The Oncogenic Activity of miR-29b-1-5p Induces the Epithelial-Mesenchymal Transition in Oral Squamous Cell Carcinoma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Surgical Specimens
2.2. Laser Capture Microdissection
2.3. Immunohistochemistry
2.4. Cell Culture
2.5. Transient Transfection
2.6. Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR) and qPCR
2.7. Immunoblotting
2.8. Luciferase Reporter Assay
2.9. Statistical Analysis
3. Results
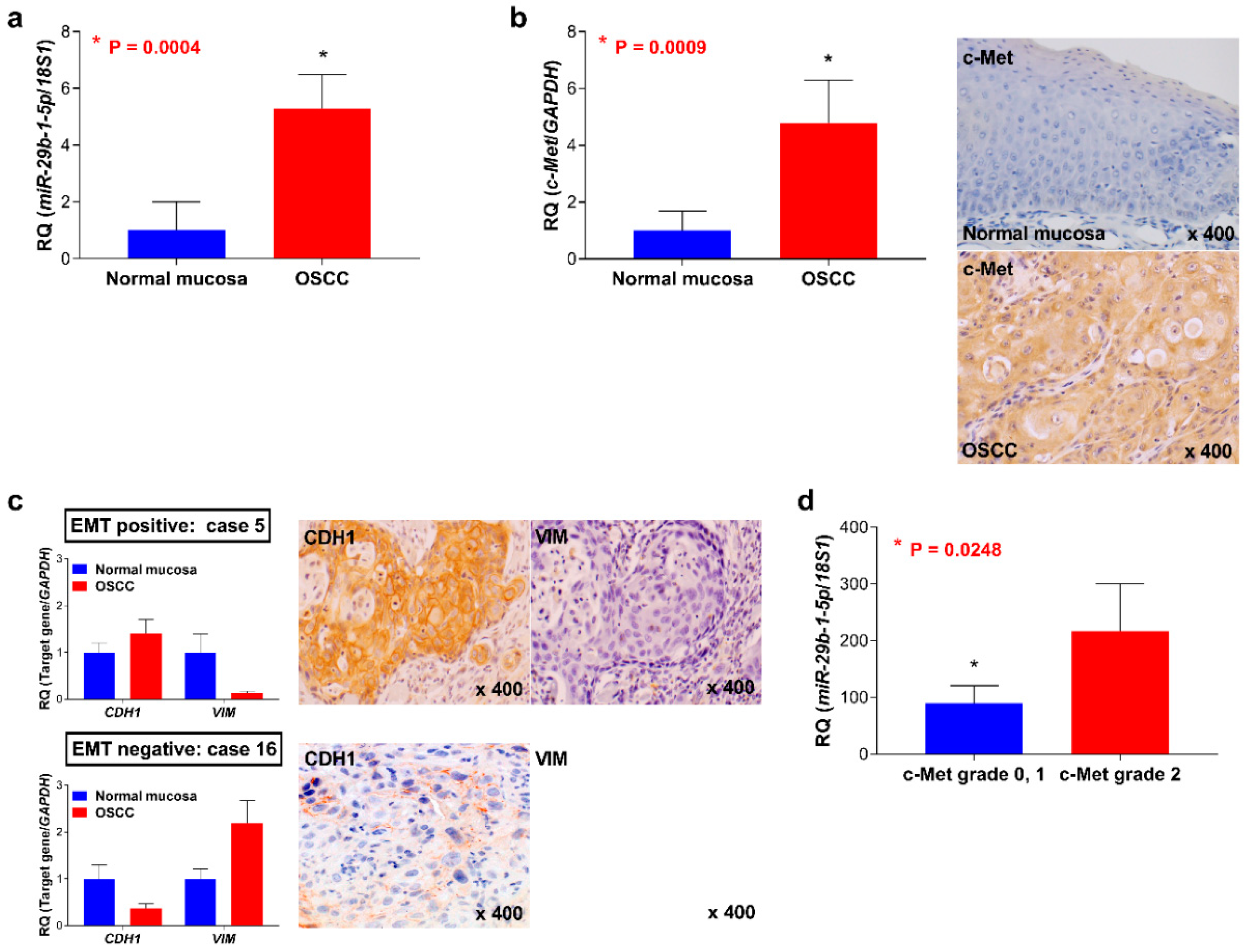
3.1. Expression of miR-29b-1-5p and c-Met and EMT Status of Patients with OSCC
3.2. Association between the Expression of miR-29b-1-5p and Clinicopathological Characteristics of Tissue Samples of Patients with OSCC
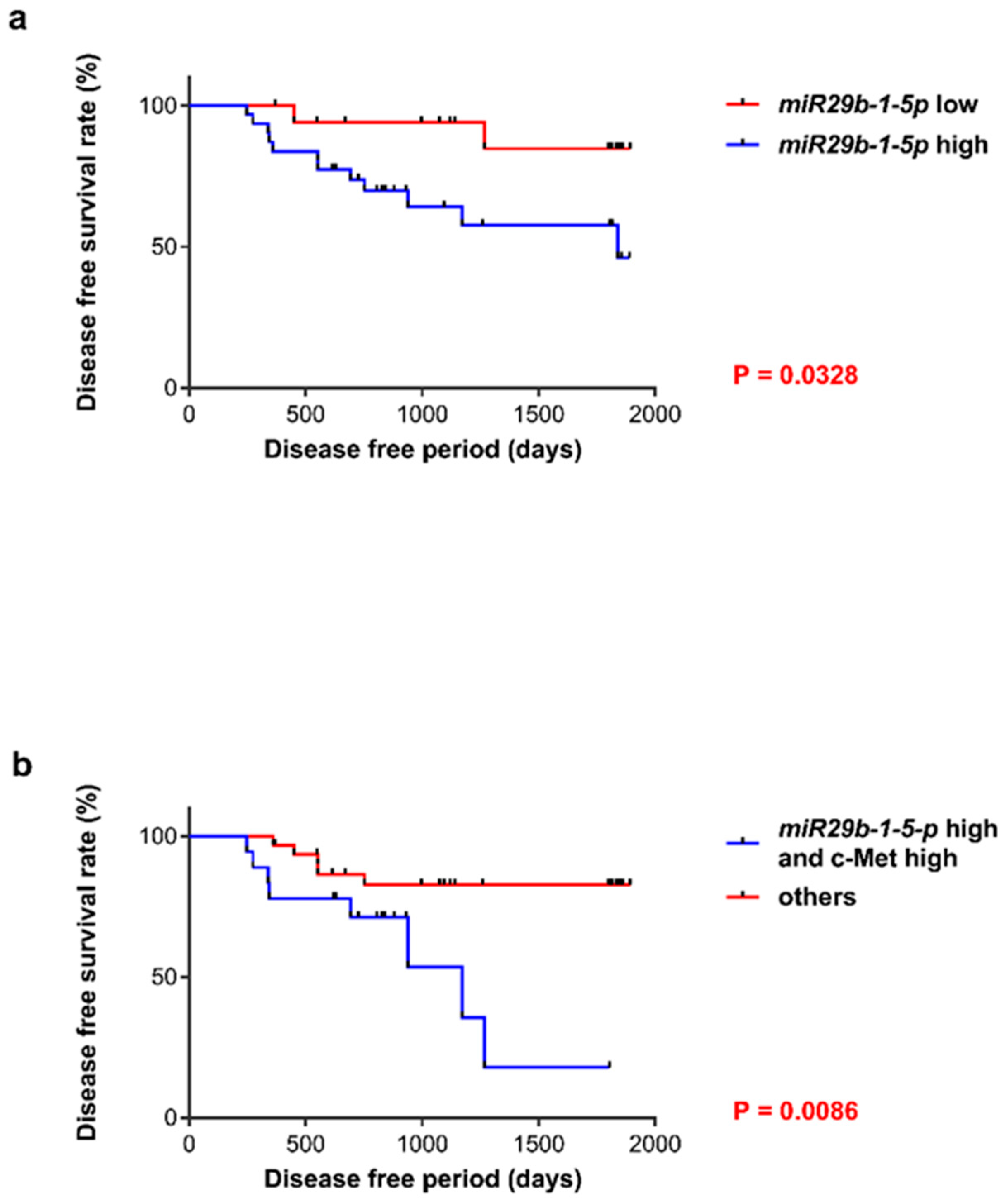
3.3. Prognosis of Patients with OSCC
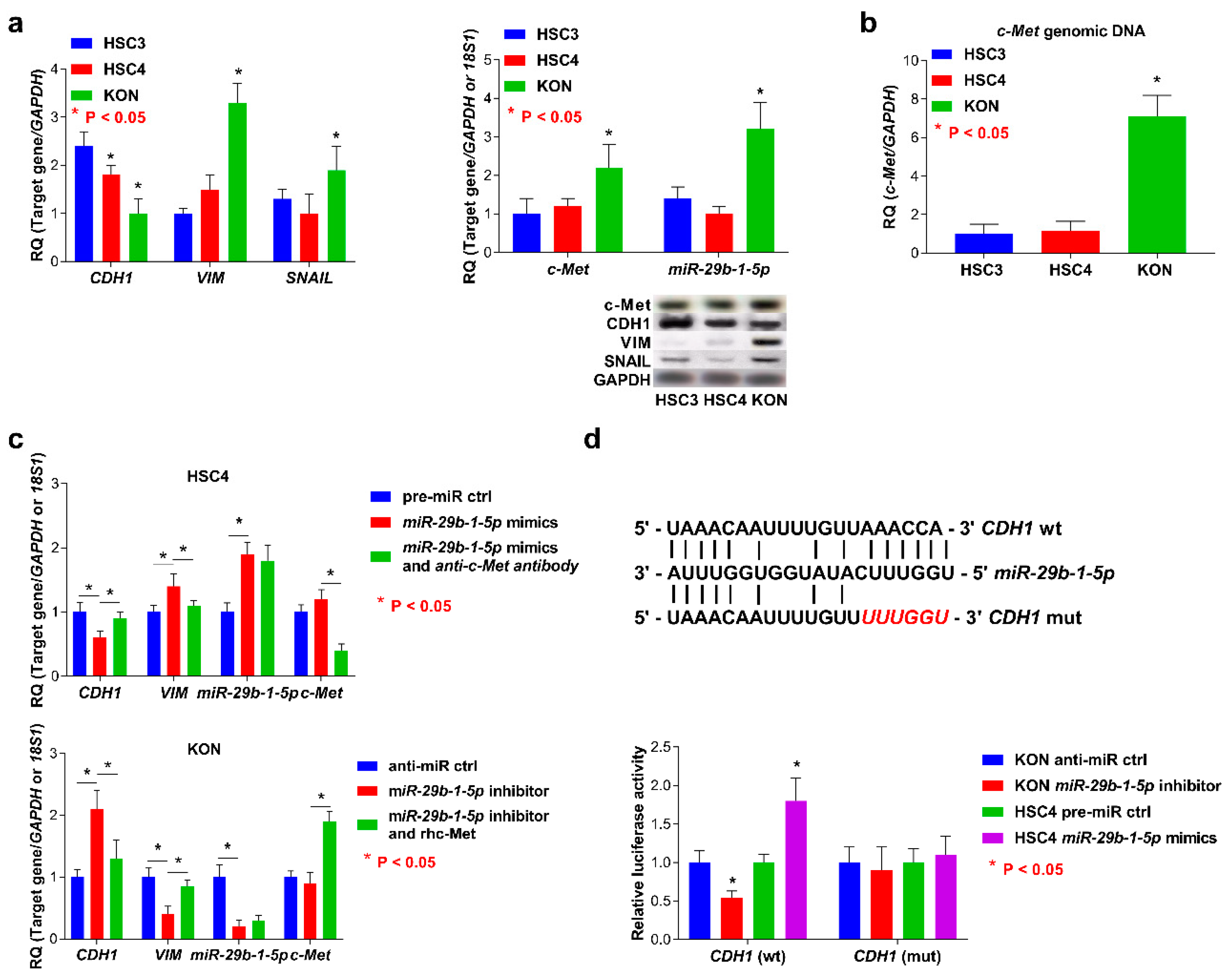
3.4. Expression of miR-29b-1-5p and c-Met and EMT Status in OSCC cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shield, K.D.; Ferlay, J.; Jemal, A.; Sankaranarayanan, R.; Chaturvedi, A.K.; Bray, F.; Soerjomataram, I. The global incidence of lip, oral cavity, and pharyngeal cancers by subsite in 2012. CA Cancer J. Clin. 2017, 67, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Sasahira, T.; Bosserhoff, A.K.; Kirita, T. The importance of melanoma inhibitory activity gene family in the tumor progression of oral cancer. Pathol. Int. 2018, 68, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Sasahira, T.; Kurihara, M.; Bhawal, U.K.; Ueda, N.; Shimomoto, T.; Yamamoto, K.; Kirita, T.; Kuniyasu, H. Downregulation of miR-126 induces angiogenesis and lymphangiogenesis by activation of VEGF-A in oral cancer. Br. J. Cancer 2012, 107, 700–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manasa, V.G.; Kannan, S. Impact of microRNA dynamics on cancer hallmarks: An oral cancer scenario. Tumour Biol. 2017, 39, 1010428317695920. [Google Scholar] [CrossRef] [PubMed]
- Sasahira, T.; Kirita, T. Hallmarks of cancer-related newly prognostic factors of oral squamous cell carcinoma. Int. J. Mol. Sci. 2018, 19, 2413. [Google Scholar] [CrossRef] [PubMed]
- Koshizuka, K.; Kikkawa, N.; Hanazawa, T.; Yamada, Y.; Okato, A.; Arai, T.; Katada, K.; Okamoto, Y.; Seki, N. Inhibition of integrin beta1-mediated oncogenic signalling by the antitumor microRNA-29 family in head and neck squamous cell carcinoma. Oncotarget 2018, 9, 3663–3676. [Google Scholar] [CrossRef] [PubMed]
- Milevskiy, M.J.G.; Sandhu, G.K.; Wronski, A.; Korbie, D.; Brewster, B.L.; Shewan, A.; Edwards, S.L.; French, J.D.; Brown, M.A. MiR-29b-1-5p is altered in BRCA1 mutant tumours and is a biomarker in basal-like breast cancer. Oncotarget 2018, 9, 33577–33588. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Guo, Q.; Fu, F.J.; Wang, Z.; Yin, Z.; Wei, Y.B.; Yang, J.R. The role of miR-29b in cancer: Regulation, function, and signaling. Onco Targets Ther. 2015, 8, 539–548. [Google Scholar] [PubMed]
- Yamaguchi, N.; Osaki, M.; Onuma, K.; Yumioka, T.; Iwamoto, H.; Sejima, T.; Kugoh, H.; Takenaka, A.; Okada, F. Identification of MicroRNAs Involved in Resistance to Sunitinib in Renal Cell Carcinoma Cells. Anticancer Res. 2017, 37, 2985–2992. [Google Scholar] [PubMed]
- Drago-Ferrante, R.; Pentimalli, F.; Carlisi, D.; De Blasio, A.; Saliba, C.; Baldacchino, S.; Degaetano, J.; Debono, J.; Caruana-Dingli, G.; Grech, G.; et al. Suppressive role exerted by microRNA-29b-1-5p in triple negative breast cancer through SPIN1 regulation. Oncotarget 2017, 8, 28939–28958. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Zhang, Q.; Cheng, W.; Zhang, Z.; Wang, J.; Ge, J. Effect of miR-29b-1* and miR-29c knockdown on cell growth of the bladder cancer cell line T24. J. Int. Med. Res. 2013, 41, 1803–1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.J.; Hwang, K.C.; Kim, S.W.; Lee, Y.C. Potential miRNA-target interactions for the screening of gastric carcinoma development in gastric adenoma/dysplasia. Int. J. Med. Sci. 2018, 15, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Lo Muzio, L.; Farina, A.; Rubini, C.; Coccia, E.; Capogreco, M.; Colella, G.; Leonardi, R.; Campisi, G.; Carinci, F. Effect of c-Met expression on survival in head and neck squamous cell carcinoma. Tumour Biol. 2006, 27, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, I.; Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Matsushita, R.; Kurozumi, A.; Kato, M.; Okato, A.; Okamoto, Y.; Seki, N. The tumor-suppressive microRNA-23b/27b cluster regulates the MET oncogene in oral squamous cell carcinoma. Int. J. Oncol. 2016, 49, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Liu, Q.; Ye, D.; Ye, K.; Yang, Z.; Li, D. Role of c-Met in the progression of human oral squamous cell carcinoma and its potential as a therapeutic target. Oncol. Rep. 2018, 39, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Tanahashi, T.; Osada, S.; Yamada, A.; Kato, J.; Yawata, K.; Mori, R.; Imai, H.; Sasaki, Y.; Saito, S.; Tanaka, Y.; et al. Extracellular signal-regulated kinase and Akt activation play a critical role in the process of hepatocyte growth factor-induced epithelial-mesenchymal transition. Int. J. Oncol. 2013, 42, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, R.; Tsuda, M.; Wang, L.; Maishi, N.; Abe, T.; Kimura, T.; Tanino, M.; Nishihara, H.; Hida, K.; Ohba, Y.; et al. Adaptor protein CRK induces epithelial-mesenchymal transition and metastasis of bladder cancer cells through HGF/c-Met feedback loop. Cancer Sci. 2015, 106, 709–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wang, X.; Li, J.; Meng, S.; Liang, Z.; Xu, X.; Zhu, Y.; Li, S.; Wu, J.; Xu, M.; et al. c-Met, CREB1 and EGFR are involved in miR-493-5p inhibition of EMT via AKT/GSK-3beta/Snail signaling in prostate cancer. Oncotarget 2017, 8, 82303–82313. [Google Scholar] [PubMed]
- Busch, A.; Bauer, L.; Wardelmann, E.; Rudack, C.; Grunewald, I.; Stenner, M. Prognostic relevance of epithelial-mesenchymal transition and proliferation in surgically treated primary parotid gland cancer. J. Clin. Pathol. 2017, 70, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Allred, D.C.; Harvey, J.M.; Berardo, M.; Clark, G.M. Prognostic and predictive factors in breast cancer by immunohistochemical analysis. Mod. Pathol. 1998, 11, 155–168. [Google Scholar] [PubMed]
- Kurihara-Shimomura, M.; Sasahira, T.; Nakashima, C.; Kuniyasu, H.; Shimomura, H.; Kirita, T. The multifarious functions of pyruvate kinase M2 in oral cancer cells. Int. J. Mol. Sci. 2018, 19, 2907. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Nakajima, G.; Gavin, E.; Morris, C.G.; Kudo, K.; Hayashi, K.; Ju, J. Systematic analysis of microRNA expression of RNA extracted from fresh frozen and formalin-fixed paraffin-embedded samples. Rna 2007, 13, 1668–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Ouyang, H.; Wang, Z.; Chen, B.; Nie, Q. A Novel Circular RNA Generated by FGFR2 Gene Promotes Myoblast Proliferation and Differentiation by Sponging miR-133a-5p and miR-29b-1-5p. Cells 2018, 7, 199. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.N.; Deng, Y.T.; Tang, J.Y.; Cheng, S.J.; Chen, S.T.; Li, Y.J.; Wu, T.S.; Yang, M.H.; Lin, B.R.; Kuo, M.Y.; et al. MicroRNA-29b regulates migration in oral squamous cell carcinoma and its clinical significance. Oral Oncol. 2015, 51, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or tumor suppressor? The duplicity of microRNAs in cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef] [PubMed]
- Kuniyasu, H.; Yasui, W.; Kitadai, Y.; Yokozaki, H.; Ito, H.; Tahara, E. Frequent amplification of the c-met gene in scirrhous type stomach cancer. Biochem. Biophys. Res. Commun. 1992, 189, 227–232. [Google Scholar] [CrossRef]
- Kuniyasu, H.; Yasui, W.; Yokozaki, H.; Kitadai, Y.; Tahara, E. Aberrant expression of c-met mRNA in human gastric carcinomas. Int. J. Cancer 1993, 55, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Kuniyasu, H.; Yasui, W.; Yokozaki, H.; Akagi, M.; Akama, Y.; Kitahara, K.; Fujii, K.; Tahara, E. Frequent loss of heterozygosity of the long arm of chromosome 7 is closely associated with progression of human gastric carcinomas. Int. J. Cancer 1994, 59, 597–600. [Google Scholar] [CrossRef] [PubMed]
- Domingues, C.; Serambeque, B.P.; Laranjo Candido, M.S.; Marto, C.M.M.; Veiga, F.J.B.; Sarmento Antunes Cruz Ribeiro, A.B.; Figueiras, A.R.R.; Botelho, M.F.R.; Dourado, M. Epithelial-mesenchymal transition and microRNAs: Challenges and future perspectives in oral cancer. Head Neck 2018, 40, 2304–2313. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Hsu, W.L.; Tseng, Y.J.; Liu, D.W.; Weng, C.F. Involvement of DNMT 3B promotes epithelial-mesenchymal transition and gene expression profile of invasive head and neck squamous cell carcinomas cell lines. BMC Cancer 2016, 16, 431. [Google Scholar]
- Reuter, C.W.; Morgan, M.A.; Eckardt, A. Targeting EGF-receptor-signalling in squamous cell carcinomas of the head and neck. Br. J. Cancer 2007, 96, 408–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tringale, K.R.; Carroll, K.T.; Zakeri, K.; Sacco, A.G.; Barnachea, L.; Murphy, J.D. Cost-effectiveness analysis of nivolumab for treatment of platinum-resistant recurrent or metastatic squamous cell carcinoma of the head and neck. J. Natl. Cancer Inst. 2018, 110, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, H.; Sasahira, T.; Nakashima, C.; Shimomura-Kurihara, M.; Kirita, T. Downregulation of DHRS9 is associated with poor prognosis in oral squamous cell carcinoma. Pathology 2018, 50, 642–647. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameters | miR-29b-1-5p | c-Met | ||
|---|---|---|---|---|
| Low | High | Negative | Positive | |
| Gender | ||||
| Male | 12 | 10 | 10 | 10 |
| Female | 10 | 19 | 12 | 17 |
| p value | 0.7679 | 0.5741 | ||
| Age | ||||
| <65 | 8 | 10 | 7 | 11 |
| >65 | 10 | 21 | 15 | 16 |
| p value | 0.5401 | 0.5647 | ||
| Site | ||||
| Tongue | 9 | 21 | 16 | 14 |
| Other | 9 | 10 | 6 | 13 |
| p value | 0.2416 | 0.1551 | ||
| Alcohol intake | ||||
| Habitual drinking | 5 | 15 | 4 | 7 |
| Social drinking | 5 | 7 | 10 | 10 |
| No drinking | 8 | 9 | 8 | 10 |
| p value | 0.3516 | 0.7650 | ||
| Smoking habit | ||||
| Yes | 6 | 8 | 9 | 8 |
| No | 12 | 23 | 13 | 19 |
| p value | 0.7442 | 0.5480 | ||
| Histological | ||||
| Differentiation * | ||||
| Well | 15 | 12 | 13 | 14 |
| Mod, Por | 3 | 19 | 9 | 13 |
| p value | 0.0031 | 0.7738 | ||
| T classification | ||||
| T1–T2 | 15 | 22 | 18 | 19 |
| T3–T4 | 3 | 9 | 4 | 8 |
| p value | 0.4942 | 0.5072 | ||
| Clinical stage | ||||
| I, II | 13 | 16 | 14 | 15 |
| III, IV | 5 | 15 | 8 | 12 |
| p value | 0.2298 | 0.7707 | ||
| Nodal metastasis | ||||
| Negative | 15 | 19 | 16 | 18 |
| Positive | 3 | 12 | 6 | 9 |
| p value | 0.1975 | 0.7597 | ||
| EMT status | ||||
| Negative | 17 | 13 | 17 | 13 |
| Positive | 1 | 18 | 5 | 14 |
| p value | 0.0002 | 0.0453 | ||
| c-Met expression | ||||
| Negative | 9 | 13 | ||
| Positive | 9 | 18 | ||
| p value | 0.7665 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurihara-Shimomura, M.; Sasahira, T.; Shimomura, H.; Nakashima, C.; Kirita, T. The Oncogenic Activity of miR-29b-1-5p Induces the Epithelial-Mesenchymal Transition in Oral Squamous Cell Carcinoma. J. Clin. Med. 2019, 8, 273. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8020273
Kurihara-Shimomura M, Sasahira T, Shimomura H, Nakashima C, Kirita T. The Oncogenic Activity of miR-29b-1-5p Induces the Epithelial-Mesenchymal Transition in Oral Squamous Cell Carcinoma. Journal of Clinical Medicine. 2019; 8(2):273. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8020273
Chicago/Turabian StyleKurihara-Shimomura, Miyako, Tomonori Sasahira, Hiroyuki Shimomura, Chie Nakashima, and Tadaaki Kirita. 2019. "The Oncogenic Activity of miR-29b-1-5p Induces the Epithelial-Mesenchymal Transition in Oral Squamous Cell Carcinoma" Journal of Clinical Medicine 8, no. 2: 273. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8020273