A Multi-Modal MRI Analysis of Cortical Structure in Relation to Gender Dysphoria, Sexual Orientation, and Age in Adolescents

 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedures
2.3. Measures
2.3.1. Age
2.3.2. Gender Dysphoria
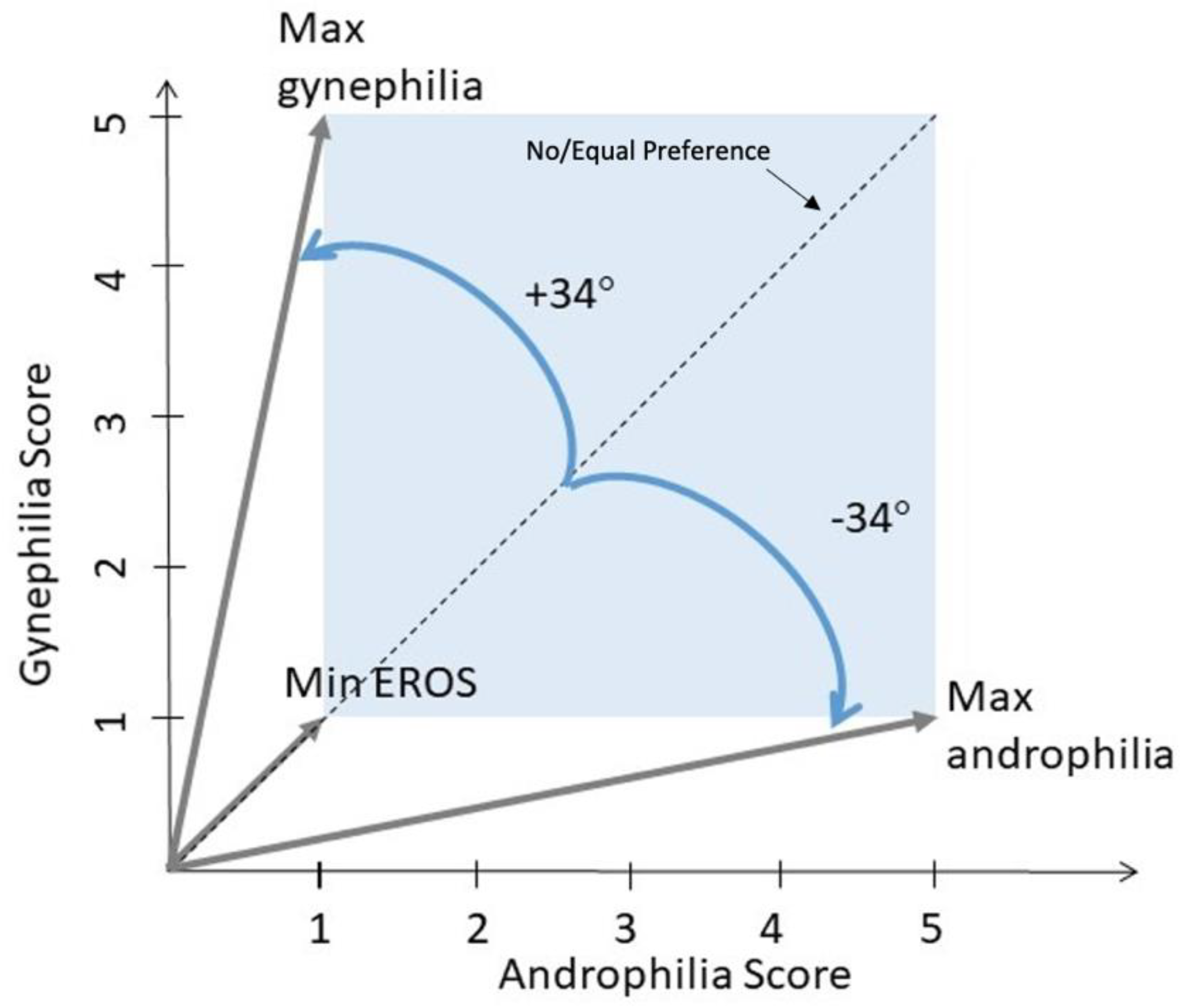
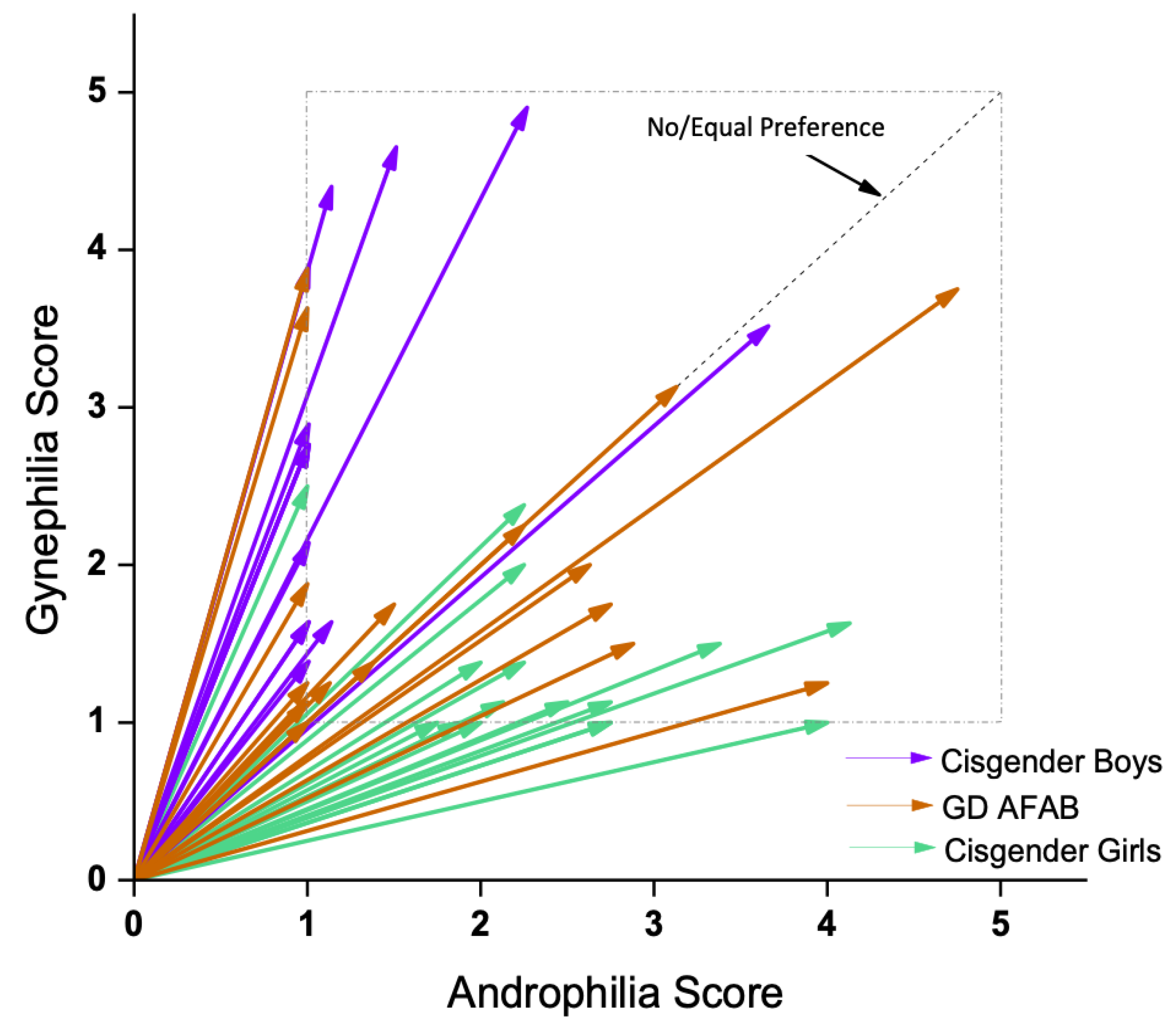
2.3.3. Sexual Orientation
2.4. Magnetic Resonance Imaging (MRI) Methods
2.4.1. Image Acquisition
2.4.2. T1w Image Processing
2.4.3. T1 Relaxation Time Map Creation
2.4.4. T1 Map Processing Pipeline
3. Statistical Analyses
3.1. Group Differences in Demographic and Psychosexual Variables
3.2. Partial Least Squares Analyses
4. Results
4.1. Demographic and Psychosexual Variables
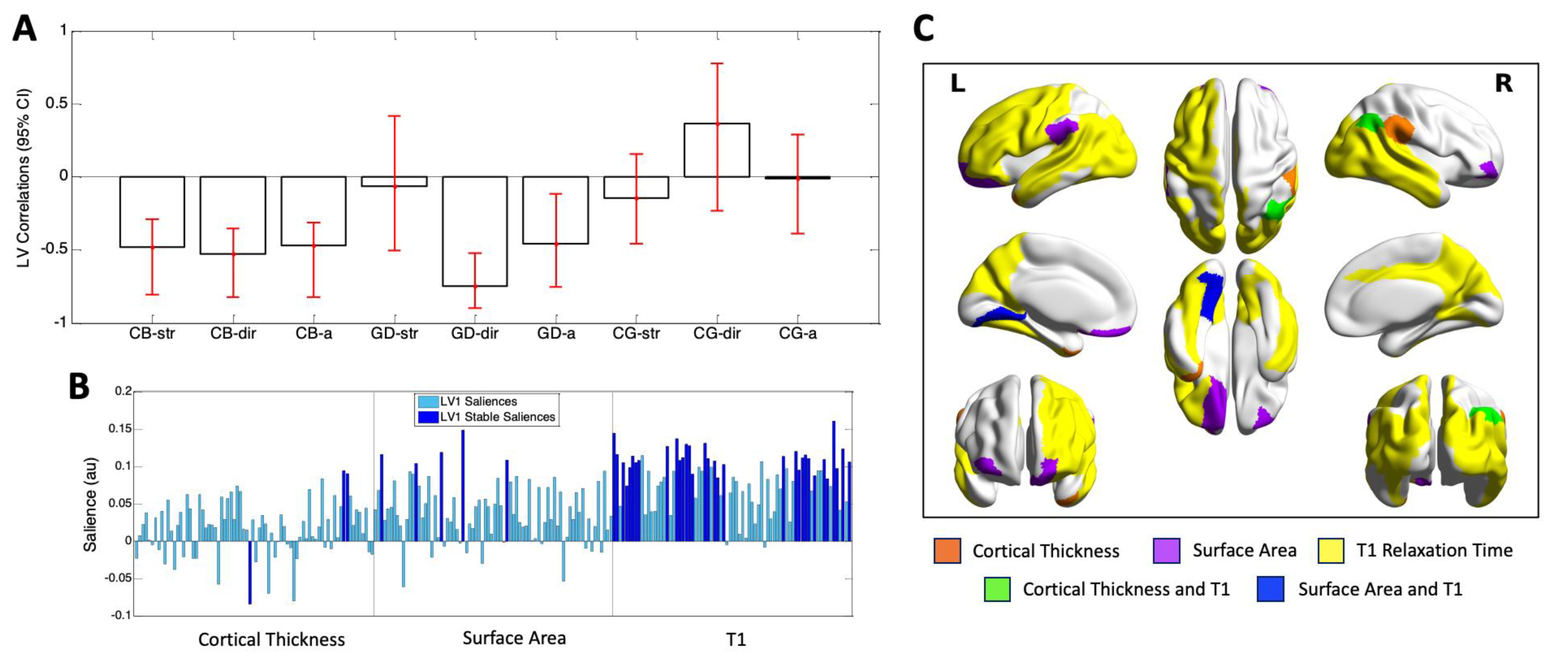
4.2. Task Partial Least Squares (PLS) Analysis
4.3. Behavior PLS Analysis
5. Discussion
Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Press: Arlington, VA, USA, 2013. [Google Scholar]
- Aitken, M.; Steensma, T.D.; Blanchard, R.; VanderLaan, D.P.; Wood, H.; Fuentes, A.; Spegg, C.; Wasserman, L.; Ames, M.; Fitzsimmons, C.L.; et al. Evidence for an altered sex ratio in clinic-referred adolescents with gender dysphoria. J. Sex. Med. 2015, 12, 756–763. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, N.M.; Giovanardi, G.; Zitz, C.; Carmichael, P. Sex ratio in children and adolescents referred to the Gender Identity Development Service in the UK (2009–2016) [Letter to the Editor]. Arch. Sex. Behav. 2018, 47, 1301–1304. [Google Scholar] [CrossRef] [PubMed]
- Meerwijk, E.L.; Sevelius, J.M. Transgender population size in the United States: A meta-regression of population-based probability samples. Am. J. Pub. Health 2017, 107, e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Hoekzema, E.; Schagen, S.E.E.; Kreukels, B.P.C.; Veltman, D.J.; Cohen-Kettenis, P.T.; Delemarre-van de Waal, H.; Bakker, J. Regional volumes and spatial volumetric distribution of gray matter in the gender dysphoric brain. Psychoneuroendocrinology 2015, 55, 59–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillamon, A.; Junque, C.; Gomez-Gil, E. A review of the status of brain structure research in transsexualism. Arch. Sex. Behav. 2016, 45, 1615–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hines, M. Neuroscience and sex/gender: Looking back and forward. J. Neurosci. 2020, 40, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Bahlburg, H.F.L. Transsexualism (“gender identity disorder”)—A CNS-limited form of intersexuality? Adv. Exp. Med. Biol. 2011, 707, 75–79. [Google Scholar] [CrossRef]
- Nguyen, H.B.; Loughead, J.; Lipner, E.; Hantsoo, L.; Kornfield, S.L.; Epperson, C.N. What has sex got to do with it? The role of hormones in the transgender brain. Neuropsychopharmacology 2018, 44, 22–37. [Google Scholar] [CrossRef]
- Ristori, J.; Cocchetti, C.; Romani, A.; Mazzoli, F.; Vignozzi, L.; Maggi, M.; Fisher, A.D. Brain sex differences related to gender identity development: Genes or hormones? Int. J. Mol. Sci. 2020, 21, 2123. [Google Scholar] [CrossRef] [Green Version]
- Roselli, C.E. Neurobiology of gender identity and sexual orientation. J. Neuroendocrinol. 2018, 30, e12562. [Google Scholar] [CrossRef]
- Hines, M. Prenatal endocrine influences on sexual orientation and on sexually differentiated childhood behavior. Front. Neuroendocrinol. 2011, 32, 170–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raznahan, A.; Shaw, P.; Lalonde, F.; Stockman, M.; Wallace, G.L.; Greenstein, D.; Clasen, L.; Gogtay, N.; Giedd, J.N. How does your cortex grow? J. Neurosci. 2011, 31, 7174–7177. [Google Scholar] [CrossRef] [PubMed]
- Wierenga, L.M.; Langen, M.; Oranje, B.; Durston, S. Unique developmental trajectories of cortical thickness and surface area. NeuroImage 2014, 87, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Wierenga, L.M.; Doucet, G.E.; Dima, D.; Agartz, I.; Aghajani, M.; Akudjedu, T.N.; Albajes-Eizagirre, A.; Alnaes, D.; Alpert, K.I.; Andreassen, O.E.; et al. Greater male than female variability in regional brain structure across the lifespan. Hum. Brain Mapp. 2020. [Google Scholar] [CrossRef] [PubMed]
- Breger, R.K.; Yetkin, Z.; Fischer, M.E.; Papke, R.A.; Haughton, V.M.; Rimm, A.A. T1 and T2 in the cerebrum: Correlation with age, gender, and demographic factors. Radiology 1991, 181, 545–547. [Google Scholar] [CrossRef]
- Cho, S.; Jones, D.; Reddick, W.E.; Ogg, R.J.; Steen, R.G. Establishing norms for age-related changes in proton T1 of human brain tissue in vivo. Magn. Reson. Imaging 1997, 15, 1133–1143. [Google Scholar] [CrossRef]
- Steen, R.G.; Ogg, R.J.; Reddick, W.E.; Kingsley, P.B. Age-related changes in the pediatric brain: Quantitative MR evidence of maturational changes during adolescence. Am. J. Neuroradiol. 1997, 18, 819–828. [Google Scholar]
- Gennatas, E.D.; Avants, B.B.; Wolf, D.H.; Satterthwaite, T.D.; Ruparel, K.; Ciric, R.; Hakonarson, H.; Gur, R.E.; Gur, R.C. Age-related effects and sex differences in gray matter density, volume, mass, and cortical thickness from childhood to young adulthood. J. Neurosci. 2017, 37, 5065–5073. [Google Scholar] [CrossRef]
- Rabinowicz, T.; Petetot, J.M.-C.; Khoury, J.C.; de Courten-Myers, G.M. Neocortical maturation during adolescence: Change in neuronal soma dimension. Brain Cogn. 2009, 69, 328–336. [Google Scholar] [CrossRef]
- Bonifacio, J.H.; Maser, C.; Stadelman, K.; Palmert, M. Management of gender dysphoria in adolescents in primary care. Can. Med. Assoc. J. 2019, 191, E69–E75. [Google Scholar] [CrossRef] [Green Version]
- Balthazart, J. Sexual partner preference in animals and humans. Neurosci. Biobehav. Rev. 2020, 115, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Bogaert, A.F.; Skorska, M.N. A short review of biological research on the development of sexual orientation. Horm. Behav. 2020, 119, 104659. [Google Scholar] [CrossRef] [PubMed]
- Ellis, L.; Ames, M.A. Neurohormonal functioning and sexual orientation: A theory of homosexuality-heterosexuality. Psychol. Bull. 1987, 101, 233–258. [Google Scholar] [CrossRef] [PubMed]
- Hines, M. Gender development and the human brain. Annu. Rev. Neurosci. 2011, 34, 69–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzouri, A.; Savic, I. Possible neurobiological underpinnings of homosexuality and gender dysphoria. Cereb. Cortex 2019, 29, 2084–2101. [Google Scholar] [CrossRef] [PubMed]
- Calzo, J.P.; Blashill, A.J. Child sexual orientation and gender identity in the Adolescent Brain Cognitive Development Cohort Study. JAMA Pediatr. 2018, 172, 1090–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herdt, G.; McClintock, M.K. The magical age of 10. Arch. Sex. Behav. 2000, 29, 587–606. [Google Scholar] [CrossRef]
- McClintock, M.K.; Herdt, G. Rethinking puberty: The development of sexual attraction. Curr. Dir. Psychol. Sci. 1996, 5, 178–183. [Google Scholar] [CrossRef]
- Bailey, J.M.; Vasey, P.L.; Diamond, L.M.; Breedlove, S.M.; Vilain, E.; Epprecht, M. Sexual orientation, controversy, and science. Psychol. Sci. Public Interest 2016, 17, 45–101. [Google Scholar] [CrossRef] [Green Version]
- Cavazos-Rehg, P.A.; Krauss, M.J.; Spitznagel, E.L.; Schootman, M.; Bucholz, K.K.; Peipert, J.F.; Sanders-Thompson, V.; Cottler, L.B.; Bierut, L.J. Age of sexual debut among US adolescents. Contraception 2009, 80, 158–162. [Google Scholar] [CrossRef] [Green Version]
- Savin-Williams, R.C.; Ream, G.L. Prevalence and stability of sexual orientation components during adolescence and young adulthood. Arch. Sex. Behav. 2007, 36, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Savin-Williams, R.C.; Joyner, K.; Rieger, G. Prevalence and stability of self-reported sexual orientation identity during young adulthood. Arch. Sex. Behav. 2012, 41, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Norton, S.; Rahman, Q. Adolescent sexual behavior patterns in a British birth cohort: A latent class analysis. Arch. Sex. Behav. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmore, J.H.; Knickmeyer, R.C.; Gao, W. Imaging structural and functional brain development in early childhood. Nat. Rev. Neurosci. 2018, 19, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Kaczkurkin, A.N.; Raznahan, A.; Satterthwaite, T.D. Sex differences in the developing brain: Insights from multimodal neuroimaging. Neuropsychopharmacol. 2019, 44, 71–85. [Google Scholar] [CrossRef]
- Lyall, A.E.; Shi, F.; Geng, X.; Woolson, S.; Li, G.; Wang, L.; Hamer, R.M.; Shen, D.; Gilmore, J.H. Dynamic development of regional cortical thickness and surface area in early childhood. Cereb. Cortex 2015, 25, 2204–2212. [Google Scholar] [CrossRef]
- Panizzon, M.S.; Fennema-Notestine, C.; Eyler, L.T.; Jernigan, T.L.; Prom-Wormley, E.; Neale, M.; Jacobson, K.; Lyons, M.J.; Grant, M.D.; Franz, C.E.; et al. Distinct genetic influences on cortical surface area and cortical thickness. Cereb. Cortex 2009, 19, 2728–2735. [Google Scholar] [CrossRef]
- Winkler, A.M.; Kochunov, P.; Blangero, J.; Almasy, L.; Zilles, K.; Fox, P.T.; Duggirala, R.; Glahn, D.C. Cortical thickness or grey matter volume? The importance of selecting the phenotype for imaging genetics studies. NeuroImage 2010, 53, 1135–1146. [Google Scholar] [CrossRef] [Green Version]
- Jha, S.C.; Xia, K.; Ahn, M.; Girault, J.B.; Li, G.; Wang, L.; Shen, D.; Zou, F.; Zhu, H.; Styner, M.; et al. Environmental influences on infant cortical thickness and surface area. Cereb. Cortex 2019, 29, 1139–1149. [Google Scholar] [CrossRef] [Green Version]
- Raznahan, A.; Greenstein, D.; Lee, N.R.; Clasen, L.S.; Giedd, J.N. Prenatal growth in humans and postnatal brain maturation into late adolescence. Proc. Natl. Acad. Sci. USA 2012, 109, 11366–11371. [Google Scholar] [CrossRef] [Green Version]
- Colby, J.B.; O’Hare, E.D.; Bramen, J.E.; Sowell, E.R. Structural brain development: Birth through adolescence. In Neural Circuit Development and Function in the Brain; Rubenstein, J.L.R., Rakic, P., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 207–230. [Google Scholar] [CrossRef]
- Deoni, S.C.L. Quantitative relaxometry of the brain. Top. Magn. Reson. Imaging 2010, 21, 101–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gracien, R.-M.; Nurnberger, L.; Hok, P.; Hof, S.-M.; Reitz, S.C.; Rub, U.; Steinmetz, H.; Hilker-Roggendorf, R.; Klein, J.C.; Deichmann, R.; et al. Evaluation of brain ageing: A quantitative longitudinal MRI study over 7 years. Eur. Radiol. 2017, 27, 1568–1576. [Google Scholar] [CrossRef] [PubMed]
- Saito, N.; Sakai, O.; Ozonoff, A.; Jara, H. Relaxo-volumetric multispectral quantitative magnetic resonance imaging of the brain over the human lifespan: Global and regional aging patterns. Magn. Reson. Imaging 2009, 27, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Seiler, A.; Schongrundner, S.; Stock, B.; Noth, U.; Hattingen, E.; Steinmetz, H.; Klein, J.C.; Baudrexel, S.; Wagner, M.; Deichmann, R.; et al. Cortical aging—New insights with multiparametric quantitative MRI. Aging 2020, 12, 16195–16210. [Google Scholar] [CrossRef] [PubMed]
- Skorska, M.N.; Chavez, S.; Devenyi, G.A.; Patel, R.; Thurston, L.T.; Lai, M.-C.; Zucker, K.J.; Chakravarty, M.M.; Lobaugh, N.J.; VanderLaan, D.P. Data from: A multi-modal MRI analysis of cortical structure in relation to gender dysphoria, sexual orientation, and age in adolescents. Sch. Portal Dataverse 2020. [Google Scholar] [CrossRef]
- Deogracias, J.J.; Johnson, L.L.; Meyer-Bahlburg, H.F.L.; Kessler, S.J.; Schober, J.M.; Zucker, K.J. The Gender Identity/Gender Dysphoria Questionnaire for Adolescents and Adults. J. Sex. Res. 2007, 44, 370–379. [Google Scholar] [CrossRef]
- Singh, D.; Deogracias, J.J.; Johnson, L.J.; Bradley, S.J.; Kibblewhite, S.J.; Owen-Anderson, A.; Badali-Peterson, M.; Meyer-Bahlburg, H.F.L.; Zucker, K.J. The Gender Identity/Gender Dysphoria Questionnaire for Adolescents and Adults: Further validity evidence. J. Sex. Res. 2010, 47, 49–58. [Google Scholar] [CrossRef]
- Zucker, K.J.; Bradley, S.J.; Owen-Anderson, A.; Kibblewhite, S.J.; Wood, H.; Singh, D.; Choi, K. Demographics, behavior problems, and psychosexual characteristics of adolescents with gender identity disorder or transvestic fetishism. J. Sex. Marital Ther. 2012, 38, 151–189. [Google Scholar] [CrossRef]
- Drummond, K.D.; Bradley, S.J.; Badali-Peterson, M.; Zucker, K.J. A follow-up study of girls with gender identity disorder. Dev. Psychol. 2008, 44, 34–45. [Google Scholar] [CrossRef]
- Storms, M.D. Theories of sexual orientation. J. Pers. Soc. Psychol. 1980, 38, 783–792. [Google Scholar] [CrossRef]
- Zucker, K.J.; Bradley, S.J.; Oliver, G.; Blake, J.; Fleming, S.; Hood, J. Psychosexual development of women with congenital adrenal hyperplasia. Horm. Behav. 1996, 30, 300–318. [Google Scholar] [CrossRef] [PubMed]
- Deoni, S.C.L. High-resolution T1 mapping of the brain at 3T with driven equilibrium single pulse observation of T1 with high-speed incorporation of RF field inhomogeneities (DESPOT1-HIFI). J. Magn. Reson. Imaging 2007, 26, 1106–1111. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qiu, M.; Kim, H.; Constable, R.T. T1 Measurements incorporating flip angle calibration and correction in vivo. J. Mag. Reson. 2006, 182, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Chavez, S.; Stanisz, G.J. A novel method for simultaneous 3D B1 and T1 mapping: The method of slopes (MoS). NMR Biomed. 2012, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Loken, C.; Gruner, D.; Groer, L.; Peltier, R.; Bunn, N.; Craig, M.; Henriques, T.; Dempsey, J.; Yu, C.-H.; Chen, J.; et al. SciNet: Lessons learned from building a power-efficient top-20 system and data centre. J. Phys. Conf. Ser. 2010, 256, 012026. [Google Scholar] [CrossRef]
- Ponce, M.; van Zon, R.; Northrup, S.; Gruner, D.; Chen, J.; Ertinaz, F.; Fedoseev, A.; Groer, L.; Mao, F.; Mundim, B.C.; et al. Deploying a top-100 supercomputer for large parallel workloads: The Niagara Supercomputer. In Proceedings of the Practice and Experience in Advanced Research Computing (PEARC 2019), Chicago, IL, USA, 28 July–1 August 2019. [Google Scholar] [CrossRef] [Green Version]
- Tustison, N.J.; Avants, B.B.; Cook, P.A.; Zheng, Y.; Egan, A.; Yushkevich, P.A.; Gee, J.C. N4ITK: Improved N3 bias correction. IEEE Trans. Med. Imaging 2010, 29, 1310–1320. [Google Scholar] [CrossRef] [Green Version]
- Eskildsen, S.F.; Coupe, P.; Fonov, V.; Manjon, J.V.; Leung, K.K.; Guizard, N.; Wassef, S.H.; Ostergaard, L.R.; Collins, D.L. The Alzheimer’s Neuroimaging Initiative. BEaST: Brain extraction based on nonlocal segmentation technique. NeuroImage 2012, 59, 2362–2373. [Google Scholar] [CrossRef]
- Bedford, S.A.; Park, M.T.M.; Devenyi, G.A.; Tullo, S.; Germann, J.; Patel, R.; Anagnostou, E.; Baron-Cohen, S.; Bullmore, E.T.; Chura, L.R.; et al. Large-scale analyses of the relationship between sex, age and intelligence quotient heterogeneity and cortical morphometry in autism spectrum disorder. Mol. Psychiatry 2020, 25, 614–628. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Patel, R.; Park, M.T.M.; Masellis, M.; Knight, J.; Chakravarty, M.M. Heritability estimates of cortical anatomy: The influence and reliability of different estimation strategies. NeuroImage 2018, 178, 78–91. [Google Scholar] [CrossRef] [Green Version]
- Ad-Dab’bagh, Y.; Einarson, D.; Lyttelton, O.; Muehlboeck, J.-S.; Mok, K.; Ivanov, O.; Vincent, R.D.; Lepage, C.; Lerch, J.; Fombonne, E.; et al. The CIVET image-processing environment: A fully automated comprehensive pipeline for anatomical neuroimaging research. In Proceedings of the 12th Annual Meeting of the Organization for Human Brain Mapping, Florence, Italy, 11–15 June 2006. [Google Scholar]
- Zijdenbos, A.P.; Forghani, R.; Evans, A.C. Automatic “pipeline” analysis of 3-D MRI data for clinical trials: Application to multiple sclerosis. IEEE Trans. Med. Imaging 2002, 21, 1280–1291. [Google Scholar] [CrossRef]
- Lerch, J.P.; Evans, A.C. Cortical thickness analysis examined through power analysis and a population simulation. NeuroImage 2005, 24, 163–173. [Google Scholar] [CrossRef]
- Lyttelton, O.; Boucher, M.; Robbins, S.; Evans, A.C. An unbiased iterative group registration template for cortical surface analysis. NeuroImage 2007, 34, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage 2002, 15, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Chavez, S. Calibrating variable flip angle (VFA)-based T1 maps: When and why a simple scaling factor is justified. In Proceedings of the International Society of Magnetic Resonance in Medicine (ISMRM), Paris, France, 16–21 June 2018. [Google Scholar]
- Avants, B.B.; Tustison, N.J.; Song, G.; Cook, P.A.; Klein, A.; Gee, J.C. A reproducible evaluation of ANTs similarity metric performance in brain image registration. NeuroImage 2011, 54, 2033–2044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tustison, N.J.; Avants, B.B. Explicit B-spline regularization in diffeomorphic image registration. Front. Neuroinform. 2013, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MATLAB, Version 9.3.0 (R2017b); The MathWorks Inc.: Natick, MA, USA, 2017.
- Wechsler, D. Wechsler Intelligence Scale for Children, 3rd ed.; The Psychological Corporation: San Antonio, TX, USA, 1991. [Google Scholar]
- Wechsler, D. Wechsler Adult Intelligence Scale, 4th ed.; Pearson: San Antonio, TX, USA, 2008. [Google Scholar]
- Wechsler, D. Wechsler Intelligence Scale for Children, 5th ed.; The Psychological Corporation: San Antonio, TX, USA, 2014. [Google Scholar]
- Petersen, A.C.; Crockett, L.; Richards, M.; Boxer, A. A self-report measure of pubertal status: Reliability, validity, and initial norms. J. Youth Adolesc. 1988, 17, 117–133. [Google Scholar] [CrossRef]
- Achenbach, T.M. Manual for the Youth Self-Report and 1991 Profile; University of Vermont Department of Psychiatry: Burlington, VT, USA, 1991. [Google Scholar]
- Achenbach, T.M.; Edelbrock, C. Manual for the Youth Self-Report and Profile; University of Vermont Department of Psychiatry: Burlington, VT, USA, 1986. [Google Scholar]
- Achenbach, T.M.; Rescorla, L.A. Manual for the ASEBA School-Age Forms & Profiles; University of Vermont, Research Center for Children, Youth, & Families: Burlington, VT, USA, 2001. [Google Scholar]
- Ruigrok, A.N.V.; Salimi-Khorshidi, G.; Lai, M.-C.; Baron-Cohen, S.; Lombardo, M.V.; Tait, R.J.; Suckling, J. A meta-analysis of sex differences in human brain structure. Neurosci. Biobehav. Rev. 2014, 39, 34–50. [Google Scholar] [CrossRef] [Green Version]
- Ducharme, S.; Albaugh, M.D.; Nguyen, T.-V.; Hudziak, J.J.; Mateos-Perez, J.M.; Labbe, A.; Evans, A.C.; Karama, S. The Brain Development Cooperative Group. Trajectories of cortical thickness maturation in normal brain development—The importance of quality control procedures. NeuroImage 2016, 125, 267–279. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, A.R.; Bookstein, F.L.; Haxby, V.; Grady, C.L. Spatial pattern analysis of functional brain images using partial least squares. NeuroImage 1996, 3, 143–157. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, A.R.; Lobaugh, N.J. Partial least squares analysis of neuroimaging data: Applications and advances. NeuroImage 2004, 23, S250–S263. [Google Scholar] [CrossRef]
- Chen, X.J.; Kovacevic, N.; Lobaugh, N.J.; Sled, J.G.; Henkelman, R.M.; Henderson, J.T. Neuroanatomical differences between mouse strains as shown by high-resolution 3D MRI. NeuroImage 2005, 29, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Geladi, P.; Kowalski, B.R. Partial Least-Squares Regression: A Tutorial. Anal. Chim. Acta 1986, 185, 1–17. [Google Scholar] [CrossRef]
- MATLAB, Version 8.3.0 (R2014a); The MathWorks Inc.: Natick, MA, USA, 2014.
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A network visualization tool for human brain connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakic, P. Specification of cerebral cortical areas. Science 1988, 241, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Rakic, P. The radial edifice of cortical architecture: From neuronal silhouettes to genetic engineering. Brain Res. Rev. 2007, 55, 204–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modabbernia, A.; Reichenberg, A.; Ing, A.; Moser, D.A.; Doucet, G.E.; Artiges, E.; Banaschewski, T.; Barker, G.J.; Becker, A.; Bokde, A.L.W.; et al. Linked patterns of biological and environmental covariation with brain structure in adolescence: A population-based longitudinal study. Mol. Psychiatry 2020. [Google Scholar] [CrossRef]
- Schulz, K.M.; Molenda-Figueira, H.A.; Sisk, C.L. Back to the future: The organizational-activational hypothesis adapted to puberty and adolescence. Horm. Behav. 2009, 55, 597–604. [Google Scholar] [CrossRef] [Green Version]
- Herting, M.M.; Sowell, E.R. Puberty and structural brain development in humans. Front. Neuroendocrinol. 2017, 44, 122–137. [Google Scholar] [CrossRef] [Green Version]
- Berenbaum, S.A.; Beltz, A.M.; Corely, R. The importance of puberty for adolescent development: Conceptualization and measurement. Adv. Child. Dev. Behav. 2015, 48, 53–59. [Google Scholar] [CrossRef]
- Cafiero, R.; Brauer, J.; Anwander, A.; Friederici, A.D. The concurrence of cortical surface area expansion and white matter myelination in human brain development. Cereb. Cortex 2019, 29, 827–837. [Google Scholar] [CrossRef]
- Breedlove, S.M. Prenatal influences on human sexual orientation: Expectations versus data. Arch. Sex. Behav. 2017, 46, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Stuber, C.; Morawski, M.; Schafer, A.; Labadie, C.; Wahnert, M.; Leuze, C.; Streigher, M.; Barapatre, N.; Reimann, K.; Geyer, S.; et al. Myelin and iron concentration in the human brain: A quantitative study of MRI contrast. NeuroImage 2014, 93, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Juraska, J.M.; Markham, J.A. The cellular basis for volume changes in the rat cortex during puberty: White and gray matter. Ann. N. Y. Acad. Sci. 2004, 1021, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Abi Ghanem, C.; Degerny, C.; Hussain, R.; Liere, P.; Pianos, A.; Tourpin, S.; Habert, R.; Macklin, W.B.; Schumacher, M.; Ghoumari, A.M. Long-lasting masculinizing effects of postnatal androgens on myelin governed by the brain androgen receptor. PLoS Genet. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Button, K.S.; Ioannidis, J.P.A.; Mokrysz, C.; Nosek, B.A.; Flint, J.; Robinson, E.S.J.; Munafo, M.R. Power failure: Why small sample size undermines the reliability of neuroscience. Nat. Rev. Neurosci. 2013, 14, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akkermans, S.E.A.; van Rooij, D.; Rommelse, N.; Hartman, C.A.; Hoekstra, P.J.; Franke, B.; Mennes, M.; Buitelaar, J.K. Effect of tobacco smoking on frontal cortical thickness development: A longitudinal study in a mixed cohort of ADHD-affected and -unaffected youth. Eur. Neuropsychopharmacol. 2017, 27, 1022–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Q.; Peng, W.; Zhang, J.; Weng, X.; Zhang, Y.; Liu, T.; Zang, Y.-F.; Wang, Z. Short-term apparent brain tissue changes are contributed by cerebral blood flow alterations. PLoS ONE 2017, 12, e0182182. [Google Scholar] [CrossRef] [Green Version]
- Tardif, C.L.; Steele, C.J.; Lampe, L.; Bazin, P.-L.; Ragert, P.; Villringer, A.; Gauthier, C.J. Investigation of the confounding effects of vasculature and metabolism on computational anatomy studies. NeuroImage 2017, 149, 233–243. [Google Scholar] [CrossRef]
- Baldinger-Melich, P.; Castro, M.F.U.; Seiger, R.; Ruef, A.; Dwyer, D.B.; Kranz, G.S.; Klobl, M.; Kambeitz, J.; Kaufmann, U.; Windischberger, C.; et al. Sex matters: A multivariate pattern analysis of sex- and gender-related neuroanatomical differences in cis- and transgender individuals using structural magnetic resonance imaging. Cereb. Cortex 2019, 30, 1345–1356. [Google Scholar] [CrossRef]
- Burke, S.M.; Manzouri, A.H.; Dhejne, C.; Bergstrom, K.; Arver, S.; Feusner, J.D.; Savic-Berglund, I. Testosterone effects on the brain in transgender men. Cereb. Cortex 2018, 28, 1582–1596. [Google Scholar] [CrossRef]
- Flint, C.; Forster, K.; Koser, S.A.; Konrad, C.; Zwitserlood, P.; Berger, K.; Hermesdorf, M.; Kircher, T.; Nenadic, I.; Krug, A.; et al. Biological sex classification with structural MRI data shows increased misclassification in transgender women. Neuropsychopharmacology 2020, 45, 1758–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilpatrick, L.A.; Holmberg, M.; Manzouri, A.; Savic, I. Cross sex hormone treatment is linked with a reversal of cerebral patterns associated with gender dysphoria to the baseline of cisgender controls. Eur. J. Neurosci. 2019, 50, 3269–3281. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.-W.; Kim, Y.-H.; Park, K.; Jeong, G.-W. A comparative study of white matter volume between postoperative female-to-male transsexuals and healthy female. Int. J. Impot. Res. 2019, 31, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.A.; Spritzer, P.M.; Suh, J.S.; Minuzzi, L.; Frey, B.N.; Schwarz, K.; Costa, A.B.; da Silva, D.C.; Garcia, C.C.G.; Fontanari, A.M.V.; et al. The link between estradiol and neuroplasticity in transgender women after gender-affirming surgery: A bimodal hypothesis. Neuroendocrinology 2020, 110, 489–500. [Google Scholar] [CrossRef]
- Starcevic, A.; Dakovic, M.; Radojicic, Z.; Filipovic, B. A structural magnetic resonance imaging study in therapy naïve transsexual individuals. Folia Morphol. 2020. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cisgender Boys | GD AFAB | Cisgender Girls | F (df) | p | |
|---|---|---|---|---|---|
| n | 14 | 16 | 17 | ||
| Age (months) | |||||
| M | 184.93 | 193.38 | 191.71 | 0.75 (2, 44) | 0.480 |
| SD | 25.61 | 14.20 | 19.00 | ||
| Range | 147–216 | 162–216 | 152–214 | ||
| GIDYQ-AA | |||||
| M | 4.91 | 2.21 | 4.90 | 693.34 (2, 44) c | <0.001 |
| SD | 0.12 | 0.35 | 0.15 | ||
| Range (1–5) a | 4.63–5.00 | 1.74–3.04 | 4.48–5.00 | ||
| Strength of attractions | |||||
| M | 3.25 | 3.00 | 2.83 | 0.54 (2, 44) d | 0.587 |
| SD | 1.28 | 1.30 | 0.76 | ||
| Range (1.41–7.07) b | 1.70–5.37 | 1.41–6.05 | 1.41–4.44 | ||
| Degree of androphilia-gynephilia | |||||
| M | 64.60 | 46.38 | 30.94 | 25.93 (2, 44) | <0.001 |
| SD | 9.29 | 15.23 | 13.18 | ||
| Range (11–79) b | 43.96–75.55 | 17.35–75.55 | 14.04–68.20 | ||
| Task PLS | Behavior PLS | |||
|---|---|---|---|---|
| SA/CT | Hem. | ROI | SA/CT/T1 | Hem. |
| Frontal Lobe | ||||
| CT | Left | Superior Frontal Gyrus: Orbital Part | SA | Left |
| CT | Right | Superior Frontal Gyrus: Orbital Part | ||
| SA | Left | Middle Frontal Gyrus | T1 | Left |
| CT | Left | Rolandic Operculum | T1 | Left |
| SA | Left | Superior Frontal Gyrus: Medial | ||
| SA | Left | Precentral Gyrus | T1 | Left |
| SA | Right | Precentral Gyrus | ||
| CT | Right | Gyrus Rectus | ||
| Gyrus Rectus | SA | Left | ||
| SA | Left | Supplementary Motor Area | ||
| Middle Frontal Gyrus: Orbital Part | T1 | Left | ||
| Middle Frontal Gyrus: Orbital Part | SA | Right | ||
| Superior Frontal Gyrus: Dorsolateral | T1 | Left | ||
| Inferior Frontal Gyrus: Opercular Part | T1 | Left | ||
| Inferior Frontal Gyrus: Triangular Part | T1 | Left | ||
| Inferior Frontal Gyrus: Orbital Part | T1 | Left | ||
| Parietal Lobe | ||||
| SA | Left | Angular Gyrus | T1 | Left |
| Angular Gyrus | CT; T1 | Right | ||
| Supramarginal Gyrus | SA | Left | ||
| Supramarginal Gyrus | CT | Right | ||
| Postcentral Gyrus | T1 | Left | ||
| Precuneus | T1 | Left | ||
| Precuneus | T1 | Right | ||
| Temporal Lobe | ||||
| CT | Left | Parahippocampal Gyrus | ||
| SA | Left | Temporal Pole: Superior Temporal Gyrus | ||
| SA | Left | Superior Temporal Gyrus | T1 | Left |
| Superior Temporal Gyrus | T1 | Right | ||
| SA | Left | Middle Temporal Gyrus | T1 | Left |
| Middle Temporal Gyrus | T1 | Right | ||
| SA | Right | Inferior Temporal Gyrus | T1 | Right |
| Temporal Pole: Middle Temporal Gyrus | CT | Left | ||
| Fusiform Gyrus | T1 | Left | ||
| Heschl Gyrus | T1 | Left | ||
| Heschl Gyrus | T1 | Right | ||
| Occipital Lobe | ||||
| Middle Occipital Gyrus | T1 | Left | ||
| SA | Right | Middle Occipital Gyrus | T1 | Right |
| Inferior Occipital Gyrus | T1 | Left | ||
| SA | Right | Inferior Occipital Gyrus | T1 | Right |
| SA | Left | Lingual Gyrus | SA; T1 | Left |
| Lingual Gyrus | T1 | Right | ||
| SA | Left | Calcarine Fissure and Surrounding Cortex | ||
| SA | Left | Cuneus | T1 | Left |
| Cuneus | T1 | Right | ||
| Superior Occipital Gyrus | T1 | Left | ||
| Superior Occipital Gyrus | T1 | Left | ||
| Insula and Cingulate Gyri | ||||
| SA | Left | Insula | ||
| SA | Right | Insula | ||
| SA | Right | Anterior Cingulate and Paracingulate Gyri | ||
| SA | Left | Posterior Cingulate Gyrus | ||
| SA | Right | Posterior Cingulate Gyrus | T1 | Right |
| Median Cingulate and Paracingulate Gyri | T1 | Right | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skorska, M.N.; Chavez, S.; Devenyi, G.A.; Patel, R.; Thurston, L.T.; Lai, M.-C.; Zucker, K.J.; Chakravarty, M.M.; Lobaugh, N.J.; VanderLaan, D.P. A Multi-Modal MRI Analysis of Cortical Structure in Relation to Gender Dysphoria, Sexual Orientation, and Age in Adolescents. J. Clin. Med. 2021, 10, 345. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10020345
Skorska MN, Chavez S, Devenyi GA, Patel R, Thurston LT, Lai M-C, Zucker KJ, Chakravarty MM, Lobaugh NJ, VanderLaan DP. A Multi-Modal MRI Analysis of Cortical Structure in Relation to Gender Dysphoria, Sexual Orientation, and Age in Adolescents. Journal of Clinical Medicine. 2021; 10(2):345. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10020345
Chicago/Turabian StyleSkorska, Malvina N., Sofia Chavez, Gabriel A. Devenyi, Raihaan Patel, Lindsey T. Thurston, Meng-Chuan Lai, Kenneth J. Zucker, M. Mallar Chakravarty, Nancy J. Lobaugh, and Doug P. VanderLaan. 2021. "A Multi-Modal MRI Analysis of Cortical Structure in Relation to Gender Dysphoria, Sexual Orientation, and Age in Adolescents" Journal of Clinical Medicine 10, no. 2: 345. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10020345