
High-Level Expression of Palmitoylated MPP1 Recombinant Protein in Mammalian Cells

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid/DNA
2.2. Cell Culture and Transfection
2.3. Transfection Analysis
2.4. Purification of MPP1
2.5. Detection of S-Palmitoylation
2.6. SDS–PAGE and Western Blot
2.7. Circular Dichroism (CD) Spectroscopy
3. Results and Discussion
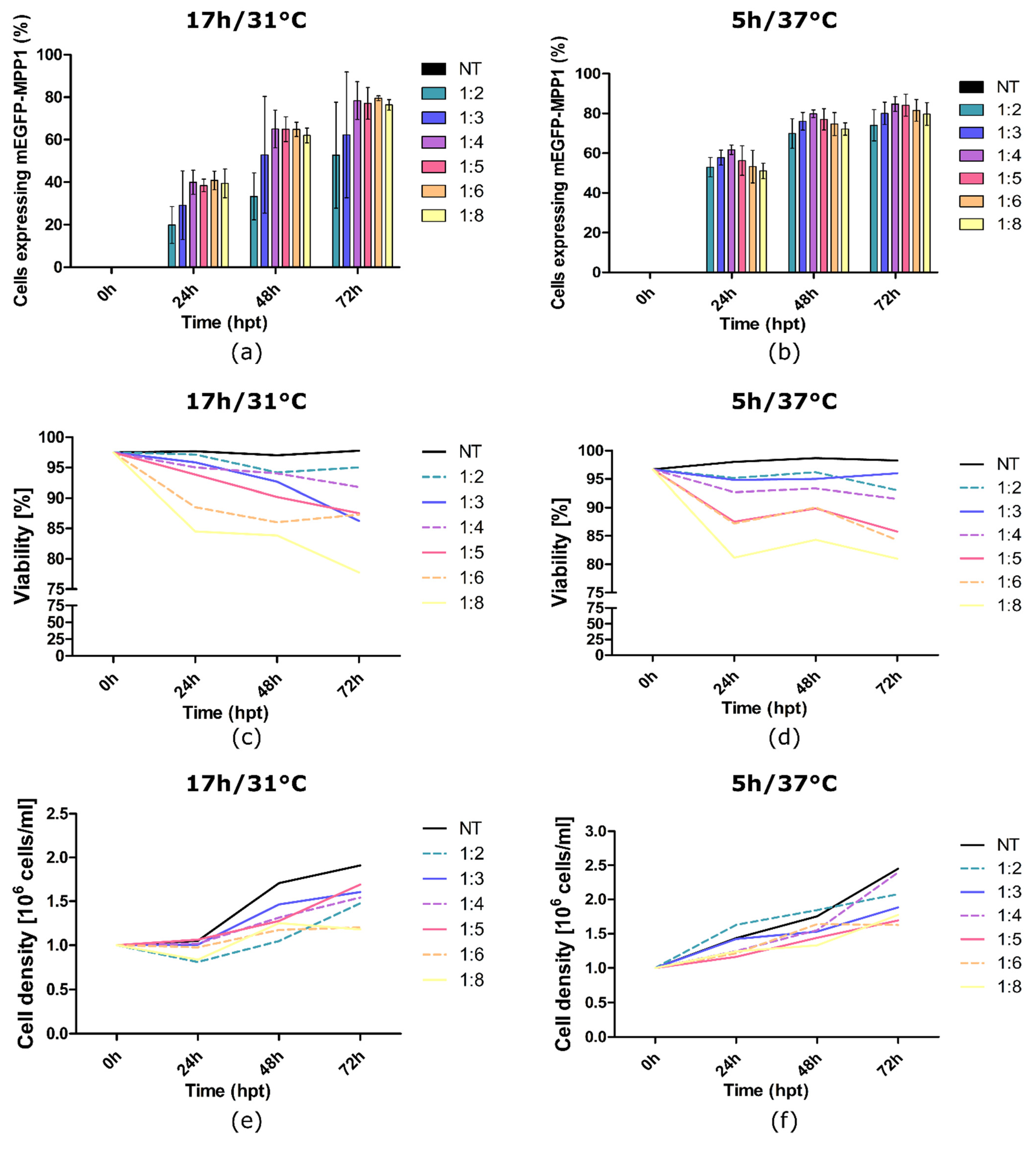
3.1. Optimization of HEK-293F Cells Transfection and MPP1 Overexpression
3.2. Protein Purification
3.3. Secondary Structure
3.4. Analysis of S-Palmitoylation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Funke, L.; Dakoji, S.; Bredt, D.S. Membrane-Associated Guanylate Kinases Regulate Adhesion and Plasticity At Cell Junctions. Annu. Rev. Biochem. 2005, 74, 219–245. [Google Scholar] [CrossRef]
- Oliva, C.; Escobedo, P.; Astorga, C.; Molina, C.; Sierralta, J. Role of the maguk protein family in synapse formation and function. Dev. Neurobiol. 2012, 72, 57–72. [Google Scholar] [CrossRef]
- Zhu, J.; Shang, Y.; Zhang, M. Mechanistic basis of MAGUK-organized complexes in synaptic development and signalling. Nat. Rev. Neurosci. 2016, 17, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Chytła, A.; Gajdzik-Nowak, W.; Olszewska, P.; Biernatowska, A.; Sikorski, A.F.; Czogalla, A. Not Just Another Scaffolding Protein Family: The Multifaceted MPPs. Molecules 2020, 25, 4954. [Google Scholar] [CrossRef] [PubMed]
- Seo, P.S.; Jeong, J.J.; Zeng, L.; Takoudis, C.G.; Quinn, B.J.; Khan, A.A.; Hanada, T.; Chishti, A.H. Alternatively spliced exon 5 of the FERM domain of protein 4.1R encodes a novel binding site for erythrocyte p55 and is critical for membrane targeting in epithelial cells. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 281–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruff, P.; Speicher, D.W.; Husain-Chishti, A. Molecular identification of a major palmitoylated erythrocyte membrane protein containing the src homology 3 motif. Proc. Natl. Acad. Sci. USA 1991, 88, 6595–6599. [Google Scholar] [CrossRef] [Green Version]
- Alloisio, N.; Dalla Venezia, N.; Rana, A.; Andrabi, K.; Texier, P.; Gilsanz, F.; Cartron, J.P.; Delaunay, J.; Chishti, A.H. Evidence that red blood cell protein p55 may participate in the skeleton- membrane linkage that involves protein 4.1 and glycophorin C. Blood 1993, 82, 1323–1327. [Google Scholar] [CrossRef] [Green Version]
- Marfatia, S.M.; Lue, R.A.; Branton, D.; Chishti, A.H. In vitro binding studies suggest a membrane-associated complex between erythroid p55, protein 4.1, and glycophorin C. J. Biol. Chem. 1994, 269, 8631–8634. [Google Scholar] [CrossRef]
- Biernatowska, A.; Podkalicka, J.; Majkowski, M.; Hryniewicz-Jankowska, A.; Augoff, K.; Kozak, K.; Korzeniewski, J.; Sikorski, A.F. The role of MPP1/p55 and its palmitoylation in resting state raft organization in HEL cells. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 1876–1884. [Google Scholar] [CrossRef] [Green Version]
- Biernatowska, A.; Augoff, K.; Podkalicka, J.; Tabaczar, S.; Gajdzik-Nowak, W.; Czogalla, A.; Sikorski, A.F. MPP1 directly interacts with flotillins in erythrocyte membrane—Possible mechanism of raft domain formation. Biochim. Biophys. Acta Biomembr. 2017, 1859, 2203–2212. [Google Scholar] [CrossRef]
- Podkalicka, J.; Biernatowska, A.; Olszewska, P.; Tabaczar, S.; Sikorski, A.F. The microdomain-organizing protein MPP1 is required for insulinstimulated activation of H-Ras. Oncotarget 2018, 9, 18410–18421. [Google Scholar] [CrossRef] [Green Version]
- Trybus, M.; Niemiec, L.; Biernatowska, A.; Hryniewicz-Jankowska, A.; Sikorski, A.F. MPP1-based mechanism of resting state raft organization in the plasma membrane. Is it a general or specialized mechanism in erythroid cells? Folia Histochem. Cytobiol. 2019, 57, 43–55. [Google Scholar] [CrossRef]
- Podkalicka, J.; Biernatowska, A.; Majkowski, M.; Grzybek, M.; Sikorski, A.F. MPP1 as a factor regulating phase separation in giant plasma membrane-derived vesicles. Biophys. J. 2015, 108, 2201–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lingwood, D.; Simons, K. Detergent resistance as a tool in membrane research. Nat. Protoc. 2007, 2, 2159–2165. [Google Scholar] [CrossRef] [PubMed]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The mystery of membrane organization: Composition, regulation and physiological relevance of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikorski, A.F.; Podkalicka, J.; Jones, W.; Biernatowska, A. Membrane Rafts in the Erythrocyte Membrane: A Novel Role of MPP1p55. In Biochemical Roles of Eukaryotic Cell Surface Macromolecules, Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2015; Volume 842, pp. 61–78. ISBN 978-3-319-11279-4. [Google Scholar]
- Banning, A.; Tomasovic, A.; Tikkanen, R. Functional Aspects of Membrane Association of Reggie/Flotillin Proteins. Curr. Protein Pept. Sci. 2011, 12, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Guillaume, E.; Comunale, F.; Do Khoa, N.; Planchon, D.; Bodin, S.; Gauthier-Rouvière, C. Flotillin microdomains stabilize cadherins at cell-cell junctions. J. Cell Sci. 2013, 126, 5284–5292. [Google Scholar] [CrossRef] [Green Version]
- Kurrle, N.; Völlner, F.; Eming, R.; Hertl, M.; Banning, A.; Tikkanen, R. Flotillins directly interact with γ-catenin and regulate epithelial cell-cell adhesion. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Kwiatkowska, K.; Matveichuk, O.V.; Fronk, J.; Ciesielska, A. Flotillins: At the intersection of protein S-palmitoylation and lipid-mediated signaling. Int. J. Mol. Sci. 2020, 21, 2283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biernatowska, A.; Olszewska, P.; Grzymajło, K.; Drabik, D.; Kraszewski, S.; Sikorski, A.F.; Czogalla, A. Molecular characterization of direct interactions between MPP1 and flotillins. Sci. Rep. 2021, 11, 14751. [Google Scholar] [CrossRef]
- Hancock, J.F. Lipid rafts: Contentious only from simplistic standpoints. Nat. Rev. Mol. Cell Biol. 2006, 7, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Levental, I.; Levental, K.R.; Heberle, F.A. Lipid Rafts: Controversies Resolved, Mysteries Remain. Trends Cell Biol. 2020, 30, 341–353. [Google Scholar] [CrossRef]
- Zhang, M.M.; Hang, H.C. Protein S-palmitoylation in cellular differentiation. Biochem. Soc. Trans. 2017, 45, 275–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Husseini, A.E.; Craven, S.E.; Chetkovich, D.M.; Firestein, B.L.; Schnell, E.; Aoki, C.; Bredt, D.S. Dual palmitoylation of PSD-95 mediates its vesiculotubular sorting, postsynaptic targeting, and ion channel clustering. J. Cell Biol. 2000, 148, 159–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnell, E.; Sizemore, M.; Karimzadegan, S.; Chen, L.; Bredt, D.S.; Nicoll, R.A. Direct interactions between PSD-95 and stargazin control synaptic AMPA receptor number. Proc. Natl. Acad. Sci. USA 2002, 99, 13902–13907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohn, H.; Park, M. Palmitoylation-mediated synaptic regulation of AMPA receptor trafficking and function. Arch. Pharm. Res. 2019, 42, 426–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulodziecka, K.; Diaz-Rohrer, B.B.; Farley, M.M.; Chan, R.B.; Di Paolo, G.; Levental, K.R.; Waxham, M.N.; Levental, I. Remodeling of the postsynaptic plasma membrane during neural development. Mol. Biol. Cell 2016, 27, 3480–3489. [Google Scholar] [CrossRef] [PubMed]
- Marfatia, S.M.; Lue, R.A.; Branton, D.; Chishti, A.H. Identification of the protein 4.1 binding interface on glycophorin C and p55, a homologue of the Drosophila discs-large tumor suppressor protein. J. Biol. Chem. 1995, 270, 715–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, D. Overview of Protein Expression by Mammalian Cells. Curr. Protoc. Protein Sci. 1997, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.; Yuan, P.; Vavilala, D.; Fox, M. Optimization of Protein Expression in Mammalian Cells. Curr. Protoc. Protein Sci. 2019, 95, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Hossler, P.; Khattak, S.F.; Li, Z.J. Optimal and consistent protein glycosylation in mammalian cell culture. Glycobiology 2009, 19, 936–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, F.D.; Oliveira, J.E.; Freire, R.P.; Suzuki, M.F.; Soares, C.R.; Bartolini, P. Expression of glycosylated human prolactin in HEK293 cells and related N-glycan composition analysis. AMB Express 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Kozak, S.; Bloch, Y.; De Munck, S.; Mikula, A.; Bento, I.; Savvides, S.N.; Meijers, R. Homogeneously N-glycosylated proteins derived from the GlycoDelete HEK293 cell line enable diffraction-quality crystallogenesis. Acta Crystallogr. Sect. D Struct. Biol. 2020, 76, 1244–1255. [Google Scholar] [CrossRef]
- Gutiérrez-Granados, S.; Cervera, L.; Kamen, A.A.; Gòdia, F. Advancements in mammalian cell transient gene expression (TGE) technology for accelerated production of biologics. Crit. Rev. Biotechnol. 2018, 38, 918–940. [Google Scholar] [CrossRef] [PubMed]
- Sunley, K.; Butler, M. Strategies for the enhancement of recombinant protein production from mammalian cells by growth arrest. Biotechnol. Adv. 2010, 28, 385–394. [Google Scholar] [CrossRef]
- Fuenmayor, J.; Cervera, L.; Gutiérrez-Granados, S.; Gòdia, F. Transient gene expression optimization and expression vector comparison to improve HIV-1 VLP production in HEK293 cell lines. Appl. Microbiol. Biotechnol. 2018, 102, 165–174. [Google Scholar] [CrossRef]
- Delafosse, L.; Xu, P.; Durocher, Y. Comparative study of polyethylenimines for transient gene expression in mammalian HEK293 and CHO cells. J. Biotechnol. 2016, 227, 103–111. [Google Scholar] [CrossRef]
- Rajendra, Y.; Kiseljak, D.; Baldi, L.; Wurm, F.M.; Hacker, D.L. Transcriptional and post-transcriptional limitations of high-yielding, PEI-mediated transient transfection with CHO and HEK-293E cells. Biotechnol. Prog. 2015, 31, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Xinyong, G.; Xianjin, C.; Yayu, W. PEI/DNA formation affects transient gene expression in suspension Chinese hamster ovary cells via a one-step transfection process. Cytotechnology 2013, 65, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Lai, W.F. In vivo nucleic acid delivery with PEI and its derivatives: Current status and perspectives. Expert Rev. Med. Devices 2011, 8, 173–185. [Google Scholar] [CrossRef]
- Won, Y.-Y.; Sharma, R.; Konieczny, S.F. Missing pieces in understanding the intracellular trafficking of polycation/DNA complexes. J. Control. Release 2009, 139, 88–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, J.; Wirtz, D.; Hanes, J. Efficient active transport of gene nanocarriers to the cell nucleus. Proc. Natl. Acad. Sci. USA 2003, 100, 3878–3882. [Google Scholar] [CrossRef] [Green Version]
- Brunner, S.; Sauer, T.; Carotta, S.; Cotten, M.; Saltik, M.; Wagner, E. Cell cycle dependence of gene transfer by lipoplex polyplex and recombinant adenovirus. Gene Ther. 2000, 7, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Cervera, L.; Gutiérrez-Granados, S.; Martínez, M.; Blanco, J.; Gòdia, F.; Segura, M.M. Generation of HIV-1 Gag VLPs by transient transfection of HEK 293 suspension cell cultures using an optimized animal-derived component free medium. J. Biotechnol. 2013, 166, 152–165. [Google Scholar] [CrossRef]
- Tom, R.; Bisson, L.; Durocher, Y. Transfection of HEK293-EBNA1 cells in suspension with linear PEI for production of recombinant proteins. Cold Spring Harb. Protoc. 2008, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Sang, Y.; Xie, K.; Mu, Y.; Lei, Y.; Zhang, B.; Xiong, S.; Chen, Y.; Qi, N. Salt ions and related parameters affect PEI–DNA particle size and transfection efficiency in Chinese hamster ovary cells. Cytotechnology 2015, 67, 67–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Domínguez, I.; Grimaldi, N.; Cervera, L.; Ventosa, N.; Gòdia, F. Impact of physicochemical properties of DNA/PEI complexes on transient transfection of mammalian cells. New Biotechnol. 2019, 49, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Daramola, O.; Stevenson, J.; Dean, G.; Hatton, D.; Pettman, G.; Holmes, W.; Field, R. A high-yielding CHO transient system: Coexpression of genes encoding EBNA-1 and GS enhances transient protein expression. Biotechnol. Prog. 2014, 30, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Bono, N.; Ponti, F.; Mantovani, D.; Candiani, G. Non-viral in vitro gene delivery: It is now time to set the bar! Pharmaceutics 2020, 12, 183. [Google Scholar] [CrossRef] [Green Version]
- Listowski, M.A.; Leluk, J.; Kraszewski, S.; Sikorski, A.F. Cholesterol interaction with the MAGUK protein family member, MPP1, via CRAC and CRAC-like motifs: An in silico docking analysis. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Micsonai, A.; Bulyáki, É.; Kardos, J. BeStSel: From Secondary Structure Analysis to Protein Fold Prediction by Circular Dichroism Spectroscopy. Methods Mol. Biol. 2021, 2199, 175–189. [Google Scholar] [CrossRef]
- Forrester, M.T.; Hess, D.T.; Thompson, J.W.; Hultman, R.; Moseley, M.A.; Stamler, J.S.; Casey, P.J. Site-specific analysis of protein S-acylation by resin-assisted capture. J. Lipid Res. 2011, 52, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.R.; Wang, C.; Adibekian, A.; Tully, S.E.; Cravatt, B.F. Global profiling of dynamic protein palmitoylation. Nat. Methods 2012, 9, 84–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, S.J.; Cheung See Kit, M.; Martin, B.R. Protein depalmitoylases. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 83–98. [Google Scholar] [CrossRef]
- Gök, C.; Plain, F.; Robertson, A.D.; Howie, J.; Baillie, G.S.; Fraser, N.J.; Fuller, W. Dynamic Palmitoylation of the Sodium-Calcium Exchanger Modulates Its Structure, Affinity for Lipid-Ordered Domains, and Inhibition by XIP. Cell Rep. 2020, 31. [Google Scholar] [CrossRef] [PubMed]
- Kokkola, T.; Kruse, C.; Roy-Pogodzik, E.M.; Pekkinen, J.; Bauch, C.; Hönck, H.H.; Hennemann, H.; Kreienkamp, H.J. Somatostatin receptor 5 is palmitoylated by the interacting ZDHHC5 palmitoyltransferase. FEBS Lett. 2011, 585, 2665–2670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, L.; McClafferty, H.; Jeffries, O.; Shipston, M.J. Multiple palmitoyltransferases are required for palmitoylation-dependent regulation of large conductance calcium- and voltage-activated potassium channels. J. Biol. Chem. 2010, 285, 23954–23962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMichael, T.M.; Zhang, L.; Chemudupati, M.; Hach, J.C.; Kenney, A.D.; Hang, H.C.; Yount, J.S. The palmitoyltransferase ZDHHC20 enhances interferon-induced transmembrane protein 3 (IFITM3) palmitoylation and antiviral activity. J. Biol. Chem. 2017, 292, 21517–21526. [Google Scholar] [CrossRef] [Green Version]
- Bolland, D.E.; Moritz, A.E.; Stanislowski, D.J.; Vaughan, R.A.; Foster, J.D. Palmitoylation by Multiple DHHC Enzymes Enhances Dopamine Transporter Function and Stability. ACS Chem. Neurosci. 2019, 10, 2707–2717. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chytła, A.; Gajdzik-Nowak, W.; Biernatowska, A.; Sikorski, A.F.; Czogalla, A. High-Level Expression of Palmitoylated MPP1 Recombinant Protein in Mammalian Cells. Membranes 2021, 11, 715. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11090715
Chytła A, Gajdzik-Nowak W, Biernatowska A, Sikorski AF, Czogalla A. High-Level Expression of Palmitoylated MPP1 Recombinant Protein in Mammalian Cells. Membranes. 2021; 11(9):715. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11090715
Chicago/Turabian StyleChytła, Agnieszka, Weronika Gajdzik-Nowak, Agnieszka Biernatowska, Aleksander F. Sikorski, and Aleksander Czogalla. 2021. "High-Level Expression of Palmitoylated MPP1 Recombinant Protein in Mammalian Cells" Membranes 11, no. 9: 715. https://0-doi-org.brum.beds.ac.uk/10.3390/membranes11090715