto, L. Differential Binding of Mutant (R116C) and Wildtype AlphaA Crystallin to Actin. Curr. Eye Res. 2007, 32, 1051–1054. [Google Scholar] [CrossRef] [Green Version]
- Grosas, A.B.; Carver, J.A. Eye Lens Crystallins: Remarkable Long-Lived Proteins. In Long-lived Proteins in Human Aging and Disease; WILEY-VCH GmbH: Weinheim, Germany, 2021; pp. 59–96. [Google Scholar]
- Litt, M.; Kramer, P.; LaMorticella, D.M.; Murphey, W.; Lovrien, E.W.; Weleber, R.G. Autosomal dominant congenital cataract associated with a missense mutation in the human alpha crystallin gene CRYAA. Hum. Mol. Genet. 1998, 7, 471–474. [Google Scholar] [CrossRef] [Green Version]
- Gu, F.; Luo, W.; Li, X.; Wang, Z.; Lu, S.; Zhang, M.; Zhao, B.; Zhu, S.; Feng, S.; Yan, Y.-B.; et al. A novel mutation in AlphaA-crystallin (CRYAA) caused autosomal dominant congenital cataract in a large Chinese family. Hum. Mutat. 2008, 29, 769. [Google Scholar] [CrossRef]
- Vicart, P.; Caron, A.; Guicheney, P.; Li, Z.; Prévost, M.-C.; Faure, A.; Chateau, D.; Chapon, F.; Tomé, F.; Dupret, J.-M.; et al. A missense mutation in the αB-crystallin chaperone gene causes a desmin-related myopathy. Nat. Genet. 1998, 20, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Liang, J.J.-N. Alteration of Protein–Protein Interactions of Congenital Cataract Crystallin Mutants. Investig. Opthalmol. Vis. Sci. 2003, 44, 1155–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodehl, A.; Gaertner-Rommel, A.; Klauke, B.; Grewe, S.A.; Schirmer, I.; Peterschröder, A.; Faber, L.; Vorgerd, M.; Gummert, J.; Anselmetti, D.; et al. The novel αB-crystallin (CRYAB) mutation p.D109G causes restrictive cardiomyopathy. Hum. Mutat. 2017, 38, 947–952. [Google Scholar] [CrossRef] [PubMed]
- Frydman, M.; Levy-Nissenbaum, E.; Bakhan, T.; Raz, J.; Assia, E.; Goldman, B.; Pras, E. A nonsense mutation (W9X) in CRYAA causes autosomal recessive cataract in an inbred Jewish Persian family. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3511–3515. [Google Scholar]
- Graw, J.; Löster, J.; Soewarto, D.; Fuchs, H.; Meyer, B.; Reis, A.; Wolf, E.; Balling, R.; Hrabé de Angelis, M. Characterization of a New, Dominant V124E Mutation in the Mouse AlphaA-Crystallin-Encoding Gene. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2909–2915. [Google Scholar]
- Berry, V.; Francis, P.; Reddy, M.A.; Collyer, D.; Vithana, E.; Mackay, I.; Dawson, G.; Carey, A.H.; Moore, A.; Bhattacharya, S.S.; et al. Alpha-B Crystallin Gene (CRYAB) Mutation Causes Dominant Congenital Posterior Polar Cataract in Humans. Am. J. Hum. Genet. 2001, 69, 1141–1145. [Google Scholar] [CrossRef] [Green Version]
- Santhoshkumar, P.; Sharma, K.K. Analysis of alpha-crystallin chaperone function using restriction enzymes and citrate synthase. Mol. Vis. 2001, 7, 172–177. [Google Scholar]
- Santhoshkumar, P.; Sharma, K.K. Phe71 Is Essential for Chaperone-like Function in αA-crystallin. J. Biol. Chem. 2001, 276, 47094–47099. [Google Scholar] [CrossRef] [Green Version]
- Bhagyalaxmi, S.; Srinivas, P.; Barton, K.A.; Kumar, K.R.; Vidyavathi, M.; Petrash, J.M.; Reddy, G.B.; Padma, T. A novel mutation (F71L) in αA-Crystallin with defective chaperone-like function associated with age-related cataract. Biochim. Biophys. Acta Mol. Basis Dis. 2009, 1792, 974–981. [Google Scholar] [CrossRef] [Green Version]
- Kamei, A.; Iwase, H.; Masuda, K. Cleavage of Amino Acid Residue(s) from the N-Terminal Region of αA- and αB-Crystallins in Human Crystalline Lens during Aging. Biochem. Biophys. Res. Commun. 1997, 231, 373–378. [Google Scholar] [CrossRef]
- Kamei, A.; Hamaguchi, T.; Matsuura, N.; Masuda, K. Does Post-translational Modification Influence Chaperone-like AActivity of α-Crystallin? I. Study on Phosphorylation. Biol. Pharm. Bull. 2001, 24, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Blakytny, R.; Carver, J.; Harding, J.J.; Kilby, G.W.; Sheil, M.M. A spectroscopic study of glycated bovine α-crystallin: Investigation of flexibility of the C-terminal extension, chaperone activity and evidence for diglycation. Biochim. Biophys. Acta Protein Struct. Mol. Enzym. 1997, 1343, 299–315. [Google Scholar] [CrossRef]
- Derham, B.K.; Harding, J.J. Effects of modifications of α-crystallin on its chaperone and other properties. Biochem. J. 2002, 364, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Van Boekel, M.A.; Hoogakker, S.E.; Harding, J.J.; De Jong, W.W. The Influence of Some Post-Translational Modifications on the Chaperone-Like Activity of α-Crystallin. Ophthalmic Res. 1996, 28, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Aquilina, J.A.; Benesch, J.L.P.; Ding, L.L.; Yaron, O.; Horwitz, J.; Robinson, C.V. Phosphorylation of αB-Crystallin Alters Chaperone Function through Loss of Dimeric Substructure. J. Biol. Chem. 2004, 279, 28675–28680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truscott, R.J.; Friedrich, M.G. The etiology of human age-related cataract. Proteins don’t last forever. Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 192–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, J.M.; Srivastava, K.; Gupta, R.; Srivastava, O.P. Structural and Functional Roles of Deamidation and/or Truncation of N- or C-Termini in Human αA-Crystallin. Biochemistry 2008, 47, 10069–10083. [Google Scholar] [CrossRef]
- Takemoto, L.J. Quantitation of asparagine-101 deamidation from alpha-A crystallin during aging of the human lens. Curr. Eye Res. 1998, 17, 247–250. [Google Scholar] [CrossRef]
- Lund, A.L.; Smith, J.B.; Smith, D.L. Modifications of the Water-insoluble Human Lens α-Crystallins. Exp. Eye Res. 1996, 63, 661–672. [Google Scholar] [CrossRef]
- Miesbauer, L.; Zhou, X.; Yang, Z.; Sun, Y.; Smith, D.; Smith, J. Post-translational modifications of water-soluble human lens crystallins from young adults. J. Biol. Chem. 1994, 269, 12494–12502. [Google Scholar] [CrossRef]
- Lin, P.P.; Barry, R.C.; Smith, D.L.; Smith, J.B. In vivo acetylation identified at lysine 70 of human lens alphaA-crystallin. Protein Sci. 1998, 7, 1451–1457. [Google Scholar] [CrossRef]
- Takemoto, L.J. Differential Phosphorylation of Alpha-A Crystallin in Human Lens of Different Age. Exp. Eye Res. 1996, 62, 499–504. [Google Scholar] [CrossRef]
- Hanson, S.R.; Hasan, A.; Smith, D.L.; Smith, J.B. The Major in vivo Modifications of the Human Water-insoluble Lens Crystallins are Disulfide Bonds, Deamidation, Methionine Oxidation and Backbone Cleavage. Exp. Eye Res. 2000, 71, 195–207. [Google Scholar] [CrossRef]
- Kim, Y.; Choi, M.; Kim, Y.; Han, J.; Lee, J.; Park, C.; Kang, S.; Choi, W.; Cho, G. Protein kinase C δ regulates anti-apoptotic αB-crystallin in the retina of type 2 diabetes. Neurobiol. Dis. 2007, 28, 293–303. [Google Scholar] [CrossRef]
- Fujii, N.; Takata, T.; Kim, I.; Morishima, K.; Inoue, R.; Magami, K.; Matsubara, T.; Sugiyama, M.; Koide, T. Asp isomerization increases aggregation of α-crystallin and decreases its chaperone activity in human lens of various ages. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140446. [Google Scholar] [CrossRef]
- Lyon, Y.A.; Collier, M.; Riggs, D.L.; Degiacomi, M.T.; Benesch, J.L.P.; Julian, R.R. Structural and functional consequences of age-related isomerization in α-crystallins. J. Biol. Chem. 2019, 294, 7546–7555. [Google Scholar] [CrossRef] [Green Version]
- De Jong, W.W.; Terwindt, E.C.; Bloemendal, H. The amino acid sequence of the A chain of human α-crystallin. FEBS Lett. 1975, 58, 310–313. [Google Scholar] [CrossRef] [Green Version]
- Nagaraj, R.H.; Nahomi, R.B.; Shanthakumar, S.; Linetsky, M.; Padmanabha, S.; Pasupuleti, N.; Wang, B.; Santhoshkumar, P.; Panda, A.K.; Biswas, A. Acetylation of αA-crystallin in the human lens: Effects on structure and chaperone function. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 120–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, R.E.; Swamy, M.; Abraham, E. Progressive changes in lens crystallin glycation and high-molecular-weight aggregate formation leading to cataract development in streptozotocin-diabetic rats. Exp. Eye Res. 1987, 44, 269–282. [Google Scholar] [CrossRef]
- Swamy, M.S.; Abraham, E.C. Lens protein composition, glycation and high molecular weight aggregation in aging rats. Investig. Ophthalmol. Vis. Sci. 1987, 28, 1693–1701. [Google Scholar]
- Swamy, M.; Tsai, C.; Abraham, A.; Abraham, E. Glycation Mediated Lens Crystallin Aggregation and Cross-linking by Various Sugars and Sugar Phosphates In Vitro. Exp. Eye Res. 1993, 56, 177–185. [Google Scholar] [CrossRef]
- Shih, M.; Lampi, K.J.; Shearer, T.R.; David, L.L. Cleavage of beta crystallins during maturation of bovine lens. Mol. Vis. 1998, 4, 4. [Google Scholar]
- Thampi, P.; Hassan, A.; Smith, J.B.; Abraham, E.C. Enhanced C-Terminal Truncation of AlphaA- and AlphaB-Crystallins in Diabetic Lenses. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3265–3272. [Google Scholar]
- Rajan, S.; Horn, C.; Abraham, E.C. Effect of Oxidation of αA- and αB-Crystallins on their Structure, Oligomerization and Chaperone Function. Mol. Cell. Biochem. 2006, 288, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R. Oxidation of Free Amino Acids and Amino Acid Residues in Proteins by Radiolysis and by Metal-Catalyzed Reactions. Annu. Rev. Biochem. 1993, 62, 797–821. [Google Scholar] [CrossRef]
- Cherian-Shaw, M.; Smith, J.B.; Jiang, X.-Y.; Abraham, E.C. Intrapolypeptide disulfides in human αA-crystallin and their effect on chaperone-like function. Mol. Cell. Biochem. 1999, 199, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Lampi, K.J.; Wilmarth, P.A.; Murray, M.R.; David, L.L. Lens β-crystallins: The role of deamidation and related modifications in aging and cataract. Prog. Biophys. Mol. Biol. 2014, 115, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampi, K.J.; Ma, Z.; Hanson, S.R.; Azuma, M.; Shih, M.; Shearer, T.R.; Smith, D.L.; Smith, J.B.; David, L.L. Age-related Changes in Human Lens Crystallins Identified by Two-dimensional Electrophoresis and Mass Spectrometry. Exp. Eye Res. 1998, 67, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Hains, P.G.; Truscott, R.J.W. Age-Dependent Deamidation of Lifelong Proteins in the Human Lens. Investig. Opthalmol. Vis. Sci. 2010, 51, 3107–3114. [Google Scholar] [CrossRef]
- Wilmarth, P.A.; Tanner, S.; Dasari, S.; Nagalla, S.R.; Riviere, M.A.; Bafna, V.; Pevzner, P.A.; David, L.L. Age-Related Changes in Human Crystallins Determined from Comparative Analysis of Post-translational Modifications in Young and Aged Lens: Does Deamidation Contribute to Crystallin Insolubility? J. Proteome Res. 2006, 5, 2554–2566. [Google Scholar] [CrossRef] [Green Version]
- Warmack, R.A.; Shawa, H.; Liu, K.; Lopez, K.; Loo, J.A.; Horwitz, J.; Clarke, S.G. The l-isoaspartate modification within protein fragments in the aging lens can promote protein aggregation. J. Biol. Chem. 2019, 294, 12203–12219. [Google Scholar] [CrossRef]
- Kato, K.; Inaguma, Y.; Ito, H.; Iida, K.; Iwamoto, I.; Kamei, K.; Ochi, N.; Ohta, H.; Kishikawa, M. Ser-59 is the major phosphorylation site in αB-crystallin accumulated in the brains of patients with Alexander’s disease. J. Neurochem. 2008, 76, 730–736. [Google Scholar] [CrossRef]
- Chiappori, F.; Mattiazzi, L.; Milanesi, L.; Merelli, I. A novel molecular dynamics approach to evaluate the effect of phosphorylation on multimeric protein interface: The αB-Crystallin case study. BMC Bioinform. 2016, 17, 57. [Google Scholar] [CrossRef] [Green Version]
- Carver, J.A.; Nicholls, K.A.; Aquilina, A.J.; Truscott, R.J. Age-related Changes in Bovine α-crystallin and High-molecular-weight Protein. Exp. Eye Res. 1996, 63, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, L.J. Identification of the in vivo truncation sites at the C-terminal region of alpha-A crystallin from aged bovine and human lens. Curr. Eye Res. 1995, 14, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Harrington, V.; McCall, S.; Huynh, S.; Srivastava, K.; Srivastava, O.P. Crystallins in water soluble-high molecular weight protein fractions and water insoluble protein fractions in aging and cataractous human lenses. Mol. Vis. 2004, 10, 476–489. [Google Scholar]
- Santhoshkumar, P.; Udupa, P.; Murugesan, R.; Sharma, K.K. Significance of Interactions of Low Molecular Weight Crystallin Fragments in Lens Aging and Cataract Formation. J. Biol. Chem. 2008, 283, 8477–8485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, O.; Srivastava, K.; Chaves, J.; Gill, A. Post-translationally modified human lens crystallin fragments show aggregation in vitro. Biochem. Biophys. Rep. 2017, 10, 94–131. [Google Scholar] [CrossRef]
- Stadtman, E.R. Protein oxidation and aging. Science 1992, 257, 1220–1224. [Google Scholar] [CrossRef] [Green Version]
- Amici, A.; Levine, R.L.; Tsai, L.; Stadtman, E.R. Conversion of Amino Acid Residues in Proteins and Amino Acid Homopolymers to Carbonyl Derivatives by Metal-catalyzed Oxidation Reactions. J. Biol. Chem. 1989, 264, 3341–3346. [Google Scholar] [CrossRef]
- Kim, I.; Saito, T.; Fujii, N.; Kanamoto, T.; Chatake, T. Site specific oxidation of amino acid residues in rat lens γ-crystallin induced by low-dose γ-irradiation. Biochem. Biophys. Res. Commun. 2015, 466, 622–628. [Google Scholar] [CrossRef]
- Cherian, M.; Abraham, E. Decreased Molecular Chaperone Property of α-Crystallins Due to Posttranslational Modifications. Biochem. Biophys. Res. Commun. 1995, 208, 675–679. [Google Scholar] [CrossRef]
- Hains, P.G.; Truscott, R.J. Proteomic analysis of the oxidation of cysteine residues in human age-related nuclear cataract lenses. Biochim. Biophys. Acta Proteins Proteom. 2008, 1784, 1959–1964. [Google Scholar] [CrossRef]
- Truscott, R.; Augusteyn, R. The state of sulphydryl groups in normal and cataractous human lenses. Exp. Eye Res. 1977, 25, 139–148. [Google Scholar] [CrossRef]
- Takemoto, L. Increase in the Intramolecular Disulfide Bonding of Alpha-A Crystallin during Aging of the Human Lens. Exp. Eye Res. 1996, 63, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, L.J. Oxidation of Cysteine Residues from Alpha-A Crystallin during Cataractogenesis of the Human Lens. Biochem. Biophys. Res. Commun. 1996, 223, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Chamorro, M.; Smith, D.L.; Smith, J.B. Identification of the major components of the high molecular weight crystalline from old human lenses. Curr. Eye Res. 1994, 13, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.; Huang, Q.-L.; Horwitz, J.; Zigler, J. Evidence that α-crystallin prevents non-specific protein aggregation in the intact eye lens. Biochim. Biophys. Acta Gen. Subj. 1995, 1245, 439–447. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timsina, R.; Mainali, L. Association of Alpha-Crystallin with Fiber Cell Plasma Membrane of the Eye Lens Accompanied by Light Scattering and Cataract Formation. Membranes 2021, 11, 447. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11060447
Timsina R, Mainali L. Association of Alpha-Crystallin with Fiber Cell Plasma Membrane of the Eye Lens Accompanied by Light Scattering and Cataract Formation. Membranes. 2021; 11(6):447. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11060447
Chicago/Turabian StyleTimsina, Raju, and Laxman Mainali. 2021. "Association of Alpha-Crystallin with Fiber Cell Plasma Membrane of the Eye Lens Accompanied by Light Scattering and Cataract Formation" Membranes 11, no. 6: 447. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11060447