
Unraveling the Signaling Secretome of Platelet-Rich Plasma: Towards a Better Understanding of Its Therapeutic Potential in Knee Osteoarthritis


Abstract
:1. Introduction
2. Materials and Methods
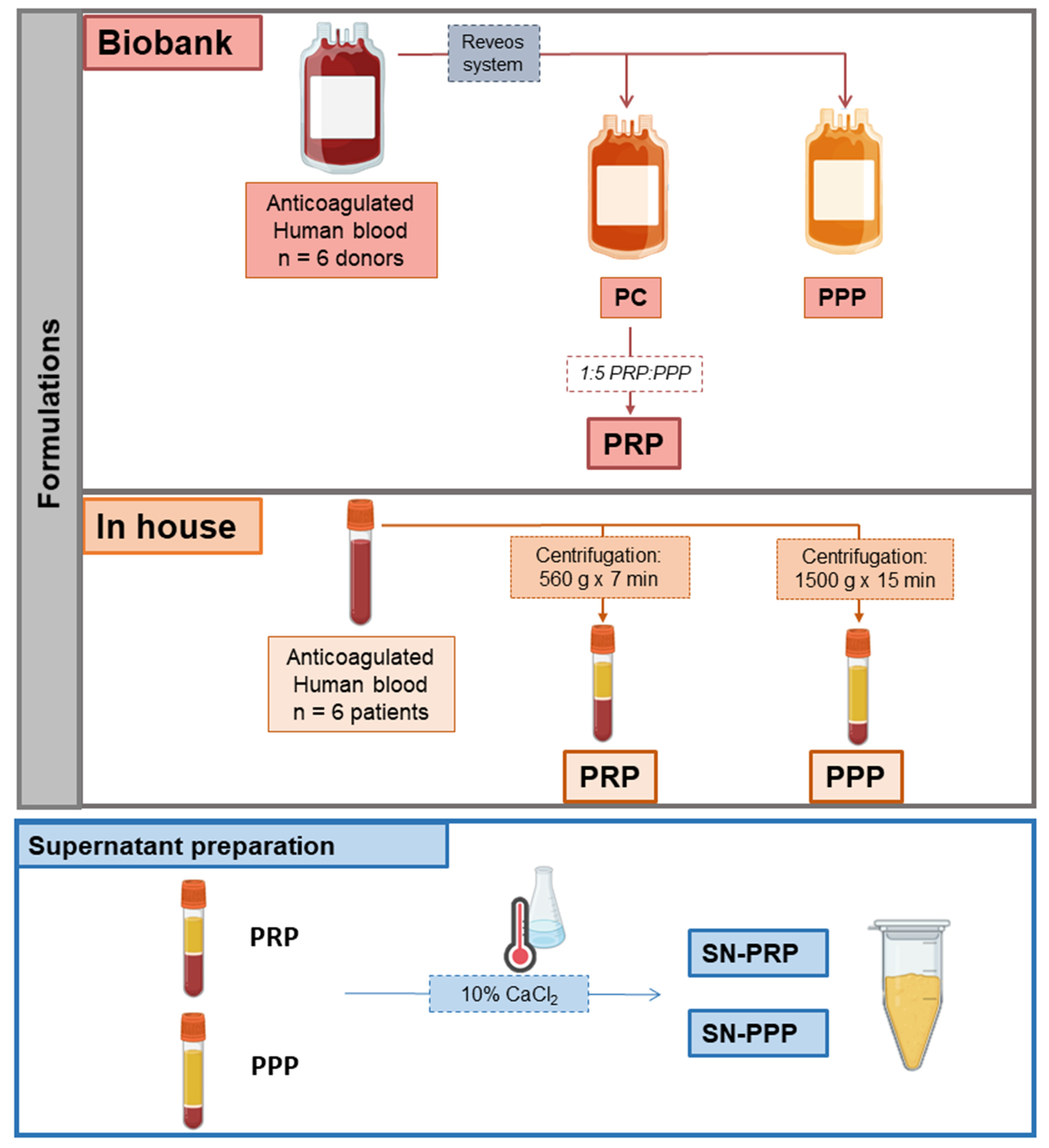
2.1. Biobank PRP and PPP
2.2. In-House PRP and PPP
2.3. Supernatants (Releasates) from PRP and PPP: SN-PRP and SN-PPP (in-House)
2.4. Molecular Characterization
2.5. ELISAs
2.6. Antibody-Based Protein Arrays
2.7. Bioinformatic Analyses
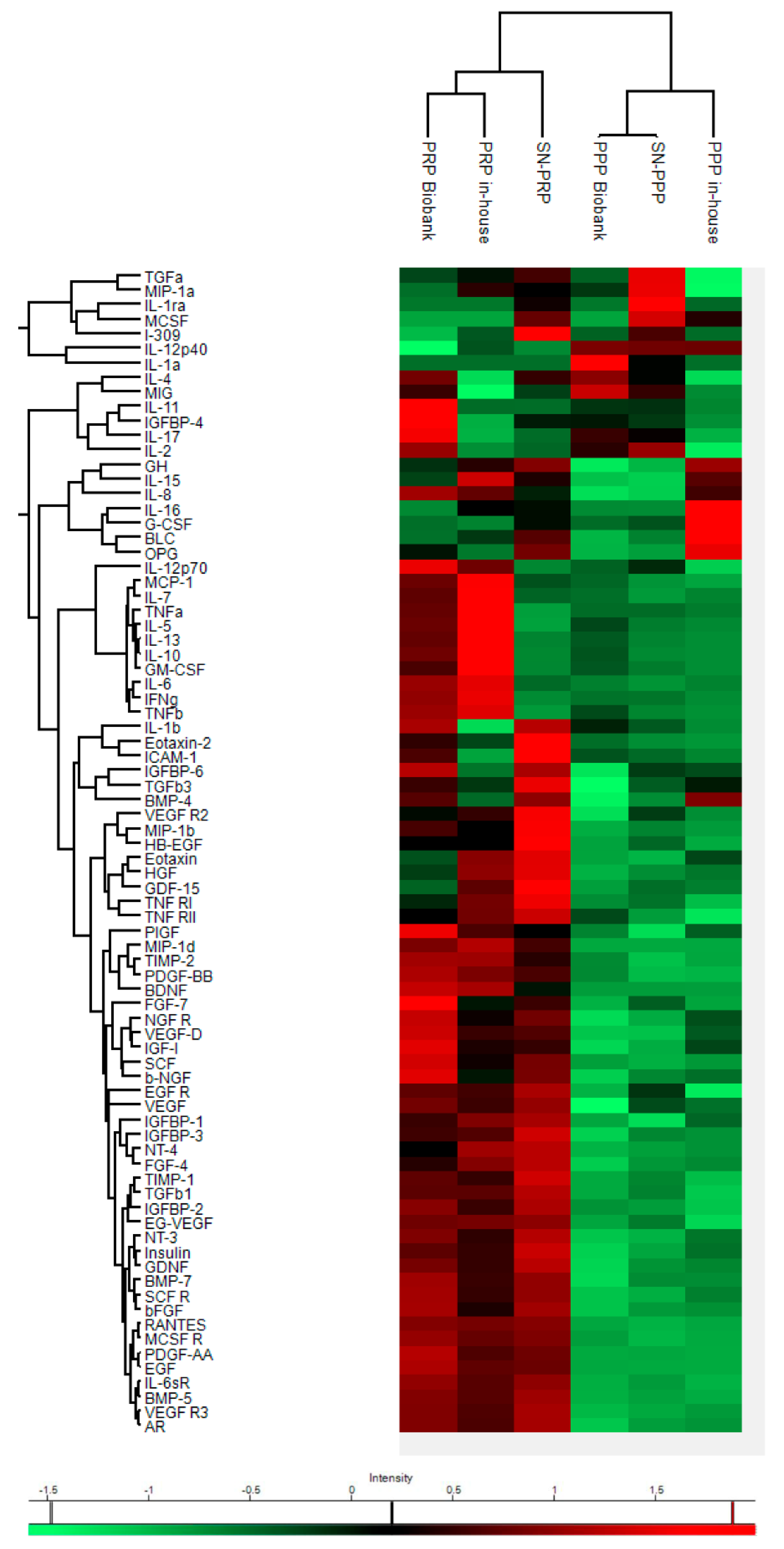
2.7.1. Clustering Analysis
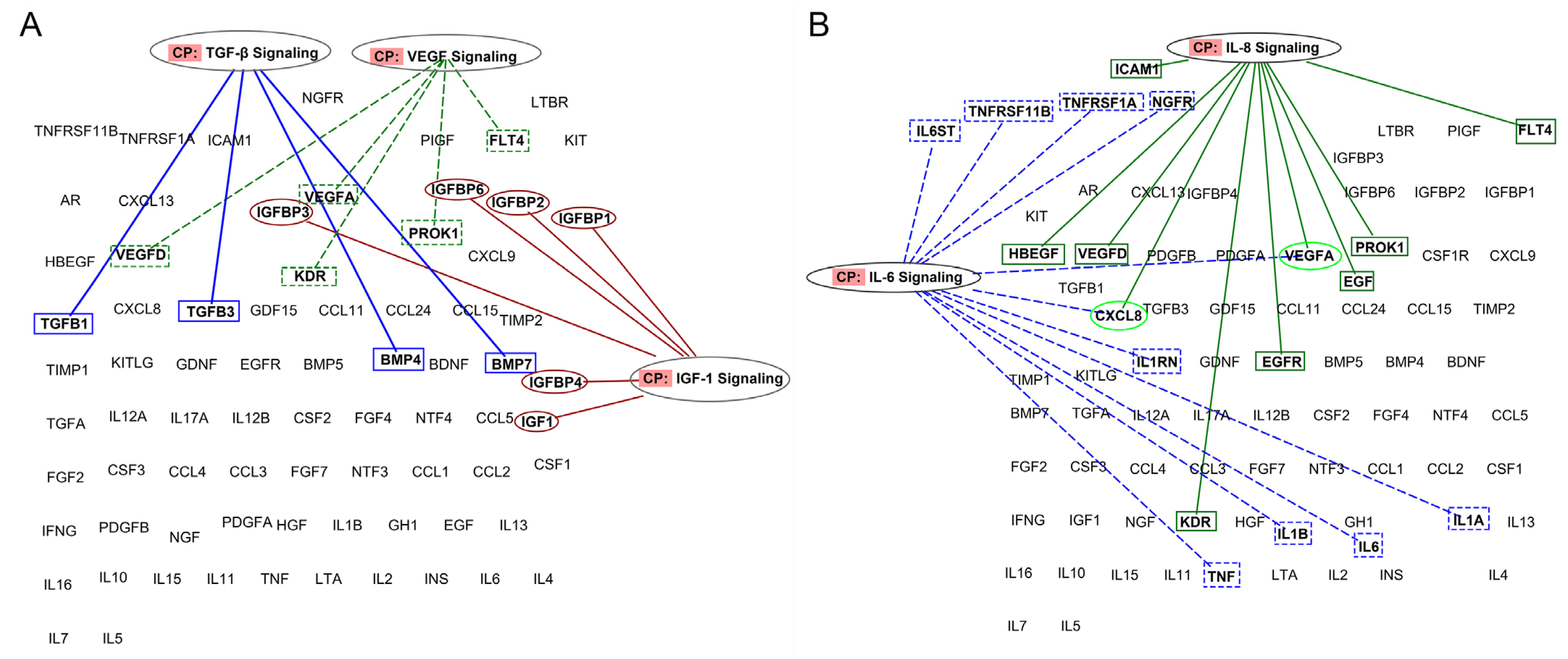
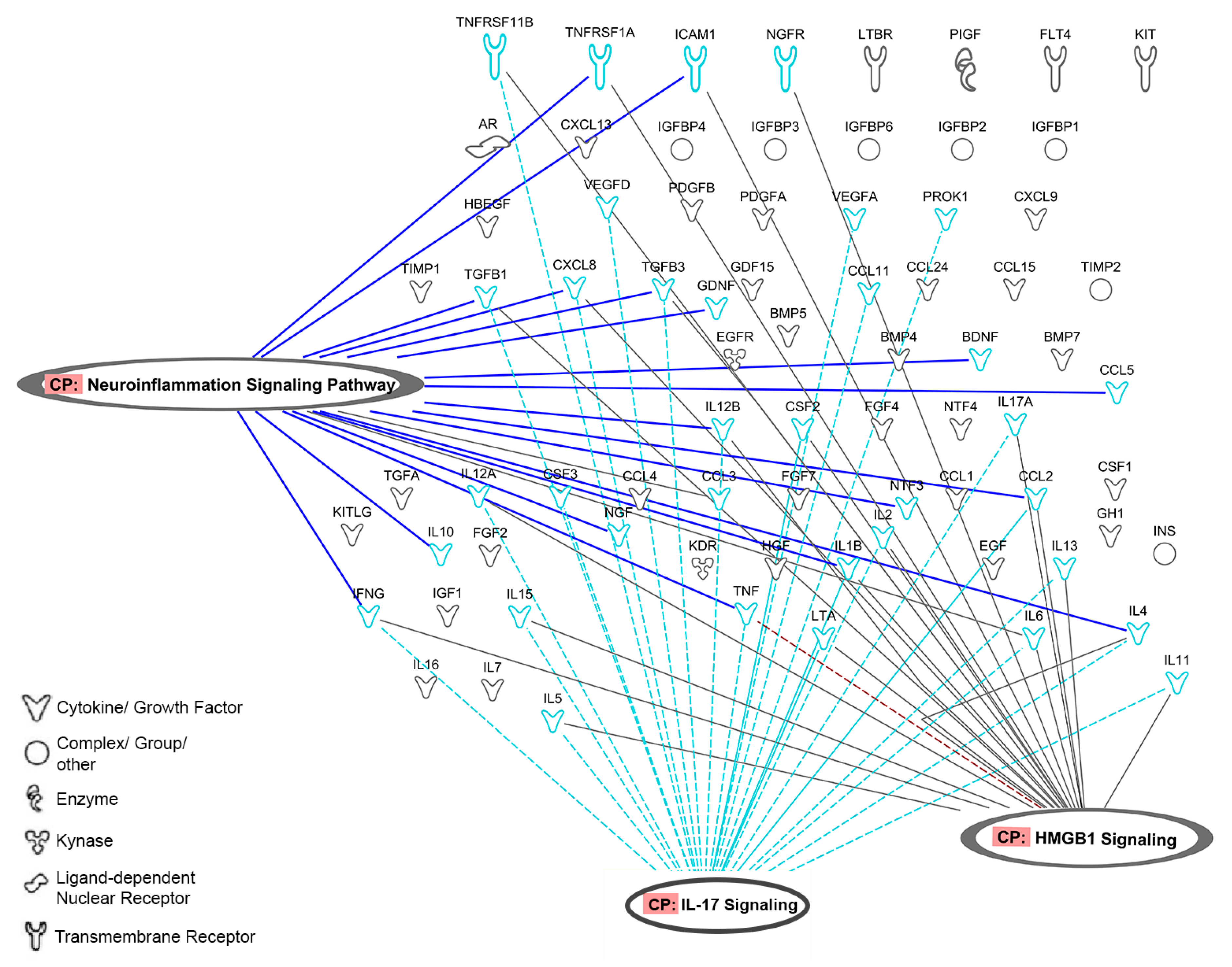
2.7.2. Ingenuity Pathway Analysis (IPA)
3. Results
3.1. Plasma Formulations/Configurations
3.2. Multiplexing Protein Arrays
3.2.1. Human Cytokine Array
3.2.2. Clustering Analyses
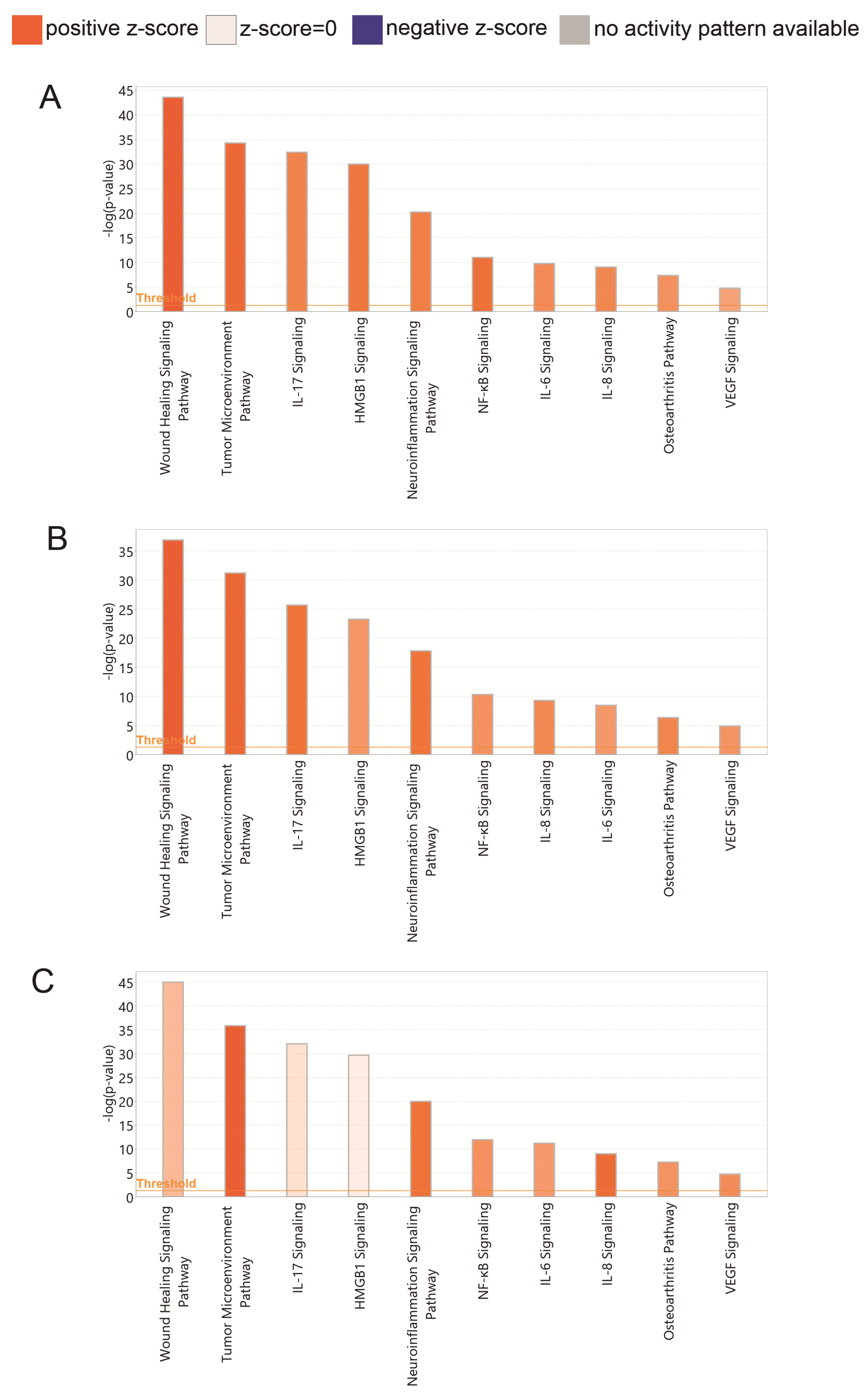
3.2.3. PRP Contains Signaling Systems (Networks) Involved in Immune Cell Interactions and Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wallace, I.J.; Worthingtonb, S.; Felsonc, D.T.; Jurmaind, R.D.; Wrene, K.T.; Maijanenf, H.; Woodsg, R.J.; Liebermana, D.E. Knee osteoarthritis has doubled in prevalence since the mid-20th century. Proc. Natl. Acad. Sci. USA 2017, 114, 9332–9336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross, M.; Smith, E.; Hoy, D.; Nolte, S.; Ackerman, I.; Fransen, M.; Bridgett, L.; Williams, S.; Guillemin, F.; Hill, C.L.; et al. The global burden of hip and knee osteoarthritis: Estimates from the global burden of disease 2010 study. Ann. Rheum. Dis. 2014, 73, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Ghebreyesus, T.A. It takes knowledge to transform the world to be a better place to grow older. Nat. Aging 2021, 1, 865. [Google Scholar] [CrossRef]
- Bull, F.C.; Al-Ansari, S.S.; Biddle, S.; Borodulin, K.; Buman, M.P.; Cardon, G.; Carty, C.; Chaput, J.P.; Chastin, S.; Chou, R.; et al. World Health Organization 2020 guidelines on physical activity and sedentary behaviour. Br. J. Sports Med. 2020, 54, 1451–1462. [Google Scholar] [CrossRef] [PubMed]
- Fry, R. The Pace of Boomer Retirements Has Accelerated in the Past Year. Pew Research Center. Available online: https://www.pewresearch.org/fact-tank/2020/11/09/the-pace-of-boomer-retirements-has-accelerated-in-the-past-year/ (accessed on 8 November 2021).
- Inacio, M.C.S.; Paxton, E.W.; Graves, S.E.; Namba, R.S.; Nemes, S. Projected increase in total knee arthroplasty in the United States—An alternative projection model. Osteoarthr. Cartil. 2017, 25, 1797–1803. [Google Scholar] [CrossRef] [Green Version]
- Losina, E.; Paltiel, A.D.; Weinstein, A.M.; Yelin, E.; Hunter, D.J.; Chen, S.P.; Klara, K.; Suter, L.G.; Solomon, D.H.; Burbine, S.A.; et al. Lifetime medical costs of knee osteoarthritis management in the United States: Impact of extending indications for total knee arthroplasty. Arthritis Care Res. 2015, 67, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Bannuru, R.; Osani, M.; Vaysbrot, E.; Arden, N.; Bennell, K.; Bierma-Zeinstra, S.; Kraus, V.; Lohmander, L.; Abbott, J.; Bhandari, M.; et al. OARSI guidelines for the non-surgical management of knee, hip, and polyarticular osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1578–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belk, J.W.; Kraeutler, M.J.; Houck, D.A.; Goodrich, J.A.; Dragoo, J.L.; McCarty, E.C. Platelet-Rich Plasma Versus Hyaluronic Acid for Knee Osteoarthritis: A Systematic Review and Meta-analysis of Randomized Controlled Trials. Am. J. Sports Med. 2021, 49, 249–260. [Google Scholar] [CrossRef]
- Migliorini, F.; Driessen, A.; Quack, V.; Sippel, N.; Cooper, B.; Mansy, Y.E.; Tingart, M.; Eschweiler, J. Comparison between intra-articular infiltrations of placebo, steroids, hyaluronic and PRP for knee osteoarthritis: A Bayesian network meta-analysis. Arch. Orthop. Trauma Surg. 2021, 141, 1473–1490. [Google Scholar] [CrossRef]
- Nurden, A.T. Platelets, inflammation and tissue regeneration. Thromb. Haemost. 2011, 105, 13–33. [Google Scholar] [CrossRef]
- Olmos Calvo, I.; Fodor, E.; Kardos, D.; Hornyák, I.; Hinsenkamp, A.; Kuten-Pella, O.; Gyevnár, Z.; Erdélyi, G.; Bárdos, T.; Paukovits, T.M.; et al. A Pilot Clinical Study of Hyperacute Serum Treatment in Osteoarthritic Knee Joint: Cytokine Changes and Clinical Effects. Curr. Issues Mol. Biol. 2021, 43, 46. [Google Scholar] [CrossRef] [PubMed]
- Mojica, E.S.; Markus, D.H.; Hurley, E.T.; Blaeser, A.M.; Jazrawi, L.M.; Campbell, K.A.; Strauss, E.J. Estimated Time to Maximum Medical Improvement of Intra-articular Injections in the Treatment of Knee Osteoarthritis—A Systematic Review. Arthroscopy 2021. [Google Scholar] [CrossRef] [PubMed]
- Van Spil, W.E.; Bierma-Zeinstra, S.M.A.; Deveza, L.A.; Arden, N.K.; Bay-Jensen, A.C.; Kraus, V.B.; Carlesso, L.; Christensen, R.; Van Der Esch, M.; Kent, P.; et al. A consensus-based framework for conducting and reporting osteoarthritis phenotype research. Arthritis Res. Ther. 2020, 22, 54. [Google Scholar] [CrossRef] [Green Version]
- Harrison, P. The use of platelets in regenerative medicine and proposal for a new classification system: Guidance from the SSC of the ISTH. J. Thromb. Haemost. 2018, 16, 1895–1900. [Google Scholar] [CrossRef] [PubMed]
- Kon, E.; Matteo, B.D.; Delgado, D.; Cole, B.J.; Dorotei, A.; Dragoo, J.L.; Filardo, G.; Fortier, L.A.; Giuffrida, A.; Jo, C.H.; et al. Platelet-rich plasma for the treatment of knee osteoarthritis: An expert opinion and proposal for a novel classification and coding system. Expert Opin. Biol. Ther. 2020, 20, 1447–1460. [Google Scholar] [CrossRef] [PubMed]
- Krüger, J.P.; Freymann, U.; Vetterlein, S.; Neumann, K.; Endres, M.; Kaps, C. Bioactive Factors in Platelet-Rich Plasma Obtained by Apheresis. Orig. Artic. Transfus Med. Hemother 2013, 40, 432–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weibrich, G.; Kleis, W.K.G.; Hafner, G.; Hitzler, W.E. Growth factor levels in platelet-rich plasma and correlations with donor age, sex, and platelet count. J. Craniomaxillofac. Surg. 2002, 30, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.C.; Yu, T.Y.; Chang, G.J.; Lin, L.P.; Lin, M.S.; Pang, J.H.S. Platelet-Rich Plasma Releasate Promotes Regeneration and Decreases Inflammation and Apoptosis of Injured Skeletal Muscle. Am. J. Sports Med. 2018, 46, 1980–1986. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Knapik, D.M.; Polce, E.M.; Eikani, C.K.; Bjornstad, A.H.; Gursoy, S.; Perry, A.K.; Westrick, J.C.; Yanke, A.B.; Verma, N.N.; et al. Relative Efficacy of Intra-articular Injections in the Treatment of Knee Osteoarthritis: A Systematic Review and Network Meta-analysis. Am. J. Sports Med. 2021. [Google Scholar] [CrossRef]
- Tan, J.; Chen, H.; Zhao, L.; Huang, W. Platelet-Rich Plasma Versus Hyaluronic Acid in the Treatment of Knee Osteoarthritis: A Meta-analysis of 26 Randomized Controlled Trials. Arthrosc. J. Arthrosc. Relat. Surg. 2021, 37, 309–325. [Google Scholar] [CrossRef]
- Miroshnychenko, O.; Chalkley, R.J.; Leib, R.D.; Everts, P.A.; Dragoo, J.L. Proteomic analysis of platelet-rich and platelet-poor plasma. Regen. Ther. 2020, 15, 226–235. [Google Scholar] [CrossRef]
- Bansal, H.; Leon, J.; Pont, J.L.; Wilson, D.A.; Bansal, A.; Agarwal, D.; Preoteasa, I. Platelet-rich plasma (PRP) in osteoarthritis (OA) knee: Correct dose critical for long term clinical efficacy. Sci. Rep. 2021, 11, 3971. [Google Scholar] [CrossRef] [PubMed]
- Zahir, H.; Dehghani, B.; Yuan, X.; Chinenov, Y.; Kim, C.; Burge, A.; Bandhari, R.; Nemirov, D.; Fava, P.; Moley, P.; et al. In vitro responses to platelet-rich-plasma are associated with variable clinical outcomes in patients with knee osteoarthritis. Sci. Rep. 2021, 11, 11493. [Google Scholar] [CrossRef]
- Hahn, O.; Kieb, M.; Jonitz-Heincke, A.; Bader, R.; Peters, K.; Tischer, T. Dose-Dependent Effects of Platelet-Rich Plasma Powder on Chondrocytes In Vitro. Am. J. Sports Med. 2020, 48, 1727–1734. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C. Identifying the hub gene and immune infiltration of osteoarthritis by bioinformatical methods. Clin. Rheumatol. 2021, 40, 1027–1037. [Google Scholar] [CrossRef]
- Thijssen, E.; Van Caam, A.; Van Der Kraan, P.M. Obesity and osteoarthritis, more than just wear and tear: Pivotal roles for inflamed adipose tissue and dyslipidaemia in obesity-induced osteoarthritis. Rheumatology 2015, 54, 588–600. [Google Scholar] [CrossRef] [Green Version]
- Werdyani, S.; Liu, M.; Zhang, H.; Sun, G.; Furey, A.; Randell, E.W.; Rahman, P.; Zhai, G. Endotypes of primary osteoarthritis identified by plasma metabolomics analysis. Rheumatology 2021, 60, 2735. [Google Scholar] [CrossRef]
- Mobasheri, A.; Kapoor, M.; Ali, S.A.; Lang, A.; Madry, H. The future of deep phenotyping in osteoarthritis: How can high throughput omics technologies advance our understanding of the cellular and molecular taxonomy of the disease? Osteoarthr. Cartil. Open 2021, 3, 100144. [Google Scholar] [CrossRef]
- Mobasheri, A.; Saarakkala, S.; Finnilä, M.; Karsdal, M.A.; Bay-Jensen, A.C.; van Spil, W.E. Recent advances in understanding the phenotypes of osteoarthritis. F1000Research 2019, 8, 2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, D.L.; Wolfe, F.; Huizinga, T.W.J. Rheumatoid arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Sharma, L. Osteoarthritis of the Knee. N. Engl. J. Med. 2021, 384, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Rosshirt, N.; Trauth, R.; Platzer, H.; Tripel, E.; Nees, T.A.; Lorenz, H.M.; Tretter, T.; Moradi, B. Proinflammatory T cell polarization is already present in patients with early knee osteoarthritis. Arthritis Res. Ther. 2021, 23, 37. [Google Scholar] [CrossRef] [PubMed]
- Mathiessen, A.; Conaghan, P.G. Synovitis in osteoarthritis: Current understanding with therapeutic implications. Arthritis Res. Ther. 2017, 19, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodell-May, J.E.; Sommerfeld, S.D. Role of Inflammation and the Immune System in the Progression of Osteoarthritis. J. Orthop. Res. 2020, 38, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Culemann, S.; Grüneboom, A.; Nicolás-Ávila, J.Á.; Weidner, D.; Lämmle, K.F.; Rothe, T.; Quintana, J.A.; Kirchner, P.; Krljanac, B.; Eberhardt, M.; et al. Locally renewing resident synovial macrophages provide a protective barrier for the joint. Nature 2019, 572, 670–675. [Google Scholar] [CrossRef]
- Daghestani, H.N.; Pieper, C.F.; Kraus, V.B. Soluble macrophage biomarkers indicate inflammatory phenotypes in patients with knee osteoarthritis. Arthritis Rheumatol. 2015, 67, 956–965. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, M.F.; Zhang, X.; Wellman, S.S.; Bolognesi, M.P.; Kraus, V.B. Synergistic Roles of Macrophages and Neutrophils in Osteoarthritis Progression. Arthritis Rheumatol. 2021, 73, 89–99. [Google Scholar] [CrossRef]
- Escobar, G.; Escobar, A.; Ascui, G.; Tempio, F.; Ortiz, M.C.; Pérez, C.A.; López, M.N. Pure platelet-rich plasma and supernatant of calcium-activated P-PRP induce different phenotypes of human macrophages. Regen. Med. 2018, 13, 427–441. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, R.; Toyoda, E.; Maehara, M.; Wasai, S.; Omura, H.; Watanabe, M.; Sato, M. Effect of Platelet-Rich Plasma on M1/M2 Macrophage Polarization. Int. J. Mol. Sci. 2021, 22, 2336. [Google Scholar] [CrossRef]
- O’Donnell, C.; Migliore, E.; Grandi, F.C.; Koltsov, J.; Lingampalli, N.; Cisar, C.; Indelli, P.F.; Sebastiano, V.; Robinson, W.H.; Bhutani, N.; et al. Platelet-Rich Plasma (PRP) From Older Males With Knee Osteoarthritis Depresses Chondrocyte Metabolism and Upregulates Inflammation. J. Orthop. Res. 2019, 37, 1760–1770. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Jo, C.H.; Lee, S.Y.; Yoon, K.S.; Oh, S.; Shin, S. Allogeneic Platelet-Rich Plasma Versus Corticosteroid Injection for the Treatment of Rotator Cuff Disease: A Randomized Controlled Trial. J. Bone Joint Surg. Am. 2020, 102, 2129–2137. [Google Scholar] [CrossRef] [PubMed]
- Andia, I.; Atilano, L.; Maffulli, N. Moving toward targeting the right phenotype with the right platelet-rich plasma (PRP) formulation for knee osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2021, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mimpen, J.Y.; Baldwin, M.J.; Cribbs, A.P.; Philpott, M.; Carr, A.J.; Dakin, S.G.; Snelling, S.J.B. Interleukin-17A Causes Osteoarthritis-Like Transcriptional Changes in Human Osteoarthritis-Derived Chondrocytes and Synovial Fibroblasts In Vitro. Front. Immunol. 2021, 12, 676173. [Google Scholar] [CrossRef]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 Family of Cytokines in Health and Disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef]
- Andia, I.; Maffulli, N. Platelet-rich plasma for managing pain and inflammation in osteoarthritis. Nat. Rev. Rheumatol. 2013, 9, 721–730. [Google Scholar] [CrossRef]
- MC, C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. NF-κB Signaling Pathways in Osteoarthritic Cartilage Destruction. Cells 2019, 8, 734. [Google Scholar] [CrossRef] [Green Version]
- Ostojic, M.; Zevrnja, A.; Vukojevic, K.; Soljic, V. Immunofluorescence Analysis of NF-kB and iNOS Expression in Different Cell Populations during Early and Advanced Knee Osteoarthritis. Int. J. Mol. Sci. 2021, 22, 6461. [Google Scholar] [CrossRef]
- Pan, H.; Dai, H.; Wang, L.; Lin, S.; Tao, Y.; Zheng, Y.; Jiang, R.; Fang, F.; Wu, Y. MicroRNA-410-3p modulates chondrocyte apoptosis and inflammation by targeting high mobility group box 1 (HMGB1) in an osteoarthritis mouse model. BMC Musculoskelet. Disord. 2020, 21, 486. [Google Scholar] [CrossRef]
- Aulin, C.; Lassacher, T.; Palmblad, K.; Harris, H.E. Early stage blockade of the alarmin HMGB1 reduces cartilage destruction in experimental OA. Osteoarthr. Cartil. 2020, 28, 698–707. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Ding, L.; Yin, S.; Huang, Z.; Zhang, L.; Mei, W.; Wu, P.; Wang, P.; Pan, K. Relationship between the pyroptosis of fibroblast-like synoviocytes and HMGB1 secretion in knee osteoarthritis. Mol. Med. Rep. 2021, 23, 97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, F.; Augi, T.; Williamson, K.M.; Onishi, K.; Hogan, M.V.; Neal, M.D.; Wang, J.H.-C. Platelet HMGB1 in Platelet-Rich Plasma (PRP) promotes tendon wound healing. PLoS ONE 2021, 16, e0251166. [Google Scholar] [CrossRef] [PubMed]
- Urits, I.; Viswanath, O.; Galasso, A.C.; Sottosani, E.R.; Mahan, K.M.; Aiudi, C.M.; Kaye, A.D.; Orhurhu, V.J. Platelet-Rich Plasma for the Treatment of Low Back Pain: A Comprehensive Review. Curr. Pain Headache Rep. 2019, 23, 52. [Google Scholar] [CrossRef]
- Pinho-Ribeiro, F.A.; Verri, W.A.; Chiu, I.M. Nociceptor Sensory Neuron-Immune Interactions in Pain and Inflammation. Trends Immunol. 2017, 38, 5–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cells | Whole Blood | PPP Biobank | PRP Biobank |
|---|---|---|---|
| Platelets (×103/µL) | 220 ± 0.42 | 24 ± 13 | 1972 ± 325 (×8.97) |
| Leukocytes (×103/µL) | 5.2 ± 0.87 | n.d. | 3.91 ± 5.02 (0.56–11.39) (lymphocytes) |
| Erythrocytes (×106/µL) | 4.17 ± 0.48 | n.d. | n.d. |
| Cells | Whole Blood | PPP in-House | PRP in-House |
|---|---|---|---|
| Platelets (×103/µL) | 226 ± 41 | 15 ± | 460 ± 103 (×2.03) |
| Leukocytes (×103/µL) | 4.9 ± 0.74 | n.d. | 0.06 ± 0.07 (lymphocytes) |
| Erythrocytes (×106/µL) | 4.38 ± 0.62 | n.d. | n.d. |
| Platelet-Rich Plasma (PRP) | Platelet-Poor Plasma (PPP) | p-Value | |
|---|---|---|---|
| Total protein | 243.78 | 225.16 | |
| (BCA) mg/mL | 264.91 | 251.96 | |
| 193.61 | 246.44 | ||
| 234.75 | 219.07 | ||
| 233.07 | 223.61 | ||
| 262.4 | 240.98 | ||
| Mean (SD) | 238.83 ± 25.72 | 234.50 ± 13.88 | |
| Interdonor %CV | 10.77% | 5.92% | p = 1.000 |
| PDGF ng/mL | 23.77 | 5.5 | |
| 24.13 | 5.59 | ||
| 38.31 | 5.79 | ||
| 17.12 | 5.86 | ||
| 29.7 | 6.32 | ||
| 21.75 | 5.33 | ||
| Mean (SD) | 25.78 ± 7.36 | 5.73 ± 0.35 | p = 0.002 |
| Interdonor %CV | 28.54% | 6.10% | |
| VEGF pg/mL | 277.19 | 94.17 | |
| 639.08 | 145.42 | ||
| 901.23 | 215.21 | ||
| 725.91 | 273.50 | ||
| 751.44 | 223.59 | ||
| 604.32 | 170.69 | ||
| Mean (SD) | 649.85 ± 209.91 | 187.10 ± 63.60 | p = 0.002 |
| Interdonor %CV | 32.30% | 33.99% | |
| MCP1 pg/mL | 429.25 | 384.70 | |
| 580.25 | 322.13 | ||
| 709.19 | 541.16 | ||
| 632.09 | 497.11 | ||
| 428.35 | 357.47 | ||
| 397.30 | 369.82 | ||
| Mean (SD) | 529.38 ± 128.97 | 412.07 ± 86.62 | p = 0.065 |
| Interdonor %CV | 24.36% | 21.02% | |
| RANTES pg/mL | 1080.25 | 71.66 | |
| 1227.34 | 116.38 | ||
| 1462.00 | 90.40 | ||
| 1001.23 | 90.30 | ||
| 849.46 | 90.75 | ||
| 1183.94 | 110.01 | ||
| Mean (SD) | 1133.92 ± 209.89 | 94.94 ± 16.05 | p = 0.002 |
| Interdonor %CV | 18.51% | 16.90% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Amo, C.; Perez-Valle, A.; Atilano, L.; Andia, I. Unraveling the Signaling Secretome of Platelet-Rich Plasma: Towards a Better Understanding of Its Therapeutic Potential in Knee Osteoarthritis. J. Clin. Med. 2022, 11, 473. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11030473
Del Amo C, Perez-Valle A, Atilano L, Andia I. Unraveling the Signaling Secretome of Platelet-Rich Plasma: Towards a Better Understanding of Its Therapeutic Potential in Knee Osteoarthritis. Journal of Clinical Medicine. 2022; 11(3):473. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11030473
Chicago/Turabian StyleDel Amo, Cristina, Arantza Perez-Valle, Leire Atilano, and Isabel Andia. 2022. "Unraveling the Signaling Secretome of Platelet-Rich Plasma: Towards a Better Understanding of Its Therapeutic Potential in Knee Osteoarthritis" Journal of Clinical Medicine 11, no. 3: 473. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11030473