Thyroid Hormones and Spermatozoa: In Vitro Effects on Sperm Mitochondria, Viability and DNA Integrity

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Selection
2.2. Sperm Analysis and Preparation
- Sperm progressive motility.
- Sperm recovery after capacitation by swim-up technique using the Biggers, Whitten, and Whittingham medium (BWW) with capacitating properties. Spermatozoa were then recovered from the supernatant according to their capacity to migrate from the bottom of the test tube to the surface.
- Bio-functional sperm parameters by flow cytometry: Mitochondrial membrane potential (MMP), vitality, early apoptosis, late apoptosis, necrosis, chromatin compactness, DNA fragmentation and lipid peroxidation (LP).
2.3. Flow Cytometric Analysis
2.4. Evaluation of Sperm Apoptosis/Vitality
2.5. Evaluation of the Mitochondrial Membrane Potential
- spermatozoa with high mitochondrial membrane potential (H-MMP)
- spermatozoa with low mitochondrial membrane potential (L-MMP).
2.6. Assessment of DNA Fragmentation
2.7. Degree of Chromatin Compactness Assessment
2.8. Sperm Membrane Lipid Peroxidation Evaluation
2.9. Statistical Analysis
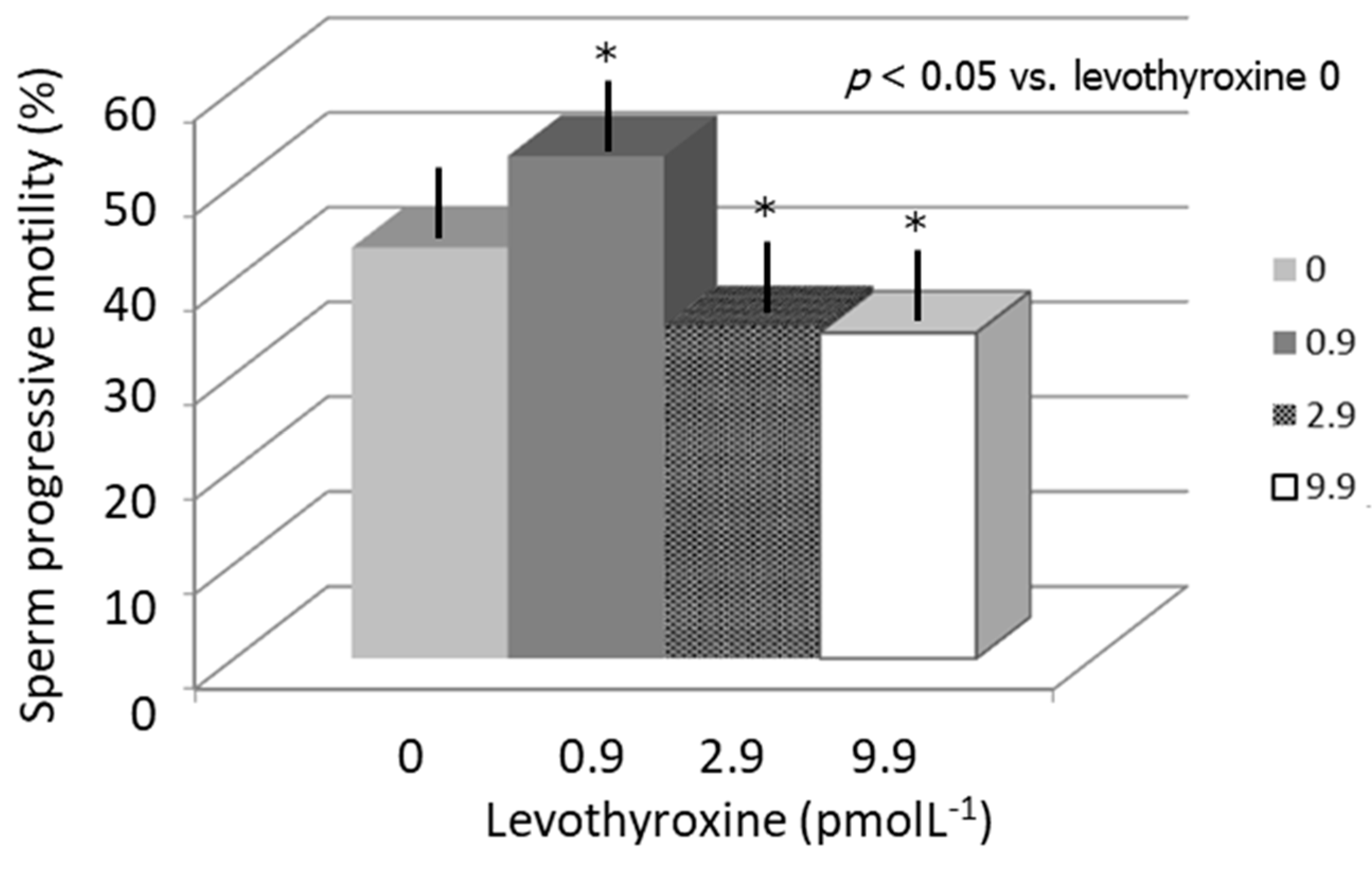
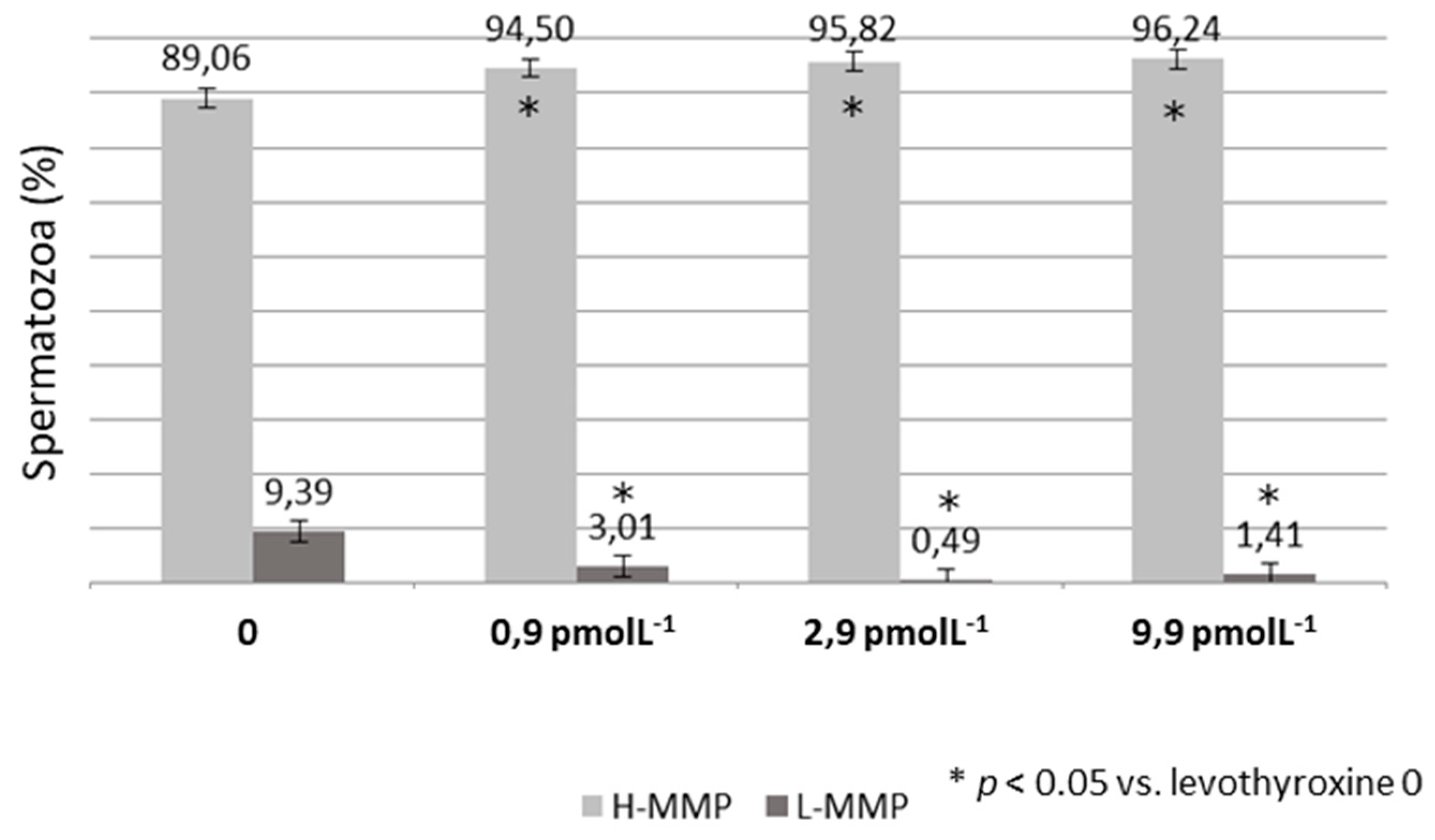
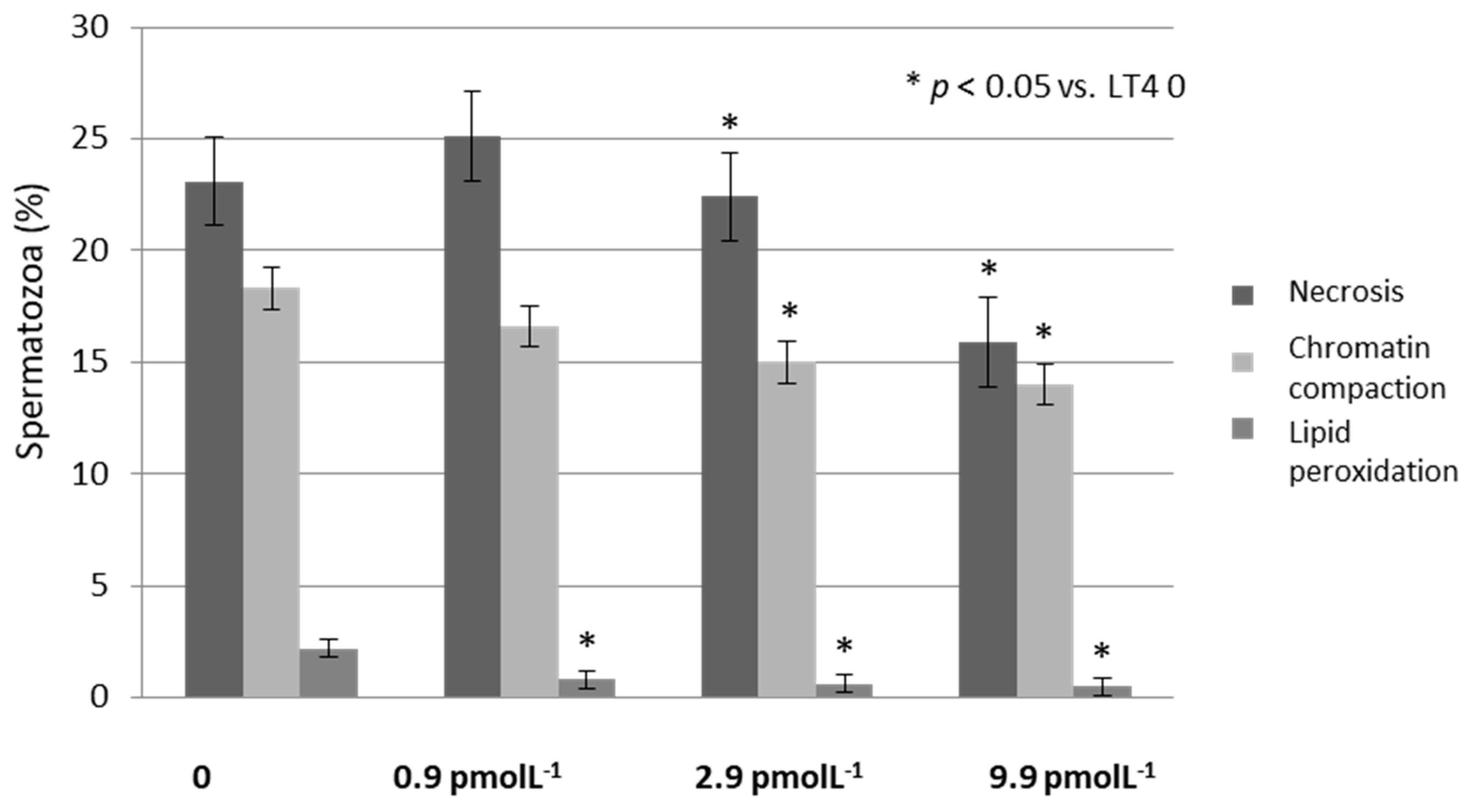
3. Results
4. Discussion
5. Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- Wagner, M.S.; Wajner, S.M.; Maia, A.L. Is there a role for thyroid hormone on spermatogenesis? Microsc. Res. Tech. 2009, 72, 796–808. [Google Scholar] [CrossRef]
- La Vignera, S.; Vita, R.; Condorelli, R.A.; Mongioì, L.M.; Presti, S.; Benvenga, S.; Calogero, A.E. Impact of thyroid disease on testicular function. Endocrine 2017, 58, 397–407. [Google Scholar] [CrossRef]
- Dittrich, R.; Beckmann, M.W.; Oppelt, P.G.; Hoffman, I.; Lotz, I.; Kuwert, T.; Mueller, A. Thyroid hormone receptors and reproduction. J. Reprod. Immunol. 2011, 90, 58–66. [Google Scholar] [CrossRef]
- Romano, R.M.; Gomes, S.N.; Cardoso, N.C.; Schiessl, L.; Romano, M.A.; Oliveira, C.A. New insights for male infertility revealed by alterations in spermatic function and differential testicular expression of thyroid-related genes. Endocrine 2017, 55, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Abalovich, M.; Levalle, O.; Hermes, R.; Scaglia, H.; Aranda, C.; Zylbersztein, C.; Oneto, A.; Aquilano, D.; Gutierrez, S. Hypothalamic-pituitary-testicular axis and seminal parameters in hyperthyroid males. Thyroid 1999, 9, 857–863. [Google Scholar] [CrossRef]
- Hudson, R.W.; Edwards, A.L. Testicular function in hyperthyroidism. J. Androl. 1992, 13, 117–124. [Google Scholar] [PubMed]
- Krassas, G.E.; Poppe, K.; Glinoer, D. Thyroid function and human reproductive health. Endocrine Rev. 2010, 31, 702–755. [Google Scholar] [CrossRef]
- Choudhury, S.; Chainy, G.B.; Mishro, M.M. Experimentally induced hypo- and hyper-thyroidism influence on the antioxidant defence system in adult rat testis. Andrologia 2003, 35, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, D.K.; Roy, A.; Bhanja, S.; Chainy, G.B. Hypothyroidism impairs antioxidant defence system and testicular physiology during development and maturation. Gen. Comp. Endocrinol. 2008, 156, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Buzzard, J.J.; Morrison, J.R.; O’Bryan, M.K.; Song, Q.; Wreford, N.G. Developmental expression of thyroid hormone receptors in the rat testis. Biol. Reprod. 2000, 62, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Jannini, E.A.; Crescenzi, A.; Rucci, N.; Screponi, E.; Carosa, E.; De Matteis, A.; Macchia, E.; D’Amati, G.; D’Armiento, M. Ontogenetic pattern of thyroid hormone receptor expression in the human testis. J. Clin. Endocrinol. Metab. 2000, 85, 3453–3457. [Google Scholar] [CrossRef] [PubMed]
- Carosa, E.; Lenzi, A.; Jannini, E.A. Thyroid hormone receptors and ligands, tissue distribution and sexual behavior. Mol. Cell Endocrinol. 2017, 30, 1–11. [Google Scholar] [CrossRef]
- La Vignera, S.; Vita, R. Thyroid dysfunction and semen quality. Int. J. Immunopathol. Pharmacol. 2018. [Google Scholar] [CrossRef]
- Fumel, B.; Froment, P.; Holzenberger, M.; Livera, G.; Monget, P.; Fouchecourt, S. Expression of domnant-negative thyroid hormone receptor alpha1 in Leydig and Sertoli cells demonstrates no additional defect compared with expression in Sertoli cells only. PLoS ONE 2015, 10, e01119392. [Google Scholar] [CrossRef] [PubMed]
- Xian, H.; Wang, F.; Teng, W.; Yang, D.; Zhang, M. Thyroid hormone induce a p53-dependent DNA damage through PI3K/Akt activation in spem. Gene 2017, 615, 1–7. [Google Scholar] [CrossRef]
- Jacob, T.N.; Pandey, J.P.; Raghuveer, K.; Sreenivasulu, G.; Gupta, A.D.; Yoshikuni, M.; Jagota, A.; Senthilkumaran, B. Thyroxine-induced alterations in the testis and seminal vesicles of air-breathing catfish, Clariasgariepinus. Fish Physiol. Biochem. 2005, 31, 271–274. [Google Scholar] [CrossRef]
- Dobrzynska, M.M.; Baumgartner, A.; Anderson, D. Antioxidant modulate thyroid hormone and noradrenaline induced DNA damage in human sperm. Mutagenesis 2004, 19, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Meeker, J.D.; Singh, N.P.; Hauser, R. Serum concentrations of estradiol and free T4 are inversely correlated with sperm DNA damage in men from an infertility clinic. J. Androl. 2008, 29, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.Y.; Umezu, M.; Sato, E. Characteristics of infertility and the improvement of fertility by thyroxine treatment in adult male hypothyroid rdw rats. Biol. Reprod. 2000, 63, 1637–1641. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Paoli, D.; Gallo, M.; Rizzo, F.; Baldi, E.; Francavilla, S.; Lenzi, A.; Lombardo, F.; Gandini, L. Mitochondrial membrane potential profile and its correlation with increasing sperm motility. Fertil. Steril. 2011, 95, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Ferramosca, A.; Provenzano, S.; Coppola, L.; Zara, V. Mitochondrial respiratory efficiency is positively correlated with human sperm motility. Urology 2012, 79, 809–814. [Google Scholar] [CrossRef]
- Moscatelli, N.; Spagnolo, B.; Pisanello, M.; Lemma, E.D.; De Vittorio, M.; Zara, V.; Pisanello, F.; Ferramosca, A. Single-cell-based evaluation of sperm progressive motility via fluorescent assessment of mitochondria membrane potential. Sci. Rep. 2017, 7, 17931. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Patients (n = 15) | |
|---|---|
| Concentration (106 mL−1) | 42.80 ± 12.0 |
| Total count (106 ejaculate−1) | 107.00 ± 30 |
| Progressive motility (%) | 39.80 ± 3.3 |
| Total motility (%) | 77.90 ± 9.8 |
| Normal form (%) | 8.30 ± 0.8 |
| Leukocytes (106 mL−1) | 0.70 ± 0.2 |
| Seminal FT4 (pmol L−1) | 3.15 ± 0.7 |
| Age | 31.2 ± 6.4 |
| BMI (kg m−2) | 24.2 ± 1.3 |
| Waist circumference (cm) | 94.0 ± 2.4 |
| TSH (μUI m L−1) | 2.1 ± 0.5 |
| FT4 (pmol L−1) | 10.2 ± 0.9 |
| FT3 (pmol L−1) | 4.7 ± 0.8 |
| FSH (UI L−1) | 2.3 ± 0.1 |
| LH (UI L−1) | 2.4 ± 0.2 |
| Total testosterone (ng mL−1) | 6.1 ± 1.1 |
| Sperm Parameters | LT4 Concentrations | |||
|---|---|---|---|---|
| 0 | 0.9 pmol L−1 | 2.9 pmol L−1 | 9.9 pmol L−1 | |
| Alive | 65.2 ± 4.6 | 53.6 ± 8.8 | 57.3 ± 6.2 | 54.0 ± 9.3 |
| Early Apoptosis | 2.0 ± 0.8 | 1.5 ± 0.5 | 1.6 ± 0.7 | 1.8 ± 0.5 |
| Late apoptosis | 18.6 ± 8.6 | 19.9 ± 8.8 | 15.2 ± 6.4 | 23.9 ± 7.8 |
| DNA fragmentation | 3.4 ± 0.5 | 3.8 ± 0.6 | 4.1 ± 0.7 | 3.9 ± 0.7 |
| Sperm recovery by swim-up | 18.7 ± 8.8 | 21.3 ± 10.0 | 14.9 ± 7.7 | 11.9 ± 10.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Condorelli, R.A.; La Vignera, S.; Mongioì, L.M.; Alamo, A.; Giacone, F.; Cannarella, R.; Calogero, A.E. Thyroid Hormones and Spermatozoa: In Vitro Effects on Sperm Mitochondria, Viability and DNA Integrity. J. Clin. Med. 2019, 8, 756. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8050756
Condorelli RA, La Vignera S, Mongioì LM, Alamo A, Giacone F, Cannarella R, Calogero AE. Thyroid Hormones and Spermatozoa: In Vitro Effects on Sperm Mitochondria, Viability and DNA Integrity. Journal of Clinical Medicine. 2019; 8(5):756. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8050756
Chicago/Turabian StyleCondorelli, Rosita A., Sandro La Vignera, Laura M. Mongioì, Angela Alamo, Filippo Giacone, Rossella Cannarella, and Aldo E. Calogero. 2019. "Thyroid Hormones and Spermatozoa: In Vitro Effects on Sperm Mitochondria, Viability and DNA Integrity" Journal of Clinical Medicine 8, no. 5: 756. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8050756