Elevated Serum Melatonin under Constant Darkness Enhances Neural Repair in Spinal Cord Injury through Regulation of Circadian Clock Proteins Expression


Abstract
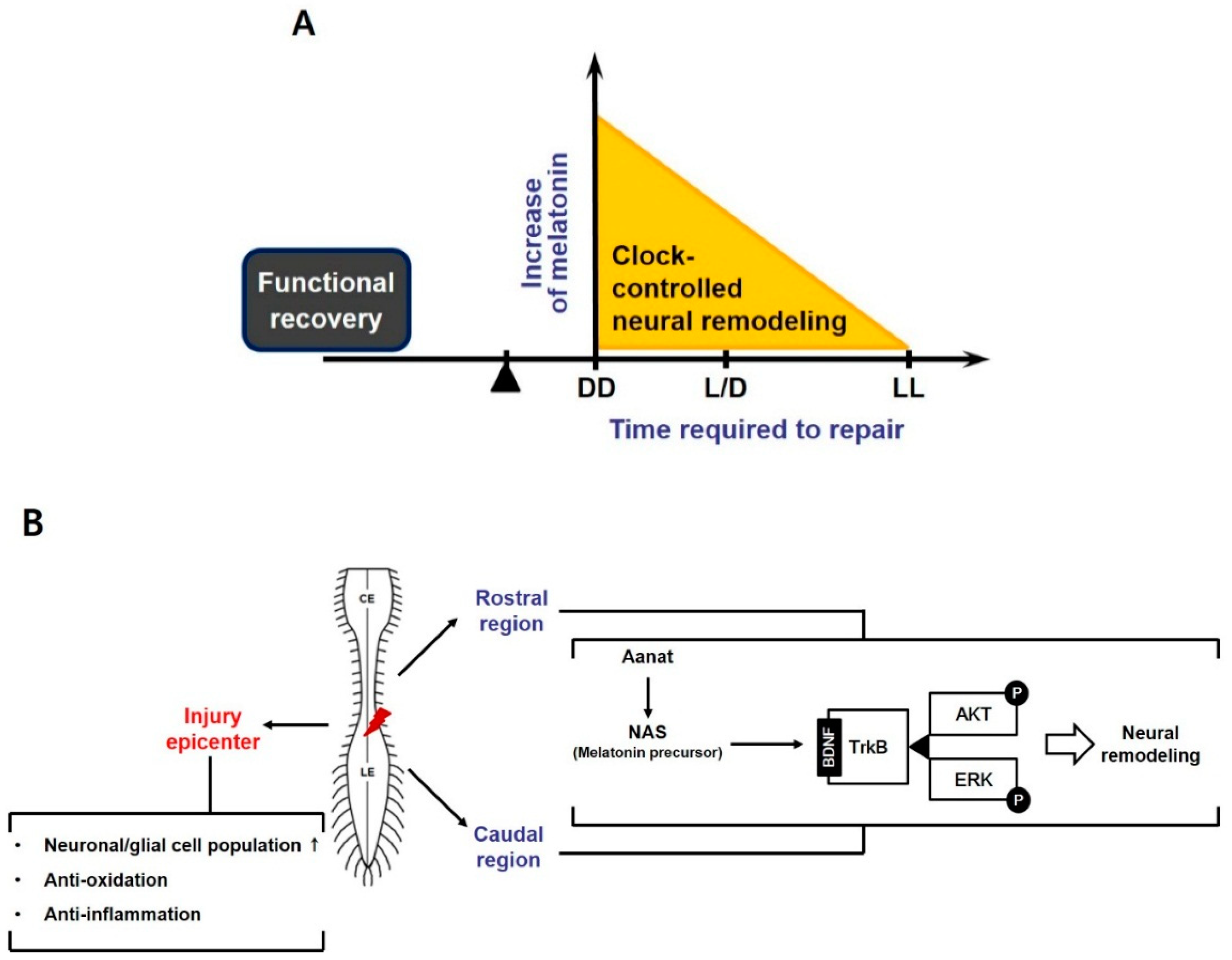
:1. Introduction
2. Materials and Methods
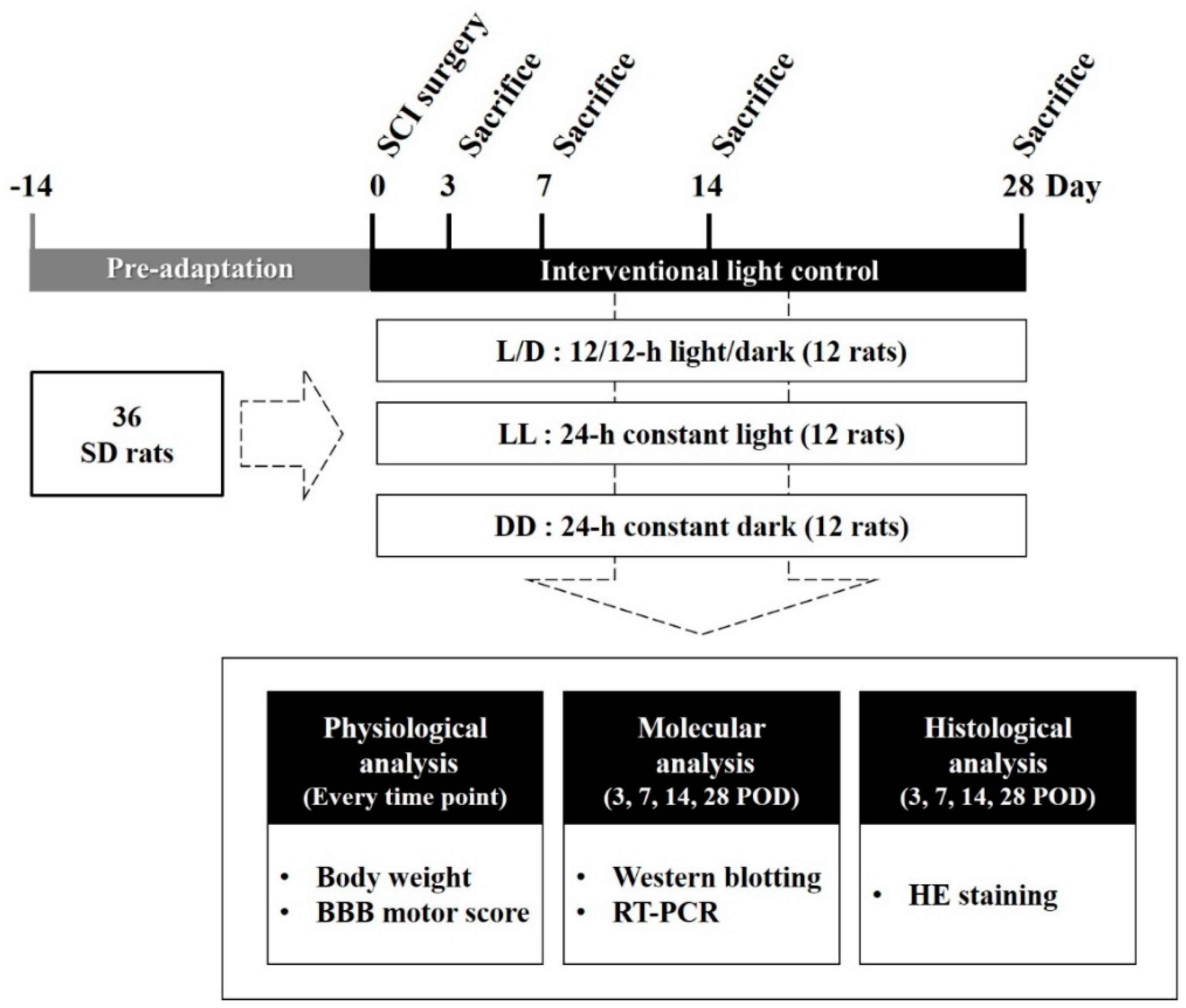
2.1. Experimental Groups
2.2. Surgical Procedures for Experimental SCI
2.3. Assessment of Motor Function
2.4. Measurement of Melatonin Concentration in Cerebrospinal Fluid
2.5. RNA Isolation and Quantificative RT-PCR
2.6. Protein Extraction and Western Blotting
2.7. Hematoxylin and Eosin Staining
2.8. Golgi-Cox Staining and Sholl Analysis
2.9. Statistical Analysis
3. Results
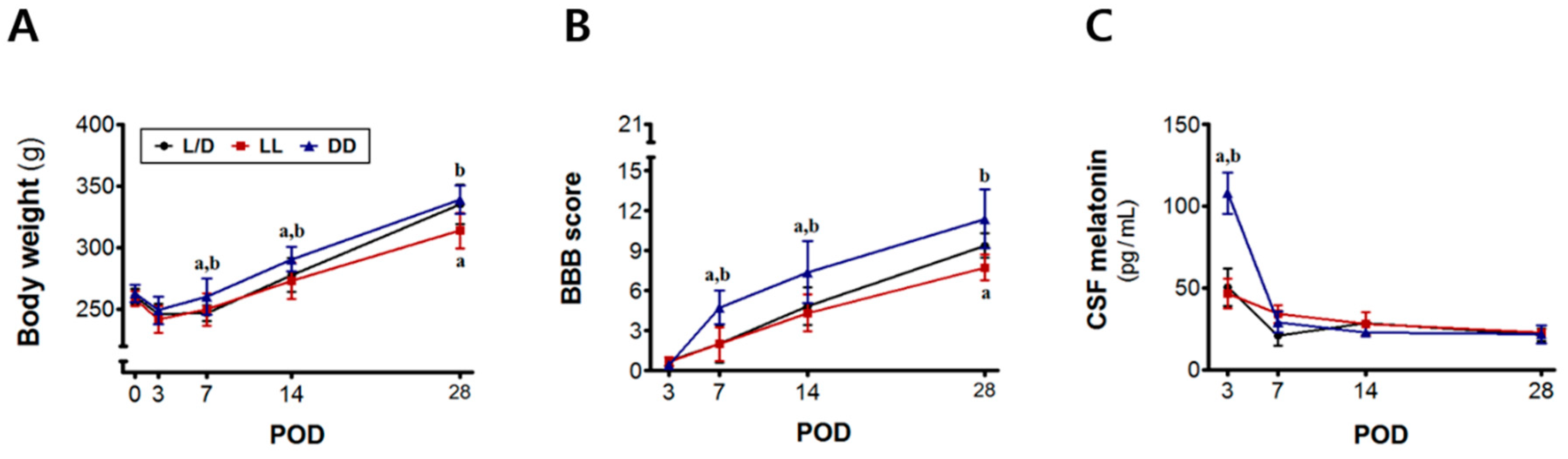
3.1. Time-Dependent Changes in Body Weight, Motor Function, and Melatonin Levels
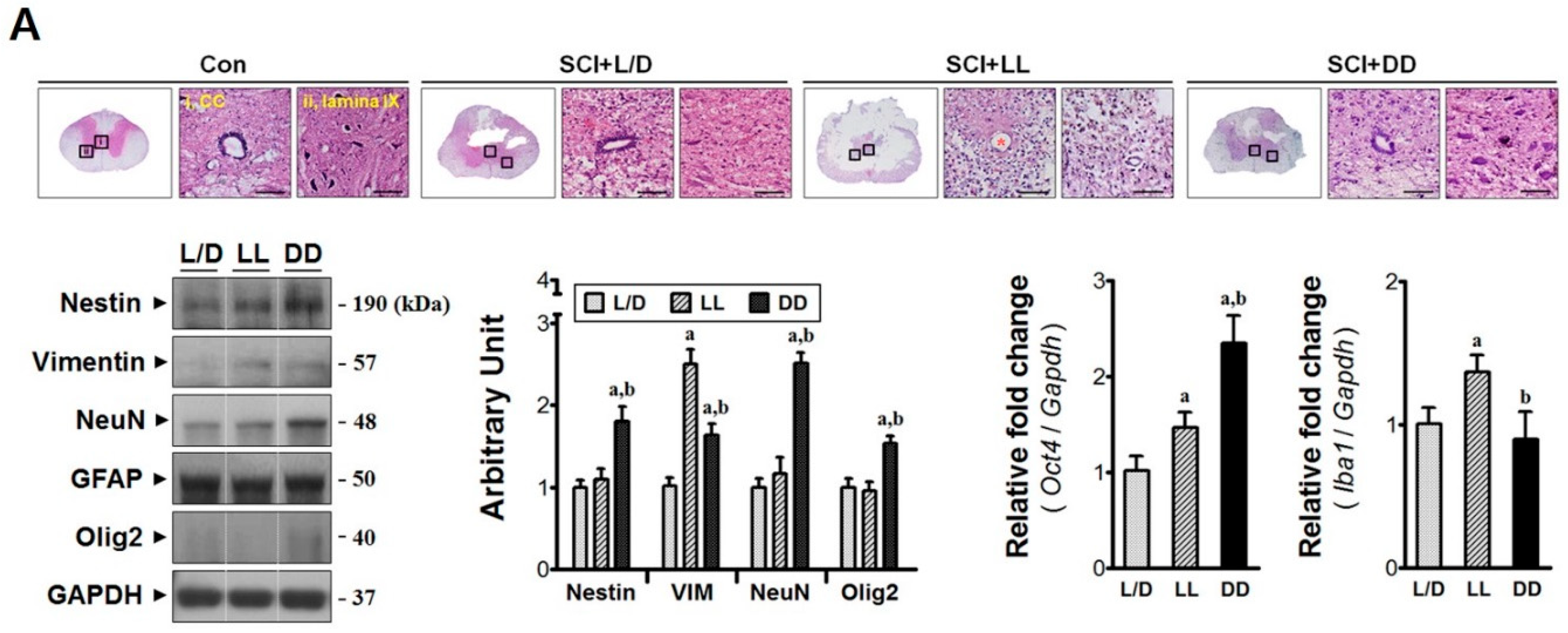
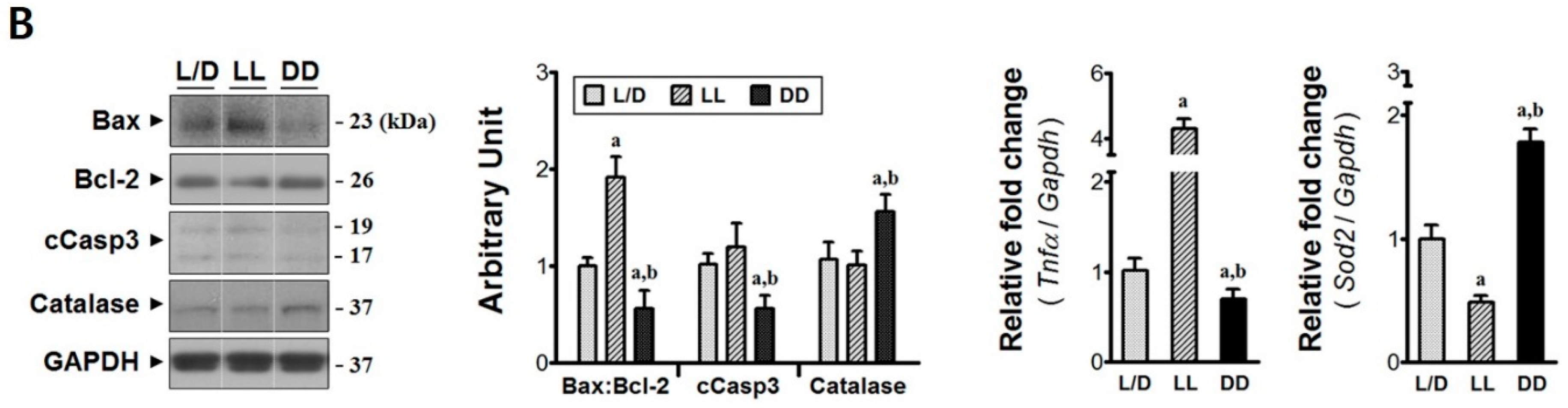
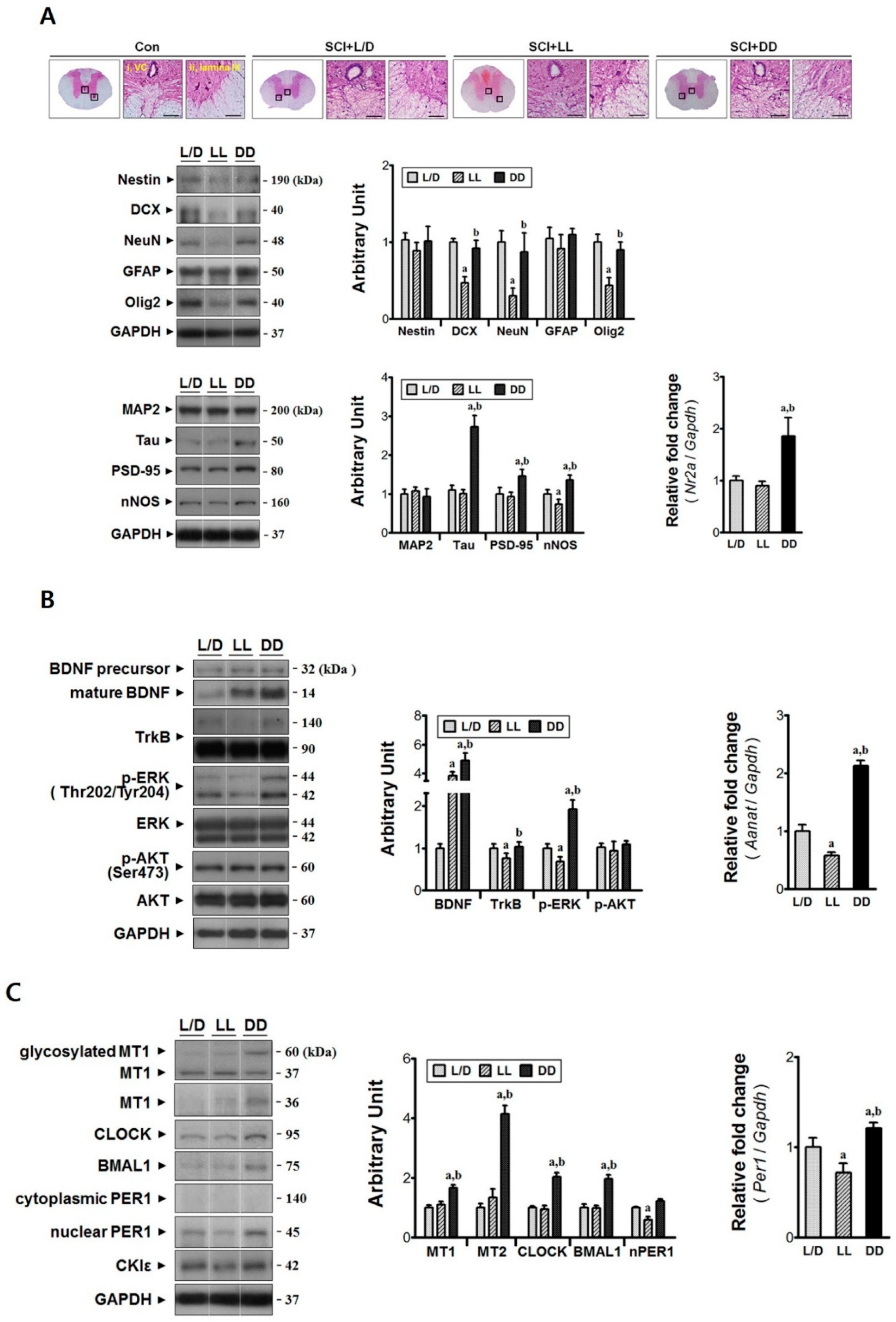
3.2. Elevated Melatonin during Acute Phase Brings Molecular Changes at the Injury Epicenter
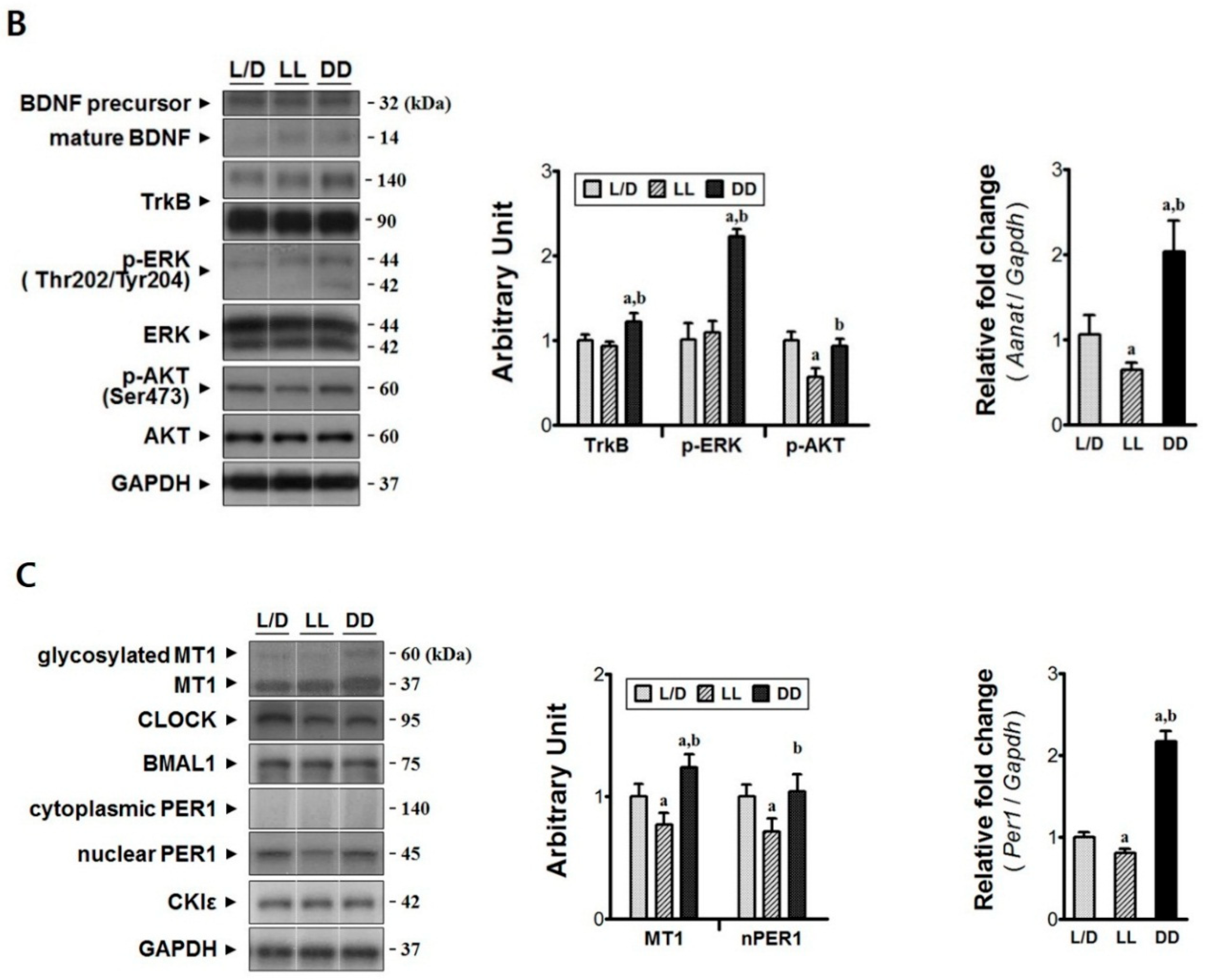
3.3. Constant Dark Condition Accelerates Recovery of the Rostral Region via Trk-ERK Signaling Pathway
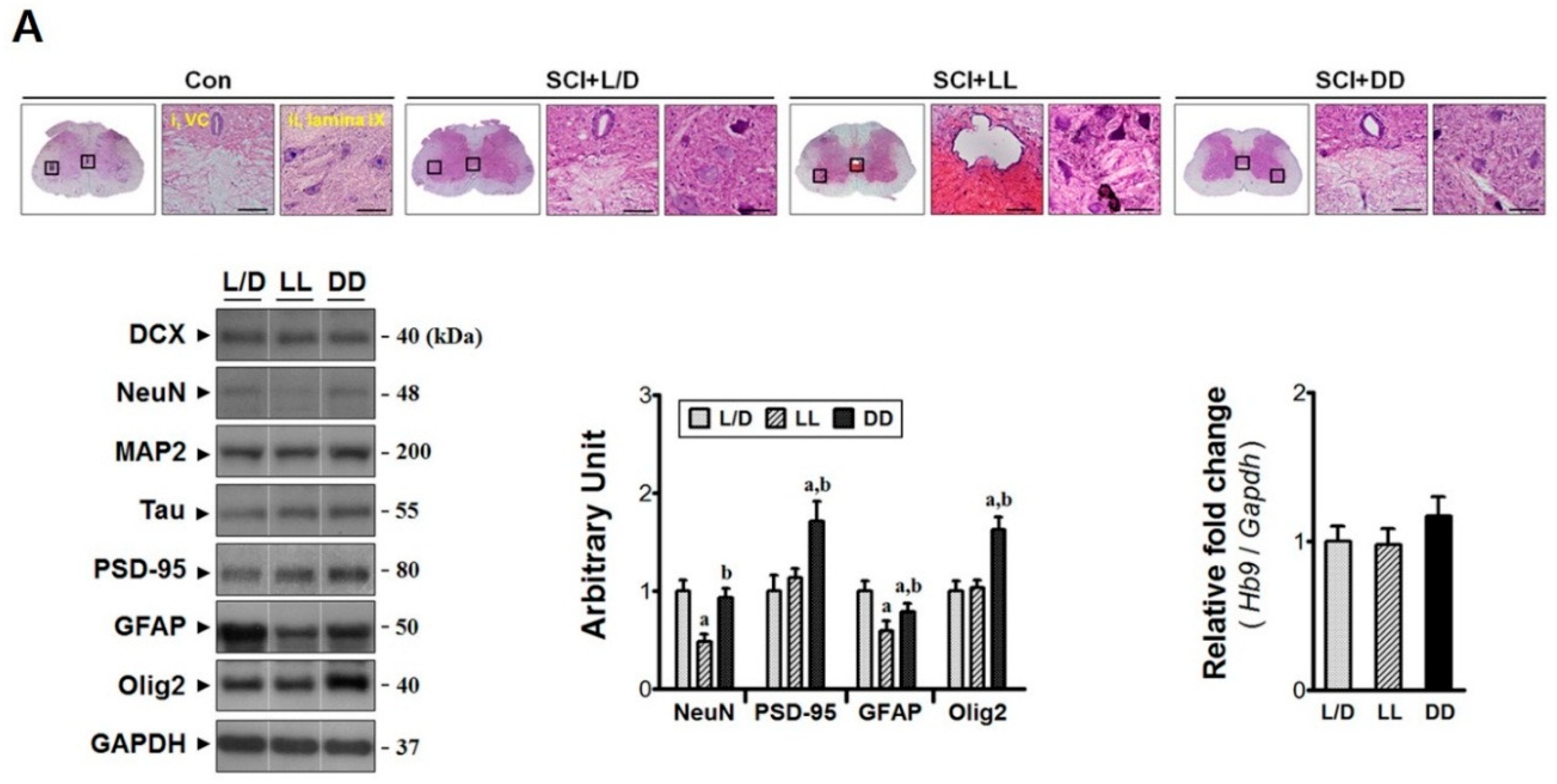
3.4. Constant Dark Condition Preserves Excitatory Circuits in the Lumbar Region
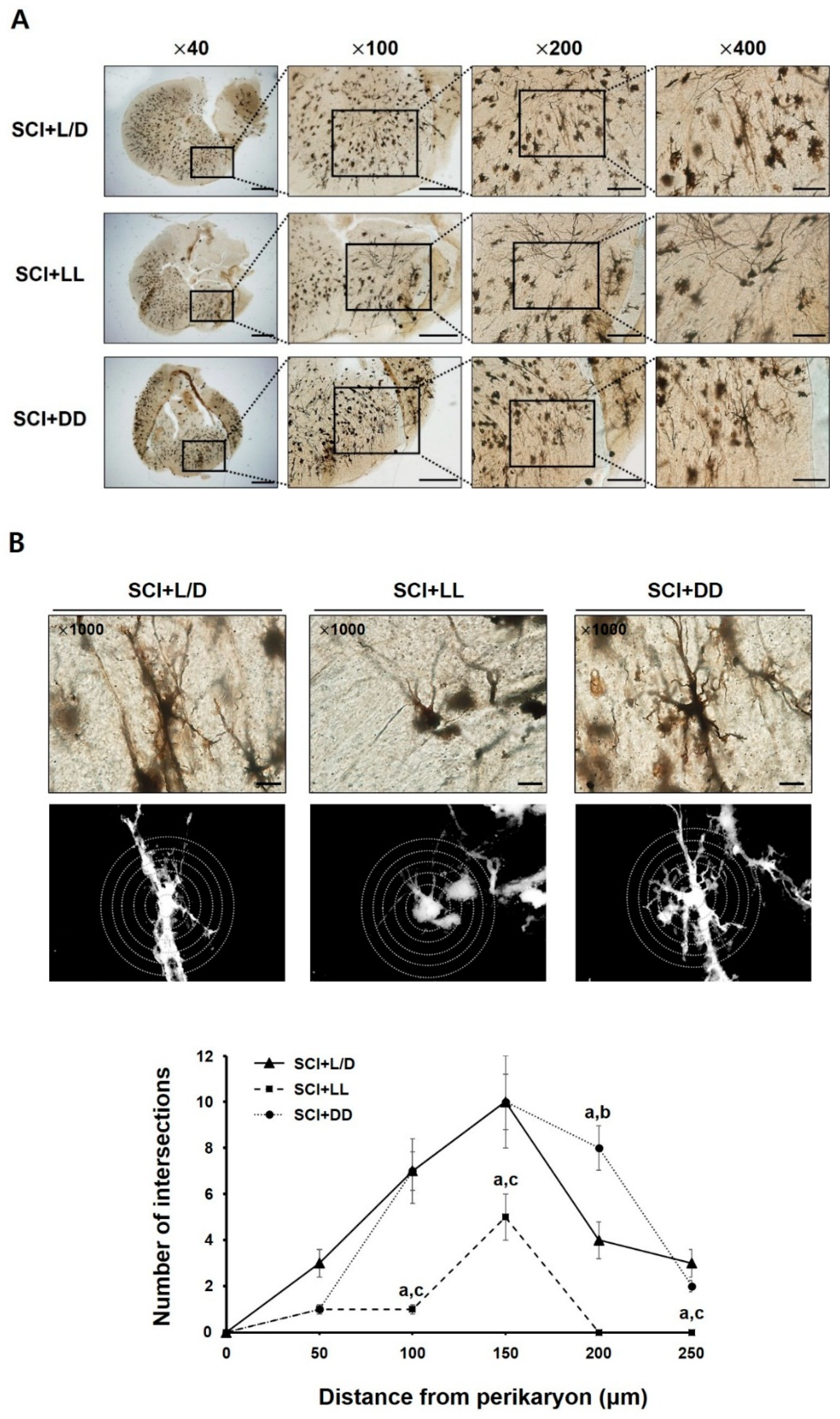
3.5. The Influences of Light/Dark Environment on the Recovery of Injured Cord Evidenced by Histological Examination
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silva, N.A.; Sousa, N.; Reis, R.L.; Salgado, A.J. From basics to clinical: A comprehensive review on spinal cord injury. Prog. Neurobiol. 2014, 114, 25–57. [Google Scholar] [CrossRef] [PubMed]
- Oyinbo, C.A. Secondary injury mechanisms in traumatic spinal cord injury: A nugget of this multiply cascade. Acta Neurobiol. Exp. 2011, 71, 281–299. [Google Scholar]
- Donnelly, D.J.; Popovich, P.G. Inflammation and its role in neuroprotection, axonal regeneration and functional recovery after spinal cord injury. Exp. Neurol. 2008, 209, 378–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abematsu, M.; Tsujumura, K.; Yamano, M.; Saito, M.; Kohno, K.; Kohyama, J.; Namihira, M.; Komiya, S.; Nakashima, K. Neurons derived from transplanted neural stem cells restore disrupted neuronal circuitry in a mouse model of spinal cord injury. J. Clin. Investig. 2010, 120, 3255–3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutts, M.; Keirstead, H.S. Stem cells for the treatment of spinal cord injury. Exp. Neurol. 2008, 209, 368–377. [Google Scholar] [PubMed]
- Lee-Kubli, C.A.; Lu, P. Induced pluripotent stem cell-derived neural stem cell therapies for spinal cord injury. Neural Regen. Res. 2015, 10, 10–16. [Google Scholar] [PubMed]
- Meletis, K.; Barnabe-Heider, F.; Carlen, M.; Evergren, E.; Tomilin, N.; Shupliakov, O.; Frisen, J. Spinal cord injury reveals multilineage differentiation of ependymal cells. PLoS Biol. 2008, 6, e182. [Google Scholar] [CrossRef]
- Cao, Q.L.; Howard, R.M.; Dennison, J.B.; Whittemore, S.R. Differentiation of engrafted neuronal-restricted precursor cells is inhibited in the traumatically injured spinal cord. Exp. Neurol. 2008, 177, 349–359. [Google Scholar] [CrossRef]
- Flynn, J.R.; Graham, B.A.; Galea, M.P.; Callister, R.J. The role of propriospinal interneurons in recovery from spinal cord injury. Neuropharmacology 2011, 60, 809–822. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, S.; Lee, S.R.; Park, K.; Hong, Y.; Lee, M.; Park, S.; Jin, Y.; Chang, K.T.; Hong, Y. Beneficial effects of melatonin combined with exercise on endogenous neural stem/progenitor cells proliferation after spinal cord injury. Int. J. Mol. Sci. 2014, 15, 2207–2222. [Google Scholar] [CrossRef]
- Kaptanoglu, E.; Tuncel, M.; Palaoglu, S.; Konan, A.; Demirpence, E.; Kilinc, K. Comparison of the effects of melatonin and methylprednisolone in experimental spinal cord injury. J. Neurosurg. 2000, 93, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Cuzzocrea, S.; Costantino, G.; Gitto, E.; Mazzon, E.; Fulia, F.; Serraino, I.; Cordaro, S.; Barberi, I.; De Sarro, A.; Caputi, A.P. Protective effects of melatonin in ischemic brain injury. J. Pineal Res. 2000, 29, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, E.; Bryant-Thomas, T.; Pacheco Quinto, J.; Henry, T.L.; Poeggeler, B.; Herbert, D.; Cruz-Sanchez, F.; Chyan, Y.J.; Smith, M.A.; Perry, G.; et al. Melatonin increases survival and inhibits oxidative and amyloid pathology in a transgenic model of Alzheimer’s disease. J. Neurochem. 2003, 85, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, T.; Nakamura, T.; Ikeda, T.; Takagi, K. Potent protective effects of melatonin on experimental spinal cord injury. Spine 2000, 25, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Genovese, T.; Mazzon, E.; Muia, C.; Bramanti, P.; De Sarro, A.; Cuzzocrea, S. Attenuation in the evolution of experimental spinal cord trauma by treatment with melatonin. J. Pineal Res. 2005, 38, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Lewy, A.J.; Wehr, T.A.; Goodwin, F.K.; Newsome, D.A.; Markey, S.P. Light suppresses melatonin secretion in humans. Science 1980, 210, 1267–1269. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, S.K.; Park, K.; Lee, Y.; Hong, Y.; Lee, S.; Jeon, J.C.; Kim, J.H.; Lee, S.R.; Chang, K.T.; et al. Beneficial effects of endogenous and exogenous melatonin on neural reconstruction and functional recovery in an animal model of spinal cord injury. J. Pineal Res. 2012, 52, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.W.; Liu, X.; Pradoldej, S.; Tosini, G.; Chang, Q.; Luvone, P.M.; Ye, K. N-acetylserotonin activates TrkB receptor in a circadian rhythm. Proc. Natl. Acad. Sci. USA 2010, 107, 3876–3881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, D.C.; Coon, S.L.; Roseboom, P.H.; Weller, J.L.; Bernard, M.; Gastel, J.A.; Zatz, M.; Luvone, P.M.; Rodriguez, I.R.; Begay, V.; et al. The melatonin rhythm-generating enzyme: Molecular regulation of serotonin N-acetyltransferase in the pineal gland. Recent Prog. Horm. Res. 1997, 52, 307–357. [Google Scholar] [PubMed]
- Choudhury, A.; Singh, S.; Palit, G.; Shukla, S.; Ganguly, S. Administration of N-acetylserotonin and melatonin alleviate chronic ketamine-induced behavioural phenotype accompanying BDNF-independent and dependent converging cytoprotective mechanisms in the hippocampus. Behav. Brain Res. 2016, 297, 204–212. [Google Scholar] [CrossRef]
- Illnerova, H.; Vanecek, J. Entrainment of the rat pineal rhythm in melatonin production by light. Reprod. Nutr. Dev. 1988, 28, 515–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.; Lee, Y.; Park, S.; Lee, S.; Hong, Y.; Lee, S.K.; Hong, Y. Synergistic effect of melatonin on exercise-induced neuronal reconstruction and functional recovery in a spinal cord injury animal model. J. Pineal Res. 2010, 48, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A sensitive and reliable locomotor rating scale for open field testing in rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Coelho, L.A.; Peres, R.; Amaral, F.G.; Reiter, R.J.; Cipolla-Neto, J. Daily differential expression of melatonin-related genes and clock genes in rat cumulus-oocyte complex: Changes after pinealectomy. J. Pineal Res. 2015, 58, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Conrad, S.; Schluesener, H.J.; Adibzahdeh, M.; Schwab, J.M. Spinal cord injury induction of lesional expression of profibrotic and angiogenic connective tissue growth factor confined to reactive astrocytes, invading fibroblasts and endothelial cells. J. Neurosurg. Spine 2005, 2, 319–326. [Google Scholar] [CrossRef]
- Kim, D.H.; Heo, S.D.; Ahn, M.J.; Sim, K.B.; Shin, T.K. Activation of embryonic intermediate filaments contributes to glial scar formation after spinal cord injury in rats. J. Vet. Sci. 2003, 4, 109–112. [Google Scholar] [PubMed]
- Bodega, G.; Suarez, I.; Rubio, M.; Fernandez, B. Ependyma: Phylogenetic evolution of glial fibrillary acidic protein (GFAP) and vimentin expression in vertebrate spinal cord. Histochemistry 1994, 102, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Victorov, I.V.; Prass, K.; Dimagl, U. Improved selective, simple, and contrast staining of acidophilic neurons with vanadium acid fuchsin. Brain Res. Brain Res. Protoc. 2000, 5, 135–139. [Google Scholar] [CrossRef]
- Wuermser, L.A.; Ho, C.H.; Chiodo, A.E.; Priebe, M.M.; Kirshclum, S.C.; Scelza, W.M. Spinal cord injury medicine. 2. Acute care management of traumatic and nontraumatic injury. Arch. Phys. Med. Rehabil. 2007, 88, S55–S61. [Google Scholar] [CrossRef]
- Fehlings, M.G.; Cadotte, D.W.; Fehlings, L.N. A series of systematic reviews on the treatment of acute spinal cord injury: A foundation for best medical practice. J. Neurotrauma 2011, 28, 1329–1333. [Google Scholar] [CrossRef]
- Wright, H.R.; Lack, L.C.; Kennaway, D.J. Differential effects of light wavelength in phase advancing the melatonin rhythm. J. Pineal Res. 2004, 36, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Zawilska, J.B.; Wawrocka, M. Chick retina and pineal gland differentially respond to constant light and darkness: In vivo studies on serotonin N-acetyltransferase (NAT) activity and melatonin content. Neurosci. Lett. 1993, 153, 21–24. [Google Scholar] [CrossRef]
- Oliveira, C.; Ortega, A.; Lopez-Olmeda, J.F.; Vera, L.M.; Sanchez-Vazquez, F.J. Influence of constant light and darkness, light intensity, and light spectrum on plasma melatonin rhythms in senegal sole. Chronobiol. Int. 2007, 24, 615–627. [Google Scholar] [CrossRef]
- Masana, M.I.; Benloucif, S.; Dubocovich, M.L. Circadian rhythm of mt1 melatonin receptor expression in the suprachiasmatic nucleus of the C3H/HeN mouse. J. Pineal Res. 2000, 28, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.B.; Tang, T.S.; Yang, H.L.; Xiao, D.S. Antioxidation of melatonin against spinal cord injury in rats. Chin. Med. J. 2004, 117, 571–575. [Google Scholar] [PubMed]
- Chern, C.M.; Liao, J.F.; Wang, Y.H.; Shen, Y.C. Melatonin ameliorates neural function by promoting endogenous neurogenesis through the MT2 melatonin receptor in ischemic-stroke mice. Free Radic. Biol. Med. 2012, 52, 1634–1647. [Google Scholar] [CrossRef] [PubMed]
- Moriya, T.; Horie, N.; Mitome, M.; Shinohara, K. Melatonin influences the proliferative and differentiative activity of neural stem cells. J. Pineal Res. 2007, 42, 411–418. [Google Scholar] [CrossRef]
- Armstrong, S.M.; Cassone, V.M.; Chesworth, M.J.; Redman, J.R.; Short, R.V. Synchronization of mammalian circadian rhythms by melatonin. J. Neural Transm. Suppl. 1986, 21, 375–394. [Google Scholar]
- Yasuniwa, Y.; Izumi, H.; Wang, K.Y.; Shimajiri, S.; Sasaguri, Y.; Kawai, K.; Kasai, H.; Shimada, T.; Miyake, K.; Kashiwagi, E.; et al. Circadian disruption accelerates tumor growth and angio/stromagenesis through a Wnt signaling pathway. PLoS ONE 2010, 5, e15330. [Google Scholar] [CrossRef]
- Seifman, M.A.; Adamides, A.A.; Nguyen, P.N.; Vallance, S.A.; Cooper, D.J.; Kossmann, T.; Rosenfeld, J.V.; Morganti-Kossmann, M.C. Endogenous melatonin increases in cerebrospinal fluid of patients after severe traumatic brain injury and correlates with oxidative stress and metabolic disarray. J. Cereb. Blood Flow Metab. 2008, 28, 684–696. [Google Scholar] [CrossRef]
- Thofner Hulten, V.D.; Biering-Sorensen, F.; Jorgensen, N.R.; Jennum, P.J. Melatonin and cortisol in individuals with spinal cord injury. Sleep Med. 2018, 51, 92–98. [Google Scholar] [CrossRef]
- Silva, S.O.; Ximenes, V.F.; Livramento, J.A.; Catalani, L.H.; Campa, A. High concentrations of the melatonin metabolite, n1-acetyl-n2-formyl-5-methoxykynuramine, in cerebrospinal fluid of patients with meningitis: A possible immunomodulatory mechanism. J. Pineal Res. 2005, 39, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Schumacher, M. Psychosocial stress affects pineal function in the tree shrew (tupaia belangeri). Physiol. Behav. 1990, 47, 713–717. [Google Scholar] [CrossRef]
- Jockers, R.; Maurice, P.; Boutin, J.A.; Delagrange, P. Melatonin receptors, heterodimerization, signal transduction and binding sites: What’s new? Br. J. Pharmacol. 2008, 154, 1182–1195. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.H.; Wong, Y.H. A molecular and chemical perspective in defining melatonin receptor subtype selectivity. Int. J. Mol. Sci. 2013, 14, 18385–18406. [Google Scholar] [CrossRef] [PubMed]
- Michineau, S.; Alhenc-Gelas, F.; Rajerison, R.M. Human bradykinin B2 receptor sialylation and N-glycosylation participate with disulfide bonding in surface receptor dimerization. Biochemistry 2006, 45, 2699–2707. [Google Scholar] [CrossRef] [PubMed]
- Waly, N.E.; Hallworth, R. Circadian Pattern of Melatonin MT1 and MT2 Receptor Localization in the Rat Suprachiasmatic Nucleus. J. Circadian Rhythm. 2015, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Hunt, A.E.; Al-Ghoul, W.M.; Gillette, M.U.; Dubocovich, M.L. Activation of MT(2) melatonin receptors in rat suprachiasmatic nucleus phase advances the circadian clock. Am. J. Physiol. Cell Physiol. 2001, 280, C110–C118. [Google Scholar] [CrossRef]
- Dubocovich, M.L.; Yun, K.; Al-Ghoul, W.M.; Benloucif, S.; Masana, M.I. Selective MT2 melatonin receptor antagonists block melatonin-mediated phase advances of circadian rhythms. FASEB J. 1998, 12, 1211–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beesley, S.; Lee, J.; Olcese, J. Circadian clock regulation of melatonin MTNR1B receptor expression in human myometrial smooth muscle cells. Mol. Hum. Reprod. 2015, 21, 662–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filli, L.; Schwab, M.E. Structural and functional reorganization of propriospinal connections promotes functional recovery after spinal cord injury. Neural Regen. Res. 2015, 10, 509–513. [Google Scholar] [CrossRef] [PubMed]
- McKinley, W.; Santos, K.; Meade, M.; Brooke, K. Incidence and outcomes of spinal cord injury clinical syndromes. J. Spinal Cord Med. 2007, 30, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Hagglund, M.; Borgius, L.; Dougherty, K.J.; Kiehn, O. Activation of groups of excitatory neurons in the mammalian spinal cord or hindbrain evokes locomotion. Nat. Neurosci. 2010, 13, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Beauparlant, J.; van den Brand, R.; Barraud, Q.; Friedli, L.; Musienko, P.; Dietz, V.; Courtine, G. Undirected compensatory plasticity contributes to neuronal dysfunction after severe spinal cord injury. Brain 2013, 136, 3347–3361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keith, D.; El-Husseini, A. Excitation Control: Balancing PSD-95 Function at the Synapse. Front. Mol. Neurosci. 2008, 1, 4. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, S.K.; Shnitka, T.K. Diversity in Reactive Astrocytes. In Neuroglia in the Aging Brain; de Vellis, J., Ed.; Humana Press: New York, NY, USA, 2002; pp. 17–33. [Google Scholar]
- Karimi-Abdolrezaee, S.; Billakanti, R. Reactive astrogliosis after spinal cord injury-beneficial and detrimental effects. Mol. Neurobiol. 2012, 46, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Wilhelmsson, U.; Bogestal, Y.R.; Pekna, M. The role of astrocytes and complement system in neural plasticity. Int. Rev. Neurobiol. 2007, 82, 95–111. [Google Scholar]
- Adams, M.M.; Hicks, A.L. Spasticity after spinal cord injury. Spinal Cord 2005, 43, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Wideman, C.H.; Murphy, H.M. Constant light induces alterations in melatonin levels, food intake, feed efficiency, visceral adiposity, and circadian rhythms in rats. Nutr. Neurosci. 2009, 12, 233–240. [Google Scholar] [CrossRef]
- Gooley, J.J.; Chamberlain, K.; Smith, K.A.; Khalsa, S.B.; Rajaratnam, S.M.; Van Reen, E.; Zeitzer, J.M.; Czeisler, C.A.; Lockley, S.W. Exposure to room light before bedtime suppresses melatonin onset and shortens melatonin duration in humans. J. Clin. Endocrinol. Metab. 2011, 96, E463–E472. [Google Scholar] [CrossRef]
- Farhadi, N.; Gharghani, M.; Farhadi, Z. Effects of long-term light, darkness and oral administration of melatonin on serum levels of melatonin. Biomed. J. 2016, 39, 81–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) | Size (bp) | GenBank Accession No. |
|---|---|---|---|
| Aanat | F: AAA GTA CAC TCA GGC ACC AAT GT | 110 | NM_012818 |
| R: GGG AAC ATA GCT GCT TTA TTA GTG TCA G | |||
| Hb9 | F: GCA ATC GAA CCT CTT GGG GA | 187 | NM_001271274.1 |
| R: TTT CAT TCG GCG GTT CTG GA | |||
| Iba1 | F: GTC CTT GAA GCG AAT GCT GG | 157 | NM_017196.3 |
| R: CAT TCT CAA GAT GGC AGA TC | |||
| Nr2a | F: TCC ATT CTT CTG TCA TCC TGC | 224 | NM_012573.3 |
| R: AAG ACC GTC TCT CAC TCT TGC | |||
| Oct4 | F: GAG GGA TGG CAT ACT GTG GAC | 272 | XM_228354 |
| R: GGT GTA CCC CAA GGT GAT CC | |||
| Per1 | F: TTT GGA GAG CTG CAA CAT TCC | 101 | NM_011065.4 |
| R: CTG CCC TCT GCT TGT CAT CA | |||
| Sod2 | F: CCG AGG AGA AGT ACC ACG AG | 174 | NM_017051.2 |
| R: GCT TGA TAG CCT CCA GCA AC | |||
| Tnfα | F: CTA CTG AAC TTC GGG GTG ATC | 292 | NM_012675.3 |
| R: CTT GTC CCT TGA AGA GAA CCT G | |||
| Gapdh | F: CTC AGT TGC TGA GGA GTC CC | 120 | NM_017008.4 |
| R: ATT CGA GAG AAG GGA GGG CT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, Y.; Jin, Y.; Park, K.; Choi, J.; Kang, H.; Lee, S.-R.; Hong, Y. Elevated Serum Melatonin under Constant Darkness Enhances Neural Repair in Spinal Cord Injury through Regulation of Circadian Clock Proteins Expression. J. Clin. Med. 2019, 8, 135. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8020135
Hong Y, Jin Y, Park K, Choi J, Kang H, Lee S-R, Hong Y. Elevated Serum Melatonin under Constant Darkness Enhances Neural Repair in Spinal Cord Injury through Regulation of Circadian Clock Proteins Expression. Journal of Clinical Medicine. 2019; 8(2):135. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8020135
Chicago/Turabian StyleHong, Yunkyung, Yunho Jin, Kanghui Park, Jeonghyun Choi, Hyunbon Kang, Sang-Rae Lee, and Yonggeun Hong. 2019. "Elevated Serum Melatonin under Constant Darkness Enhances Neural Repair in Spinal Cord Injury through Regulation of Circadian Clock Proteins Expression" Journal of Clinical Medicine 8, no. 2: 135. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8020135