
Extracellular Vesicles in Cervical Cancer and HPV Infection

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Protein Cargo in Extracellular Vesicles
3. Genomic DNA and Human Papillomavirus DNA in Extracellular Vesicles
4. RNA in Extracellular Vesicles

5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bebelman, M.; Smit, M.J.; Pegtel, D.M.; Baglio, S.R. Biogenesis and function of extracellular vesicles in cancer. Pharmacol. Ther. 2018, 188, 1–11. [Google Scholar] [CrossRef]
- Minciacchi, V.R.; Freeman, M.R.; Di Vizio, D. Extracellular vesicles in cancer: Exosomes, microvesicles and the emerging role of large oncosomes. Semin. Cell Dev. Biol. 2015, 40, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLO-BOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Walboomers, J.M.; Jacobs, M.V.; Manos, M.M.; Bosch, F.X.; Kummer, J.A.; Shah, K.V.; Snijders, P.J.; Peto, J.; Meijer, C.J.; Muñoz, N. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J. Pathol. 1999, 189, 12–19. [Google Scholar] [CrossRef]
- Castellsagué, X.; Muñoz, N. Chapter 3: Cofactors in human papillomavirus carcinogenesis—Role of parity, oral contraceptives, and tobacco smoking. J. Natl. Cancer Inst. Monogr. 2003, 2003, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guenat, D.; Hermetet, F.; Prétet, J.L.; Mougin, C. Exosomes and other extracellular vesicles in HPV transmission and car-cinogenesis. Viruses 2017, 9, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chargaff, E.; West, R. The biological significance of the thromboplastic protein of blood. J. Biol. Chem. 1946, 166, 189–197. [Google Scholar] [CrossRef]
- Harding, C.; Heuser, J.; Stahl, P. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J. Cell Biol. 1983, 97, 329–339. [Google Scholar] [CrossRef]
- Chiang, C.Y.; Chen, C. Toward characterizing extracellular vesicles at a single-particle level. J. BioMed Sci. 2019, 26, 9. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, T.L. Tumor-derived exosomes and their role in cancer progression. Adv. Appl. Microbiol. 2016, 74, 103–141. [Google Scholar] [CrossRef] [Green Version]
- Kowal, J.; Tkach, M. Dendritic cell extracellular vesicles. Biol. Endoplasmic Reticulum 2019, 349, 213–249. [Google Scholar] [CrossRef]
- Yokoi, A.; Villar-Prados, A.; Oliphint, P.A.; Zhang, J.; Song, X.; De Hoff, P.; Morey, R.; Liu, J.; Roszik, J.; Clise-Dwyer, K.; et al. Mechanisms of nuclear content loading to exosomes. Sci. Adv. 2019, 5, eaax8849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of MicroRNAs in living cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [Green Version]
- Villarroya-Beltri, C.; Gutiérrez-Vázquez, C.; Sánchez-Cabo, F.; Pérez-Hernández, D.; Vázquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sánchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppers-Lalic, D.; Hackenberg, M.; Bijnsdorp, I.V.; van Eijndhoven, M.A.; Sadek, P.; Sie, D.; Zini, N.; Middeldorp, J.; Ylstra, B.; de Menezes, R.X.; et al. Nontemplated nucleotide additions distinguish the small RNA composition in cells from exosomes. Cell Rep. 2014, 8, 1649–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berti, F.C.B.; Salviano-Silva, A.; Beckert, H.C.; de Oliveira, K.B.; Cipolla, G.A.; Malheiros, D. From squamous intraepithelial lesions to cervical cancer: Circulating microRNAs as potential biomarkers in cervical carcinogenesis. Biochim. Biophys. Acta BBA Bioenerg. 2019, 1872, 188306. [Google Scholar] [CrossRef]
- Chiantore, M.V.; Mangino, G.; Iuliano, M.; Capriotti, L.; Di Bonito, P.; Fiorucci, G.; Romeo, G. Human papillomavirus and carcinogenesis: Novel mechanisms of cell communication involving extracellular vesicles. Cytokine Growth Factor Rev. 2020, 51, 92–98. [Google Scholar] [CrossRef]
- Nahand, J.S.; Moghoofei, M.; Salmaninejad, A.; Bahmanpour, Z.; Karimzadeh, M.; Nasiri, M.; Mirzaei, H.R.; Pourhanifeh, M.H.; Bokharaei-Salim, F.; Mirzaei, H.; et al. Pathogenic role of exosomes and microRNAs in HPV-mediated inflammation and cervical cancer: A review. Int. J. Cancer 2020, 146, 305–320. [Google Scholar] [CrossRef]
- Khan, S.; Jutzy, J.M.S.; Aspe, J.R.; McGregor, D.W.; Neidigh, J.W.; Wall, N.R. Survivin is released from cancer cells via exosomes. Apoptosis 2010, 16, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honegger, A.; Leitz, J.; Bulkescher, J.; Hoppe-Seyler, K.; Hoppe-Seyler, F. Silencing of human papillomavirus (HPV) E6/E7oncogene expression affects both the contents and the amounts of extracellular microvesicles released from HPV-positive cancer cells. Int. J. Cancer 2013, 133, 1631–1642. [Google Scholar] [CrossRef] [PubMed]
- Ranjit, S.; Kodidela, S.; Sinha, N.; Chauhan, S.; Kumar, S. Extracellular vesicles from human papilloma virus-infected cervical cancer cells enhance HIV-1 replication in differentiated U1 cell line. Viruses 2020, 12, 239. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.-J.; Yang, Y.; Wei, W.-F.; Wu, X.-G.; Yan, R.-M.; Zhou, C.-F.; Chen, X.-J.; Wu, S.; Wang, W.; Fan, L.-S. Tumor-secreted exosomal Wnt2B activates fibroblasts to promote cervical cancer progression. Oncogenesis 2021, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhang, C.; Xiang, P.; Shen, J.; Sun, W.; Yu, H. Exosomes derived from HeLa cells break down vascular integrity by triggering endoplasmic reticulum stress in endothelial cells. J. Extracell. Vesicles 2020, 9, 1722385. [Google Scholar] [CrossRef] [Green Version]
- Flanagan, J.; Middeldorp, J.; Sculley, T. Localization of the Epstein–Barr virus protein LMP 1 to exosomes. J. Gen. Virol. 2003, 84, 1871–1879. [Google Scholar] [CrossRef]
- Kapoor, N.R.; Chadha, R.; Kumar, S.; Choedon, T.; Reddy, V.S.; Kumar, V. The HBx gene of hepatitis B virus can influence hepatic microenvironment via exosomes by transferring its mRNA and protein. Virus Res. 2017, 240, 166–174. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, X.; Yu, Q.; He, J.J. Exosome-associated hepatitis C virus in cell cultures and patient plasma. Biochem. Biophys. Res. Commun. 2014, 455, 218–222. [Google Scholar] [CrossRef]
- Cone, A.S.; York, S.B.; Meckes, D.G. Extracellular vesicles in Epstein-Barr virus pathogenesis. Curr. Clin. Microbiol. Rep. 2019, 6, 121–131. [Google Scholar] [CrossRef]
- Kim, S.M. Human papilloma virus in oral cancer. J. Korean Assoc. Oral Maxillofac. Surg. 2016, 42, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, S.; Sharma, P.; Theodoraki, M.-N.; Pietrowska, M.; Yerneni, S.S.; Lang, S.; Ferrone, S.; Whiteside, T.L. Molecular and functional profiles of exosomes from HPV(+) and HPV(−) head and neck cancer cell lines. Front. Oncol. 2018, 8, 445. [Google Scholar] [CrossRef]
- Zhang, J.; Burn, C.; Young, K.; Wilson, M.; Ly, K.; Budhwani, M.; Tschirley, A.; Braithwaite, A.; Baird, M.; Hibma, M. Micro-particles produced by human papillomavirus type 16 E7-expressing cells impair antigen presenting cell function and the cyto-toxic T cell response. Sci Rep. 2018, 8, 2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannan, A.; Hertweck, K.L.; Philley, J.V.; Wells, R.B.; Dasgupta, S. Genetic mutation and exosome signature of human papilloma virus associated oropharyngeal cancer. Sci. Rep. 2017, 7, srep46102. [Google Scholar] [CrossRef] [PubMed]
- Guescini, M.; Genedani, S.; Stocchi, V.; Agnati, L.F. Astrocytes and glioblastoma cells release exosomes carrying mtDNA. J. Neural Transm. 2010, 117, 1–4. [Google Scholar] [CrossRef]
- Balaj, L.; Lessard, R.; Dai, L.; Cho, Y.-J.; Pomeroy, S.L.; Breakefield, X.O.; Skog, J. Tumour microvesicles contain retrotransposon elements and amplified oncogene sequences. Nat. Commun. 2011, 2, 180. [Google Scholar] [CrossRef] [PubMed]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-stranded DNA in exosomes: A novel biomarker in cancer detection. Cell Res. 2014, 24, 766–769. [Google Scholar] [CrossRef] [Green Version]
- Thippabhotla, S.; Zhong, C.; He, M. 3D cell culture stimulates the secretion of in vivo like extracellular vesicles. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Campitelli, M.; Jeannot, E.; Peter, M.; Lappartient, E.; Saada, S.; de la Rochefordière, A.; Fourchotte, V.; Alran, S.; Petrow, P.; Cottu, P.; et al. Human papillomavirus mutational insertion: Specific marker of circulating tumor DNA in cervical cancer pa-tients. PLoS ONE 2012, 7, e43393. [Google Scholar] [CrossRef]
- Cocuzza, C.E.; Martinelli, M.; Sina, F.; Piana, A.; Sotgiu, G.; Dell’Anna, T.; Musumeci, R. Human papillomavirus DNA detection in plasma and cervical samples of women with a recent history of low grade or precancerous cervical dysplasia. PLoS ONE 2017, 12, e0188592. [Google Scholar] [CrossRef]
- De Carolis, S.; Pellegrini, A.; Santini, D.; Ceccarelli, C.; De Leo, A.; Alessandrini, F.; Arienti, C.; Pignatta, S.; Tesei, A.; Mantovani, V.; et al. Liquid biopsy in the diagnosis of HPV DNA in breast lesions. Future Microbiol. 2018, 13, 187–194. [Google Scholar] [CrossRef] [Green Version]
- De Carolis, S.; Storci, G.; Ceccarelli, C.; Savini, C.; Gallucci, L.; Sansone, P.; Santini, D.; Seracchioli, R.; Taffurelli, M.; Fabbri, F.; et al. HPV DNA associates with breast cancer malignancy and it is transferred to breast cancer stromal cells by extracellular vesicles. Front. Oncol. 2019, 9, 860. [Google Scholar] [CrossRef] [Green Version]
- Ambrosio, M.R.; Vernillo, R.; De Carolis, S.; Carducci, A.; Mundo, L.; Ginori, A.; Rocca, B.J.; Nardone, V.; Fei, A.L.; Carfagno, T.; et al. Putative role of circulating human papillomavirus DNA in the development of primary squamous cell carcinoma of the middle rectum: A case report. Front. Oncol. 2019, 9, 93. [Google Scholar] [CrossRef]
- Mata-Rocha, M.; Rodríguez-Hernández, R.M.; Chávez-Olmos, P.; Garrido, E.; Robles-Vázquez, C.; Aguilar-Ruiz, S.; Torres-Aguilar, H.; González-Torres, C.; Gaytan-Cervantes, J.; Mejía-Aranguré, J.M.; et al. Presence of HPV DNA in extracellular vesicles from HeLa cells and cervical samples. Enferm. Infecc. Microbiol. Clin. 2020, 38, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Tong, F.; Andress, A.; Tang, G.; Liu, P.; Wang, X. Comprehensive profiling of extracellular RNA in HPV-induced cancers using an improved pipeline for small RNA-seq analysis. Sci. Rep. 2020, 10, 19450. [Google Scholar] [CrossRef]
- Schageman, J.; Zeringer, E.; Li, M.; Barta, T.; Lea, K.; Gu, J.; Magdaleno, S.; Setterquist, R.; Vlassov, A.V. The complete exosome workflow solution: From isolation to characterization of RNA cargo. BioMed Res. Int. 2013, 2013, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Hou, L.; Ma, Y.; Zhou, L.; Wang, F.; Cheng, B.; Wang, W.; Lu, B.; Liu, P.; Lu, W.; et al. Exosomal let-7d-3p and miR-30d-5p as diagnostic biomarkers for non-invasive screening of cervical cancer and its precursors. Mol. Cancer 2019, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Honegger, A.; Schilling, D.; Bastian, S.; Sponagel, J.; Kuryshev, V.; Sültmann, H.; Scheffner, M.; Hoppe-Seyler, K.; Hoppe-Seyler, F. Dependence of intracellular and exosomal microRNAs on viral E6/E7 oncogene expression in HPV-positive tumor cells. PLOS Pathog. 2015, 11, e1004712. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, H.; Wang, X.; Yu, Q.; Li, S.; Yu, X.; Gong, W. Increased exosomal MicroRNA-21 and MicroRNA-146a levels in the cervicovaginal lavage specimens of patients with cervical cancer. Int. J. Mol. Sci. 2014, 15, 758–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, G.; Song, G.; Zou, X.; Shan, X.; Liu, Q.; Xia, T.; Zhou, X.; Zhu, W. Circulating plasma microRNA signature for the diagnosis of cervical cancer. Cancer Biomark. 2019, 26, 491–500. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Guo, X.; Hu, Z.; Shi, H. Interfering human papillomavirus E6/E7 oncogenes in cervical cancer cells inhibits the angiogenesis of vascular endothelial cells via increasing miR-377 in cervical cancer cell-derived microvesicles. OncoTargets Ther. 2020, 13, 4145–4155. [Google Scholar] [CrossRef]
- Chiantore, M.V.; Mangino, G.; Iuliano, M.; Zangrillo, M.S.; De Lillis, I.; Vaccari, G.; Accardi, R.; Tommasino, M.; Columba Cabezas, S.; Federico, M.; et al. Human papillomavirus E6 and E7 oncoproteins affect the expression of cancer-related mi-croRNAs: Additional evidence in HPV-induced tumorigenesis. J. Cancer Res. Clin. Oncol. 2016, 142, 1751–1763. [Google Scholar] [CrossRef]
- Harden, M.E.; Munger, K. Human papillomavirus 16 E6 and E7 oncoprotein expression alters microRNA expression in ex-tracellular vesicles. Virology 2017, 508, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, X.; Meng, L.; Li, W.; Li, C.; Li, P.; Xu, S. Changes of miRNA expression profiles from cervical-vaginal fluid-derived exosomes in response to HPV16 infection. BioMed Res. Int. 2020, 2020, 7046894. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.G.; Zhou, C.F.; Zhang, Y.M.; Yan, R.M.; Wei, W.F.; Chen, X.J.; Yi, H.Y.; Liang, L.J.; Fan, L.S.; Liang, L.; et al. Cancer-derived exosomal miR-221-3p promotes angiogenesis by targeting THBS2 in cervical squamous cell carcinoma. Angiogenesis 2019, 22, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.-F.; Ma, J.; Huang, L.; Yi, H.-Y.; Zhang, Y.-M.; Wu, X.-G.; Yan, R.-M.; Liang, L.; Zhong, M.; Yu, Y.-H.; et al. Cervical squamous cell carcinoma-secreted exosomal miR-221-3p promotes lymphangiogenesis and lymphatic metastasis by targeting VASH1. Oncogene 2019, 38, 1256–1268. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, H.; Yuan, M.; Li, M.; Zhang, S. Cervical cancer cells-secreted exosomal microRNA-221-3p promotes invasion, migration and angiogenesis of microvascular endothelial cells in cervical cancer by down-regulating MAPK10 expression. Cancer Manag. Res. 2019, 11, 10307–10319. [Google Scholar] [CrossRef] [Green Version]
- You, X.; Wang, Y.; Meng, J.; Han, S.; Liu, L.; Sun, Y.; Zhang, J.; Sun, S.; Li, X.; Sun, W.; et al. Exosomal miR-663b exposed to TGF-β1 promotes cervical cancer metastasis and epithelial-mesenchymal transition by targeting MGAT3. Oncol. Rep. 2021, 45, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jiang, M.; Qian, L.; Lin, X.; Song, W.; Gao, Y.; Zhou, Y. The STAT3-miR-223-TGFBR3/HMGCS1 axis modulates the progression of cervical carcinoma. Mol. Oncol. 2020, 14, 2313–2331. [Google Scholar] [CrossRef]
- Zhou, C.; Wei, W.; Ma, J.; Yang, Y.; Liang, L.; Zhang, Y.; Wang, Z.; Chen, X.; Huang, L.; Wang, W.; et al. Cancer-secreted exosomal miR-1468-5p promotes tumor immune escape via the immunosuppressive reprogramming of lymphatic vessels. Mol. Ther. 2021, 29, 1512–1528. [Google Scholar] [CrossRef]
- Gezer, U.; Özgür, E.; Cetinkaya, M.; Isin, M.; Dalay, N. Long non-coding RNAs with low expression levels in cells are enriched in secreted exosomes. Cell Biol. Int. 2014, 38, 1076–1079. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.-C.; Luo, X.-H.; Tao, G.-X.; Guan, M.; Yuan, H.; Hu, D.-K. Exosomal long noncoding RNAs are differentially expressed in the cervicovaginal lavage samples of cervical cancer patients. J. Clin. Lab. Anal. 2016, 30, 1116–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.Z.; Zhang, S.Q.; Deng, X.L.; Qiang, J.H. Serum exosomal lncRNA DLX6-AS1 is a promising biomarker for prognosis prediction of cervical cancer. Technol. Cancer Res. Treat. 2021, 20, 1533033821990060. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Mou, Q. Exosomal taurine up-regulated 1 promotes angiogenesis and endothelial cell proliferation in cervical cancer. Cancer Biol. Ther. 2020, 21, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Wei, J.; Yang, F.-L.; Pang, X.-X.; Shi, F.; Wei, Y.-X.; Liao, B.-Y.; Wang, J.-L. Exosomal lncRNA HNF1A-AS1 affects cisplatin resistance in cervical cancer cells through regulating microRNA-34b/TUFT1 axis. Cancer Cell Int. 2019, 19, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Liu, X.; Du, B.; Liu, X.; Xue, M.; Yan, Q.; Wang, X.; Wang, Q. LncRNA LINC01305 promotes cervical cancer progression through KHSRP and exosome-mediated transfer. Aging 2021, 13. [Google Scholar] [CrossRef]
- Wang, H.; Wei, M.; Kang, Y.; Xing, J.; Zhao, Y. Circular RNA circ_PVT1 induces epithelial-mesenchymal transition to promote metastasis of cervical cancer. Aging 2020, 12, 20139–20151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhou, Q.; Wei, Y.; Da, M.; Zhang, C.; Zhong, J.; Liu, J.; Shen, J. The exosome-mediated PI3k/Akt/mTOR signaling pathway in cervical cancer. Int. J. Clin. Exp. Pathol. 2019, 12, 2474–2484. [Google Scholar] [PubMed]
- Iuliano, M.; Mangino, G.; Chiantore, M.V.; Zangrillo, M.S.; Accardi, R.; Tommasino, M.; Fiorucci, G.; Romeo, G. Human papillomavirus E6 and E7 oncoproteins affect the cell microenvironment by classical secretion and extracellular vesicles delivery of inflammatory mediators. Cytokine 2018, 106, 182–189. [Google Scholar] [CrossRef]
- Qiu, J.-J.; Sun, S.-G.; Tang, X.-Y.; Lin, Y.-Y.; Hua, K.-Q. Extracellular vesicular Wnt7b mediates HPV E6-induced cervical cancer angiogenesis by activating the β-catenin signaling pathway. J. Exp. Clin. Cancer Res. 2020, 39, 1–17. [Google Scholar] [CrossRef]
- Wojakowska, A.; Zebrowska, A.; Skowronek, A.; Rutkowski, T.; Polanski, K.; Widlak, P.; Marczak, L.; Pietrowska, M. Metabolic profiles of whole serum and serum-derived exosomes are different in head and neck cancer patients treated by radio-therapy. J. Pers. Med. 2020, 10, 229. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Gillespie, D.G.; Reichert, T.E.; Jackson, E.K.; Whiteside, T.L. Purine metabolites in tumor-derived exosomes may facilitate immune escape of head and neck squamous cell carcinoma. Cancers 2020, 12, 1602. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
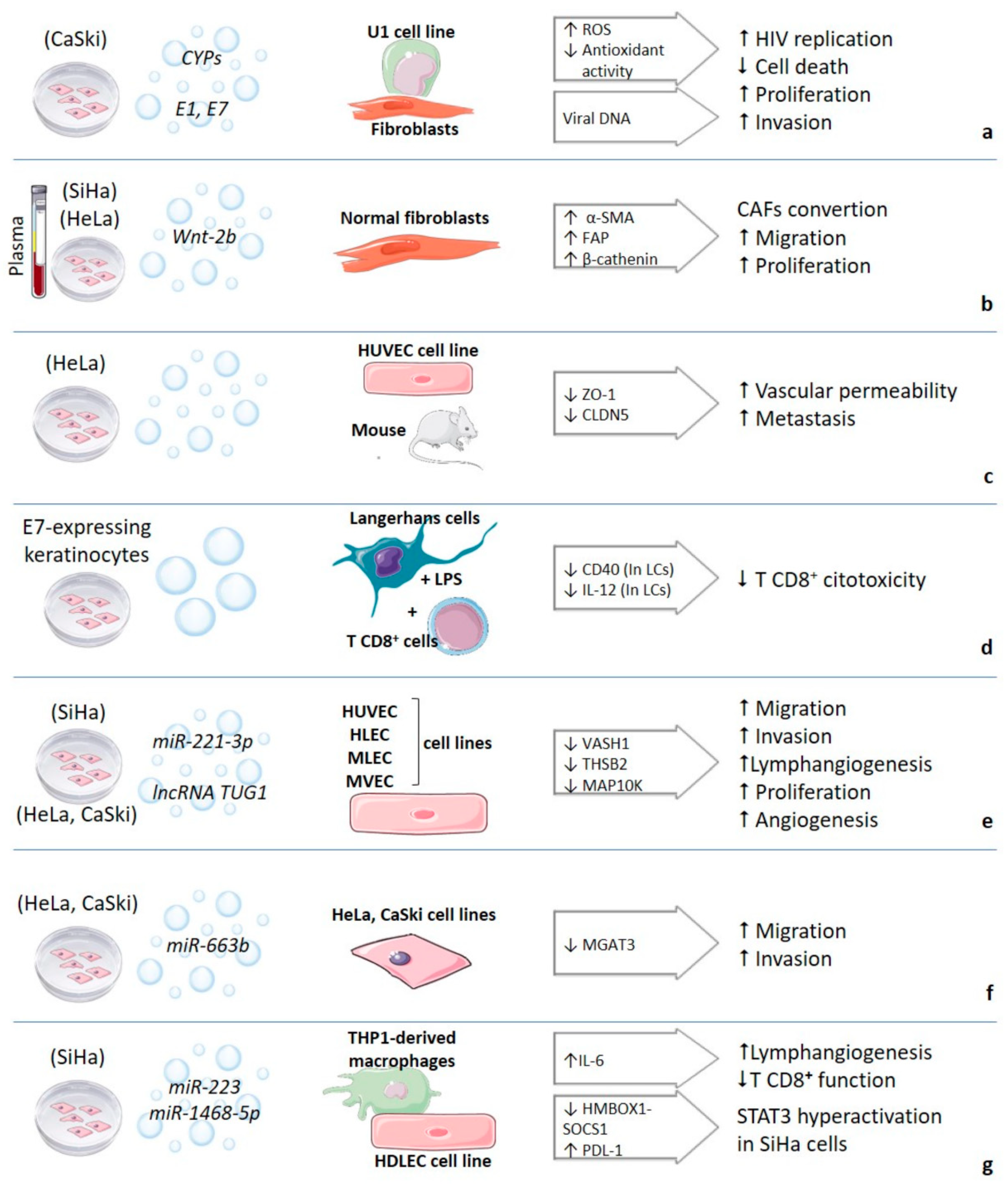{kind=link}
| Proteins in EVs from CC and Other HPV+ Cancer | |||
| EVs Type/Source | HPV | Cargo | Reference |
| Exo HeLa | HPV18 | Survivin | Khan et al., 2011 |
| MVs HeLa | HPV18 | XIAP, c-IAP1, Livin, Survivin | Honegger et al., 2013 |
| Exo Serum from patients with oropharyngeal cancer | HPV16 | MUC16, SIRPA, E7 * | Kannan et al., 2017 |
| Exo UM-SCC-104 | |||
| Exo UM-SCC-2, UM-SCC-47, UPCI: SCC-90 | HPV16 | p16, Survivin, Rb, Cyclin D1, p53, SHP-2, TGF-β, FasL, OX40, OX40L, E6 *, E7 * | Ludwig et al., 2018 |
| EVs CaSki | HPV16 | CYP1A1, CYP1B1, CYP2A6, E6 * | Ranjit et al., 2020 |
| Exo HeLa | HPV18 | CHMP4B, STX-7, RPL28, and others | Lin et al., 2020 |
| Exo SiHa | HPV16 | MMP14, MMP15 | |
| Exo Serum from patients with CC | Unknown | Wnt-2b | Liang et al., 2021 |
| Exo SiHa | HPV16 | ||
| Exo HeLa | HPV18 | ||
| RNAs in EVs from CC | |||
| EVs Type/Source | HPV | Cargo | Reference |
| microRNAs | |||
| Exo HeLa vs HeLa cells | HPV18 | miR-21, miR-3160-1, miR-4739, miR-31, miR-23a, miR-24-2, miR-1273a, mir-30a, mir-451a, mir-1273g | Schageman et al., 2013 |
| Exo Cervicovaginal lavages from CC patients and HPV+ without CC patients | HPV18 | miR-21, miR-146a | Liu et al., 2014 |
| Exo HeLa | |||
| Exo E6-E7-silenced HeLa | HPV18 | miR-21-5p | Honegger et al., 2015 |
| let-7d-5p, miR-20a-5p, miR-378a-3p, miR-423-3p, miR-7-5p, miR-92a-3p | |||
| Exo E6-E7-silenced SiHa | HPV16 | miR-21-5 | |
| miR-20a-5p, miR-378a-3p, miR-423-3p, miR-7-5p, miR-92a-3p | |||
| Exo HFK transduced with E6/E7 of HPV16 | HPV16 | miR-24, miR-191, miR-200c, miR-212, miR-320, miR-342-3p, miR-483-5p, miR-222 | Chiantore et al., 2016 |
| (K16) or HPV38 (K38) | HPV38 | miR-92a, miR-320, miR-323-3p, miR-494 | |
| Exo HFK transduced with E6/E7 of HPV16 vs. HFK-E6/E7 cells | HPV16 | miR-222-3p miR-16-5p, miR-200b-3p, miR-320a, miR-378a-3p | Harden & Munger, 2017 |
| Exo HFK transduced with E6/E7 of HPV16 vs. Exo HFK | miR-16-5p, miR-18a-5p, miR-19a-3p, miR-25-3p, miR-93-5p, miR-103a-3p, miR-106b-5p, miR-107, miR-130a-3p, miR-148b-3p, miR-155-5p, miR-182-5p, miR-195-5p, miR-218-5p, miR-222-3p, miR-335-5p, miR-375, miR-625-3p, miR-652-3p let-7i-5p, miR-21-5p, miR-22-3p, miR-34a-5p, miR-151a-3p, miR-192-5p, miR-200b-3p, miR-205-5p, miR-221-3p, miR-320a, miR-376c-3p, miR-378a-3p | ||
| Exo Plasma from CC patients | Unknown | miR-146a-5p, miR-151a-3, miR-2110 | Ma et al., 2019 |
| Exo Plasma from CC and CIN II, III patients | Unknown | let-7d-3p and miR-30d-5p | Zheng et al., 2019 |
| Exo Serum from CC patients | Unknown | miR-221-3p | Zhou et al., 2019; Wu et al., 2019; Zhang L. et al., 2019 |
| Exo SiHa | HPV16 | ||
| Exo Cervicovaginal-fluid from CC patients | HPV16 | miR-6746-3p, miR-4667-5p, miR-6775-5p, miR-802, miR-4477a, miR-6815-5p, miR-3190-3p, miR-5095, miR-6865-5p, miR-6786-3p, miR-4669, miR-6829-5p, miR-3619-3p, miR-4778-5p, miR-6867-5p | Wu et al., 2020 |
| miR-363-5p, miR-621, miR-6810-3p, miR-6728-3p, miR-3178, miR-211-5p, miR-3927-5p, miR-3145-5p, miR-4322, miR-548t-5p, miR-4436a, miR-5700, miR-645, miR-299-5p, miR-1911-3p | |||
| MVs E6-E7-interfering HeLa | HPV18 | miR-377 | Zhang Y. et al., 2020 |
| Exo SiHa | HPV16 | miR-223 | Zhang J. et al., 2020 |
| Exo HPV+: SiHa, CaSki, C4I, SW756, SCC-154, HeLa, SCC-090, SCC-047 | HPV16 | miR-1306-5p, miR-193b-5p, miR-92b-3p, miR-92b-5p, miR-365b-3p, miR-125a-5p, miR-let-7b-5p | Tong et al., 2020 |
| vs Exo HPV-: UPCI-068, UPCI-017, SCC-4, SCC-1, HT-3, C-33 A | HPV18 | ||
| Exo Serum from CC patients | Unknown | miR-1468-5p | Zhou et al., 2021 |
| Exo SiHa | HPV16 | ||
| Exo HeLa | HPV18 | miR-663b | You et al., 2021 |
| Exo CaSki | HPV16 | ||
| lncRNAs | |||
| Exo HeLa cells vs HeLa cells | HPV18 | HOTAIR, lincRNA-p21, GAS5, TUG1, CCND1-ncRNA | Gezer et al., 2014 |
| Exo Cervicovaginal lavages from CC patients and HPV+ without CC patients | Unknown | HOTAIR, MALAT1 | Zhang et al., 2016 |
| MEG3 | |||
| Exo HeLa (DDP-resistant) | HPV18 | HNF1A-AS1 | Luo et al., 2019 |
| Exo HeLa | HPV18 | TUG1 | Lei & Mou, 2020 |
| Exo CaSki | HPV16 | ||
| Exo Serum from CC and CIN patients | Unknown | lncRNA DLX6-AS1 | Ding et al., 2021 |
| Exo C-33 A | Negative | LINC01305 | Huang et al., 2021 |
| CircRNA | |||
| Exo Plasma and urine from CC patients | Unknown | circ_PVT1 | Wang et al., 2020 |
| mRNAs | |||
| Exo HeLa vs HeLa cells | HPV18 | SUSD2, BRWD3, SENP6, FAM59B, TUBBA4, QRFPR, MDK, MTRNR2L2, CWC25, DUSP13 | Schageman et al., 2013 |
| Exo HFK transduced with E6/E7 of HPV16 (K16) | HPV16 | E6/E7 * | Chiantore et al., 2016 |
| Exo SiHa | |||
| EVs HFK transduced with E6/E7 of HPV16 (K16) or HPV38 (K38) vs. EVs HFK | HPV16 | CXCL10 | Iuliano et al., 2018 |
| CCL27, CCL20, CXCL3, CXCL1, IL-1α, IL-1β, angiogenin | |||
| HPV38 | CCL2, TNFα | ||
| CCL27, CXCL3 | |||
| Exo Vaginal secretions from CC patients | Unknown | PI3k, Akt, mTOR | Zhang W. et al., 2019 |
| Exo Serum from CC patients Exo HeLa and SW756 | Unknown | Wnt7b | Qiu et al., 2020 |
| HPV18 | |||
| Exo SiHa and CaSki | HPV16 | ||
| DNA in EVs from CC and other HPV+ cancer | |||
| EVs Type/Source | HPV | Cargo | Reference |
| EVs Serum from patients with breast cancer | HPV16 | E6 * | De Carolis et al., 2018 |
| EVs Serum from patients with breast cancer | HPV53 | L1 * | De Carolis et al., 2019 |
| EVs CaSki | HPV16 | E1 *, E7 * | |
| Exo Plasma from patients with colorectal cancer | HPV16 | L1 * | Ambrosio et al., 2019 |
| Exo CaSki | E1 *, E6 *, L1 * | ||
| Exo HeLa | HPV18 | E1 *, E6 *, E7 *, L1 * | Mata-Rocha et al., 2020 |
| Exo from CC patients | Unknown | E6 *, E7 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acevedo-Sánchez, V.; Rodríguez-Hernández, R.M.; Aguilar-Ruíz, S.R.; Torres-Aguilar, H.; Romero-Tlalolini, M.d.l.A. Extracellular Vesicles in Cervical Cancer and HPV Infection. Membranes 2021, 11, 453. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11060453
Acevedo-Sánchez V, Rodríguez-Hernández RM, Aguilar-Ruíz SR, Torres-Aguilar H, Romero-Tlalolini MdlA. Extracellular Vesicles in Cervical Cancer and HPV Infection. Membranes. 2021; 11(6):453. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11060453
Chicago/Turabian StyleAcevedo-Sánchez, Víctor, Ruth M. Rodríguez-Hernández, Sergio R. Aguilar-Ruíz, Honorio Torres-Aguilar, and María de los A. Romero-Tlalolini. 2021. "Extracellular Vesicles in Cervical Cancer and HPV Infection" Membranes 11, no. 6: 453. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11060453