Uncoupling Traditional Functionalities of Metastasis: The Parting of Ways with Real-Time Assays


National Centre for Cell Science, Savitribai Phule Pune University, Ganeshkhind, Pune 411007, India
*
Author to whom correspondence should be addressed.
J. Clin. Med. 2019, 8(7), 941; https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8070941
Submission received: 29 April 2019
/
Revised: 31 May 2019
/
Accepted: 4 June 2019
/
Published: 28 June 2019
(This article belongs to the Special Issue Epithelial-Mesenchymal Plasticity in Cancer Metastasis: Molecular Reprogramming, Cellular Adaptation, and Clinical Implications)
{kind=link}
{kind=link}
{kind=link}
Abstract
:The experimental evaluation of metastasis overly focuses on the gain of migratory and invasive properties, while disregarding the contributions of cellular plasticity, extra-cellular matrix heterogeneity, niche interactions, and tissue architecture. Traditional cell-based assays often restrict the inclusion of these processes and warrant the implementation of approaches that provide an enhanced spatiotemporal resolution of the metastatic cascade. Time lapse imaging represents such an underutilized approach in cancer biology, especially in the context of disease progression. The inclusion of time lapse microscopy and microfluidic devices in routine assays has recently discerned several nuances of the metastatic cascade. Our review emphasizes that a complete comprehension of metastasis in view of evolving ideologies necessitates (i) the use of appropriate, context-specific assays and understanding their inherent limitations; (ii) cautious derivation of inferences to avoid erroneous/overestimated clinical extrapolations; (iii) corroboration between multiple assay outputs to gauge metastatic potential; and (iv) the development of protocols with improved in situ implications. We further believe that the adoption of improved quantitative approaches in these assays can generate predictive algorithms that may expedite therapeutic strategies targeting metastasis via the development of disease relevant model systems. Such approaches could potentiate the restructuring of the cancer metastasis paradigm through an emphasis on the development of next-generation real-time assays.
1. Introduction
Metastasis generates a systemic disease driven by the concerted alliance of tumor cell dissociation, physical translocation, and distant colonization. The dissociation of cells from the primary tumor initiates the metastatic cascade and generates distinct entities defined by severance dynamics of cell adhesion complexes. Further, acquisition of migratory/invasive properties by tumor cells facilitates their entry into circulation (or lymphatic system), wherein they evade host immune responses and endure extrinsic pressures followed by extravasation at secondary site(s). Tumor cell dormancy and secondary site remodeling then define the latency of colonization, which eventually establishes metastatic lesions [1,2,3]. An intricate interplay of molecular networks drives these programs and contributes to the efficacy of tumor progression [1,2,3,4]. Phenotypic switches induced by epithelial–mesenchymal transitions (EMT) are widely studied, and are deemed crucial for successful metastasis. Alternatively, recent studies highlight a collective mode of dissemination, wherein the retention of several epithelial properties is well documented [1,2,3,4]. These processes are further influenced by numerous physiological parameters, including the tissue stroma, architecture, and extra-cellular matrix (ECM) composition, in order to facilitate the successful establishment of a secondary disease [1,2,3,4]. Thus, collaboration amongst diverse functionalities outlines a complex blueprint of the process, wherein distinct modes of tumor cell dissemination can influence disease aggressiveness and therapeutic response.
Routine analyses in cancer metastasis studies inadvertently employ endpoint or “snapshot” assays that widely focus on the physical translocation of cancer cells. Functional read-outs tend to simplify various aspects of metastasis, and often neglect the role of tumor heterogeneity in disease progression. Furthermore, current notions extensively associate dissemination with a mesenchymal phenotype, and disregard the contributions of the alternate processes that are often unnoticed by existing experimental systems [3,5,6]. Detailed examination of the metastatic cascade thus necessitates the integration of the recent conceptual and technical advances for the development of informative assay protocols. In this article, we present a reassessment of the functional read-outs routinely employed in metastasis-associated studies, and accentuate the application of high-resolution imaging approaches to derive relevant patho-physiological conclusions.
2. Snapshot Assays for Metastasis Assessment
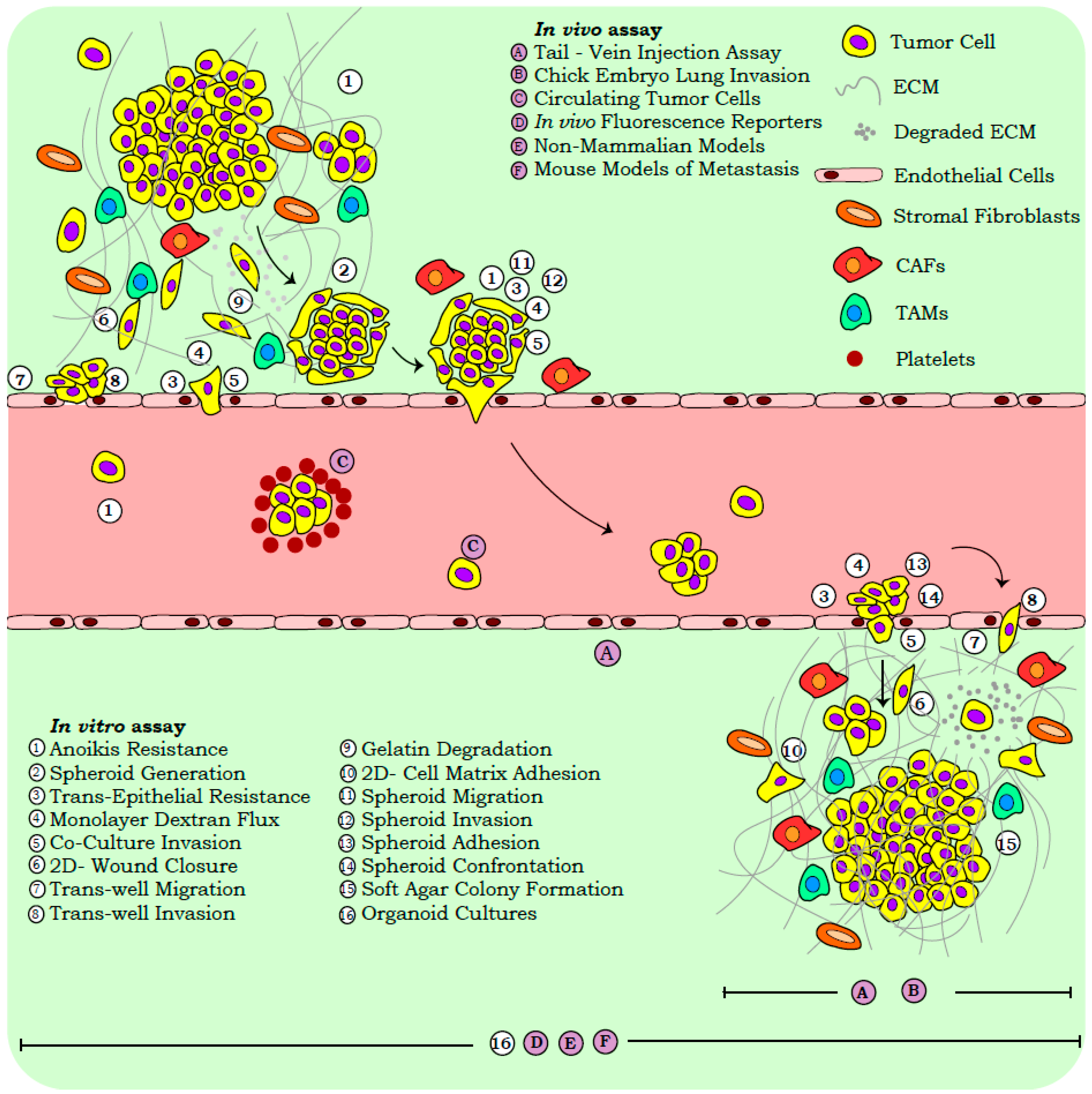
Reliance on snapshot assays in cancer research is accounted for by several advantages, including the ease of execution, reproducibility, and applicability in high throughput screens. In vitro functional read-outs provide a preliminary assessment of the metastatic capabilities, while the variables contributing to tumor heterogeneity viz., micro-environmental milieu, tissue specific metabolic gradients, and systemic architecture, are often assessed with in vivo models. While our review focusses on the widely employed functional assays corresponding to three distinct stages of metastasis, viz., primary dissociation, physical translocation, and colonization, we briefly discuss the molecular approaches routinely used in cancer biology (Figure 1).
2.1. Molecular Assays
The functional assessment of metastasis is often correlated with the molecular signatures derived from tumor cells or cell line models. Primary profiling studies employ a wide range of markers identified across the metastatic cascade, which include cell junction and cytoskeletal components, transcription factors (TFs), secretory enzymes, and cell surface receptors [7]. Molecular profiles, averaged from a cell population, can often misrepresent disease heterogeneity, as affirmed by the reports on single cell characterization, besides over-emphasizing the role of EMT during metastasis [8,9,10]. Microscopy studies further associate the sub-cellular localization of several phenotype associated markers and TFs with distinct cellular functions [11,12]. Importantly, recent reports associating altered marker sub-cellular localization with pathological conditions necessitate the inclusion of this parameter in clinical assessments [13,14,15]. Furthermore, mechanistic studies on cell state maintenance employ fluorescence or enzyme (luciferase, β-galactosidase, and chloramphenicol acetyltransferase) assisted reporter systems for quantifying gene regulation [16,17]. Apart from the static molecular profiles, cytoskeletal, vesicular, and membrane dynamics, as captured by microscopy, offer deeper insight into the alterations of the cell shape and function [18,19,20].
Molecular assays, however, rely on markers that often exhibit extensive disparities across model systems, and are subject to cellular context-specific modulation [21,22,23]. E-cadherin expression and membrane localization, often gauged in clinical specimens by immuno-histochemical scoring, were exclusively associated with the lack of metastasis [24,25,26]; however, the detection of this adherens junction molecule in collectively metastasizing cells challenges its inverse correlation with dissemination [27,28]. Recently, E-cadherin negative cells have also been reported to exhibit collective migration by virtue of CD44 mediated cell-cell adhesion in invasive breast lobular carcinoma [29]. Such discrepancies arise from tissue-specific plasticity programs that are influenced by the local microenvironment. Similarly, the divergent contribution of regulatory TFs in metastasis has been reported; some examples include the stage specific roles of the EMT-mediating TFs Zeb1 and Zeb2 in pancreatic cancer and melanoma dissemination [30,31]; an EMT-TF circuitry switch in melanoma, wherein Slug–Zeb2 act as tumor suppressors in melanocytes, while Twist1–Zeb1 function towards neoplastic transformation [32]; the tissue-specific expression of the Prrx1 isoforms (Prrx1a and Prrx1b) that govern the distinct phenotypic states in pancreatic and breast cancer progression [33,34]; the co-operative role of Slug and Sox9 in the maintenance of breast epithelium homeostasis [35]; and so on. Thus, assigning relevance to metastases signatures requires an accompanying physiological comprehension of the cellular plasticity, and corroboration with tissue specific molecular profiles, mechanistic approaches, and imaging protocols.
2.2. Functional Assays
The examination of functionalities across the physiological and pathological states is robustly aided by cell-based assays. Routinely employed assays in cancer biology gauge the properties of anoikis resistance, stemness, migration, invasion, and colonization, so as to correlate with clinical observations (Table S1 and Figure 1).
2.2.1. In Vitro Assays
The loss of cell–cell and cell–matrix contacts initiates the anoikis cascade under physiological conditions, whereas resistance to this cell-death pathway in tumor cells permits effective disease progression [36]. The in vitro assessment of “anoikis” relies on the differential uptake of fluorescent dyes or the biochemical conversions of fluorophores by viable versus non-viable cells. Dissociated tumor cells exhibit a tendency to generate organized multi-cellular structures termed “spheroids”, disorganized “cellular aggregates”, or they exist as single cells [37]. Cell aggregates and spheroids exhibit stem-like and anoikis resistance properties, and are associated with disease aggressiveness [38,39].
The functional attributes of cell migration and invasion are also hijacked during tumor progression. Routine assessment of migratory capabilities by the “in vitro wound closure assays” is often enhanced by the fluorescent tagging of cells with reporter proteins or membrane labelling dyes [40,41]. The efficacy of tumor cell intra-/extra-vasation is recapitulated in vitro by trans-well inserts of pre-defined pore sizes. Such assays are employed widely to quantify “cell motility” in response to a chemotactic agent, while the layering of artificial matrix components (such as matrigel, carboxy methyl cellulose, and hydrogels) over these inserts gauge properties of “cell invasion” and “ECM remodeling” [38]. Recent trans-well systems employ micro-electrode coupled inserts that measure the impedance flux in response to cell migration/invasion, with enhanced precision [42]. Tumor cell invasion also entails the biochemical and mechanical modification of ECM components. “ECM degradation assays” measure the enzymatic activity of secreted matrix metalloproteinases (MMPs) by employing fluorescently labelled substrates and quantifying the signal intensity in the vicinity of invasive tumor cells [43]. Additional information pertaining to tumor cell invasion is gained from the “co-culture assays” that recapitulate the in-situ disruption of non-transformed tissue linings often encountered by metastasizing cancer cells [44]. The use of differentially labelled non-transformed and cancer cells improves the resolution of this assay, by enhancing the visualization and quantification of monolayer perturbance and invasion by tumor cells. Monolayer disruption by invading cancer cells is also quantified by the “trans-epithelial resistance (TER)” and “dextran flux assays” that gauge monolayer integrity and permeability, respectively [45,46]. Recent developments in advanced microfluidic and flow cytometry approaches have also facilitated the isolation of “circulating tumor cells (CTCs)”, which are actual proof of an ongoing metastatic cascade [10,47]. Several recent reviews detail, at length, the role of CTCs in tumor biology and translational medicine, and associate them with stem-like properties, immune evasion signatures, and immense phenotypic plasticity [48,49,50]. CTCs also offer excellent diagnostic/prognostic value and specific targeting opportunities [51,52,53]. Hence, their detection, quantification, and analyses are being developed in view of clinical applications.
Cell-substrate adhesion, governed by tissue specific ECM components, represents a critical determinant of metastatic seeding [54,55,56]. The in vitro assessment of “adhesion” employs ECM pre-coated plates to identify the critical molecular players mediating cell–matrix interactions [57]. Separately, anchorage independent growth of tumor cells as gauged by the “soft agar assay” utilizes a three-dimensional (3D) matrix devoid of ECM components, and quantifies the tumor seeding capacity in vitro [58]. Apart from studies pertaining to single cell colonization, spheroids and cell aggregates are also functionally examined for their properties of adhesion, migration, and invasion, which can be extrapolated to the metastatic cascade [38]. Furthermore, the competitive interaction of these entities in suspension is assessed by “spheroid confrontation”; such an assay evaluates the differential invasive capabilities, as well as co-operation between these cell populations in view of metastases seeding [38].
Despite their obvious advantages, cell line models provide limited information on the in situ landscape of a disease, because of a lack of higher order organization conferred by the tissue architecture. “Organoid cultures” represent an in vitro 3D model system reminiscent of the in-situ organization, and provide improved clinical correlations [59]. The sustenance of genetic features from patient samples by organoids make them improved models for studying metastases as opposed to cell lines, which can often accumulate genetic aberrations over multiple passages [60]. Their amenability to in vitro functional assays further permits a high throughput assessment of pathological states. Studies with pancreatic and colorectal cancer organoids, established from clinical specimens, demonstrate the recapitulation of histological features reminiscent of the parental tissue, when injected into immunocompromised mice [61,62]. Interestingly, stage-specific organoids have also captured the evolving heterogeneity and molecular landscapes of tumors, and can improve the efficacy of therapeutic interventions in personalized medicine [63]. Similarly, organoids have been developed from CTCs that exhibit drug-resistance responses similar to the patients, and are amenable to high-throughput screening for the design of personalized therapeutic regimes [64]. Additional details on the advancements in organoid generation and utility in cancer biology are stated elsewhere [64,65,66].
2.2.2. In Vivo Systems
Despite the widespread applicability of in vitro systems, several aspects of metastasis that contribute to its complexity and heterogeneity are comprehended only when examined in vivo. Several non-mammalian systems have been extensively studied so as to comprehend the functionalities associated with the metastatic cascade. Examples include chemotactic migration observed in Dictyostelium discoideum, anchor cell invasion documented during vulva morphogenesis in Caenorrhabditis elegans, collective cell migration of ovarian border cells in Drosophila melanogaster, and the amoeboid migratory phenotype associated with primordial germ cells of Danio rerio. Extensive details on these non-mammalian models have been documented elsewhere [67]. Similarly, the developing chick embryo has been utilized for the assessment of its metastatic capabilities. The traditional chick “chorioallantoic membrane” assay quantifies tumor cell invasion across embryonic layers, and is assessed microscopically [68,69]. Recent “ex ovo embryo-xenograft models”, however, serve as improved visual and quantitative systems for metastatic dissemination and intra-vital imaging [69]. Despite the evolutionary divergence, the conservation of distinct cellular functionalities in these animal models, along with their amenability to genetic screens and live cell imaging, permit the extrapolation of relevant observations to mammalian systems. However, the routine and widespread utility of mouse models in cancer biology as opposed to non-mammalian systems continues, because of emphasis on the clinically relevant settings that are more effectively mimicked by genetically engineered mice.
“Mouse models” recapitulate physiological variables contributing to disease progression, which are absent in vitro and allow for the derivation of clinically relevant outputs. We describe briefly a few mouse models and assays employed in tumor biology; interested readers may refer to other detailed articles [70,71,72]. The metastatic cascade is routinely captured by spontaneous models for metastasis that involves ectopic/orthotopic tumor cell transplantation in immunocompromised mice. Orthotopic models more effectively represent disease progression, as they expose tumor cells to the micro-environmental cues encountered in the tissue of origin. Alternatively, experimental models of metastasis include the inoculation of tumor cells into mice so as to assess the property of distant colonization. Intra-cardiac, intra-peritoneal, intra-splenic, the tail vein, and so on, are known routes of tumor cell injection, and have been documented to govern the tissue specificity of metastatic seeding [72]. Recent advances in bioluminescent imaging also permits the non-invasive detection of metastatic seeding by tagged tumor cells, thus ensuring real-time assessment [17,73]. These mouse models include cell-line generated allograft or xenograft systems that often lack the stromal/immune cell heterogeneity associated with the disease; the development of carcinogen-induced and genetically engineered mouse models has allowed cancer biologists to overcome this drawback. 7,12-Dimethylbenz[a]anthracene and azoxymethane-induced skin squamous cell and colorectal carcinoma mouse models, respectively, have been employed in deciphering the mechanisms of carcinogen mediated disease progression [74,75]. Cell lineage specific disease models, generated by Cre-lox approaches, have identified the molecular and cellular cascades contributing to metastasis and discerned disease associated patterns across stochastic cellular events [76,77]. For the improved recapitulation of breast cancer in mice, the mammary fat pad model system was developed so as to ensure the repopulation and manipulation of the mouse mammary gland with human-derived epithelia [78]. Similarly, patient-derived xenografts (PDXs) have been established, which can recapitulate metastatic and organ homing properties similar to the clinical specimen [79]. Fluorescent tagging of lineage-specific tumor cells and the establishment of confetti mouse models permits the tracing, isolation, and characterization of cell populations, thereby systematically dissecting the events involved in development and disease progression [80,81,82,83]. Reporter tags also facilitate CTC isolation and the detection of low frequency tumor cells at secondary sites during the colonization phase of metastasis [79]. Similarly, inducible reporter systems permit the fine tuning of specific molecular events, besides contributing to the spatiotemporal resolution of gene regulatory networks driving metastasis [75].
Functional assays and model systems thus simplify the comprehension of several mechanisms contributing to metastasis. However, the stromal and immune cell populations that facilitate metastasis are often under-represented in classical assays. These cellular interactions include platelet-coated CTCs, which exhibit enhanced survival and immune evasion; cancer associated fibroblasts (CAFs) or tumor associated macrophages (TAMs) in the primary and metastasizing tumor, which facilitate the generation of a supportive niche; and so on. [2,84,85]. Furthermore, the information provided by each assay exhibits cell-type and experimental-system associated context dependency [86]. Hence, an acknowledgement of the inherent shortcomings for each methodology is crucial, prior to the derivation of relevant conclusions, and may necessitate the development of improved protocols.
2.3. Scrutinizing Outcomes of Metastasis Assays
Most conventional in vitro assays associate metastasis with migratory and invasive capacities, while the processes of cell dissociation and colonization remain under-represented. Isolated read-outs can misrepresent the metastatic cascade and hinder effective translation of in vitro observations. Herein, we summarize the relevant biological limitations associated with in vitro read-outs (Table S2), and emphasize that a routine re-evaluation of assays is necessary for deriving appropriate clinical inferences.
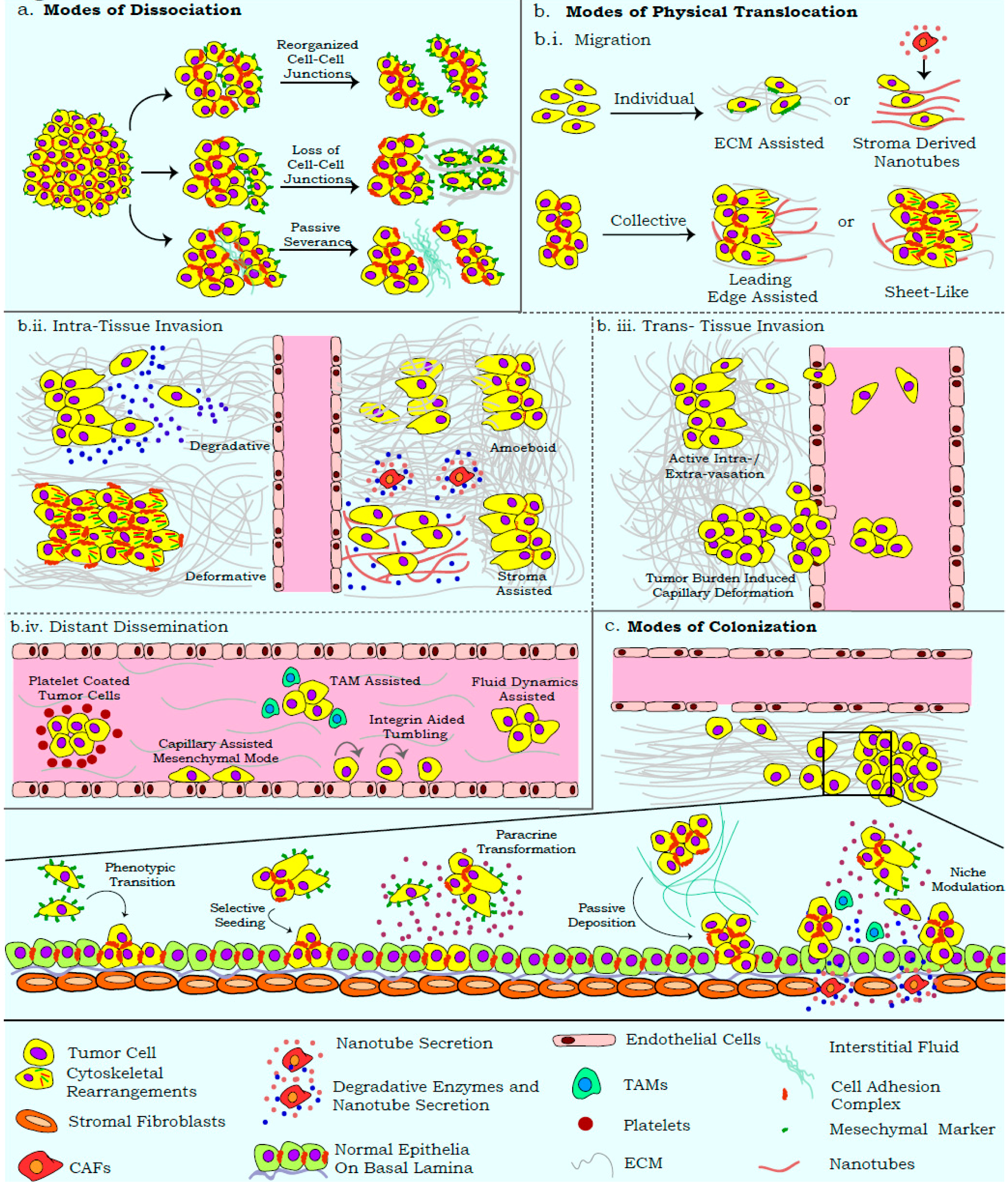
The effective dissociation of cells from the primary tumor generates varied metastasizing entities (heterogeneous cell aggregates, single mesenchymal cells, and epithelial cell clusters) based on their modes of severance [37,87,88]. Differential molecular programs activated during cell dissociation from the primary tumor endow cells with functional signatures that can influence assay read-outs (Figure 2a). The acquisition of anoikis resistance by these entities is influenced by membrane dynamics, modified secretome, immune/stromal cell recruitment, and forces generated by primary tissue/interstitial fluids; parameters that are often neglected by in vitro assays [36,89,90]. Similar influences also distinguish cell aggregates from spheroids, and demand unambiguous identification approaches for these suspension entities [37]. The re-organization of tissue architecture during metastasis further employs processes that can degrade and/or realign specific ECM components in order to ensure optimal dissemination and colonization (Figure 2b.i) [91,92,93]. While ECM-based assays discern the molecular players of cell adhesion and degradation, there is limited comprehension of the influences from 3D-ECM rearrangement. For example, with minimal biochemical changes, collagens can undergo rearrangement with respect to fiber density and crosslinking, so as to alter cell–ECM interactions [94]. Similarly, degradation assays more commonly quantify the biochemical aspect of cell–ECM interaction, while the mechanical forces that can distort ECM arrangement are largely ignored (Figure 2b.ii) [43,95,96]. Existing assays also disregard the amoeboid mode of invasion, wherein cells exhibit a greater degree of deformability, which facilitates displacement across the ECM with minimal biochemical or mechanical alterations of the matrix [97,98].
In vitro migration and invasion assays present with similar shortcomings, wherein the heterogeneity and mechanics of these processes are excessively simplified and the modalities of translocation are overlooked. Wound closure assays widely imply cell migration, while disregarding the proliferative potential of the wound edge [99]. Similarly, experimental systems often employ scratch and gap closure assays interchangeably, with a complete disregard for the differential biological annotations represented by each method. While “gaps” (cell-free zones) are passively generated by artificial barriers positioned around proliferating cells, scratch assays involve the active disruption of cell monolayers. These differences can influence the activation of differential molecular networks and affect assay outputs [40,41]. Furthermore, trans-well assays restrict the movement of collectively invading cells, because of the pre-defined insert pore size that usually permits the passage of single cells. Similarly, co-culture systems often do not account for the paracrine effects of invading tumor cells on the epithelial/endothelial monolayers; these can activate the trans-differentiation and chemotaxis programs involved in the recruitment of CAFs, TAMs, and so on [84,85]. Most read-outs for migration and invasion also assume an onset of EMT, while the contributions of extra-tumoral cells recruited for their cooperative effects are often disregarded (Figure 2b.ii) [100]. Similarly, the plasticity of migratory/invasive modalities in response to differential ECM density, composition, and arrangement also requires critical scrutiny [101].
While several assays depict the properties associated with cell dissemination, the direct detection of metastatic seeding is not achieved in vitro. Metastasis-associated studies also disregard the stages of intra-/extra-vasation and the associated cellular plasticity that may govern efficacy of the metastatic cascade [102,103]. Importantly, routine in vivo models are usually limited by their inherent immunodeficiency that impacts the heterogeneity and efficiency of metastatic dissemination [104]. Similarly, the ectopic transplantation of tumor cells in mice and the infiltration of mouse stroma in these models make them less suitable for studying the role of tumor microenvironment on metastatic dissemination [104]. Thus, despite the existence of numerous elegant model systems, the complexity of metastasis necessitates regular improvements in assay resolution, and emphasizes the inclusion of recent ideologies when inferring from these outputs.
3. Uncoupling the Migration–Invasion–Metastasis Ideology
Tumors hijack several molecular programs associated with organogenesis; hence, concepts derived from developmental systems could elucidate the functional attributes observed during tumor progression and metastasis. Apart from tumor cells, the recruitment of tissue stroma/immune components vastly influences each stage of the metastatic cascade in a context dependent manner. The exclusion of several physiological parameters from experimental systems may stem from the limited familiarity and visualization of such processes, which can limit their collation with clinical observations. In this section, we aim to highlight the alternate physiological mechanisms adopted by tumors that uncouple the traditional migration–invasion–metastasis ideology, and warrant inclusion in routine assays (Figure 2).
The dissociation of cells from the primary tumor is widely attributed to active EMT as well as its molecular manifestations that affect cell adhesion and cytoskeletal complexes [105]; other studies highlight the realignment of adhesion complexes towards the generation of passively disseminating epithelial cell clusters [19]. Passive dissemination presents a challenge for experimental detection, although its contribution to metastasis is undeniable (Figure 2a) [106,107]. The degradative secretome that often accompanies EMT can further lead to the severance of single cells from the primary mass [96]. Tumors are also subject to hydrostatic pressures from organ-specific interstitial fluids that slough off weakly connected proliferative cell masses. Separately, the tumor edge can undergo cytoskeleton mediated delamination as epithelial sheets, a feature also noted during development [108].
Several existing studies associate metastatic efficacy with EMT facilitated migration and invasion [109,110,111]. However, recent reports highlight the differential migratory and invasive capabilities of tumor cells, significantly influenced by the microenvironment (Figure 2b.i). Passively dissociated cells can disperse and seed as proximal metastatic lesions in the absence of active migration/invasion programs [106,112]. Furthermore, efficient cell migration along chemo-/duro-tactic gradients relies on the sustained cellular contacts that underscore the existence of epithelial properties and fortify collective migration as a crucial process in disseminating tumor cells [112]. Recent studies also highlight the role of tumor/stroma cell derived membrane invaginations, termed “nanotubes” that can alter ECM arrangement and serve as directed migratory tracks for cancer cells [113].
The uncoupling of EMT from the metastatic cascade is also evident from reports highlighting the invasion of epithelial cell clusters by virtue of tensile forces exerted on the ECM (Figure 2b.ii) [114,115]. Invasion is also reported to be mediated by an amoeboid transition, which allows cells to squeeze through the ECM, without its biochemical or mechanical alteration [98]. Invasion modalities can be influenced by organ-specific architecture, which presents as an anatomical barrier for metastasis. For example, fenestrated bone marrow sinusoids offer a lower mechanical challenge for invasion, as opposed to the blood–brain barrier [116]. Distal metastasis is further ascertained by the density of lymphatic/blood vessels, and the ability of tumor cells to intra- and extra-vasate; invasive cell clusters capable of disrupting the basal lamina and entering circulation may stay lodged in the blood vessels and fail to extravasate (Figure 2b.iii) [117]. Tumor cells in capillaries or lymphatic vessels ensure survival in response to immune surveillance and hydrostatic pressures by existing as cell clusters or recruiting cells from the hematopoietic lineage [2]. Eventually, cells achieve distant dissemination under the influence of systemic circulatory flow, and can also exhibit migration along the capillary linings by virtue of mesenchymal motility or integrin mediated tumbling (Figure 2b.iv) [118,119].
Several of the above phenomena are reversed during the colonization stage of metastasis, wherein tumor cells re-establish cellular contacts and lodge into a supportive niche (Figure 2c). The reversal of certain mesenchymal properties is thus deemed crucial for colonization, as reports highlight the inability of rigid mesenchymal cells to metastasize, despite in vitro migratory and invasive capabilities [34,120]. Phenotypic plasticity plays a key role in enhancing the adaptability of cells to the altered niche, and may generate a heterogeneous disease at distinct sites in response to varying microenvironments [121,122]. The passive deposition and selective seeding of epithelial tumor cells from heterogeneous metastasizing clusters can dictate the efficacy of metastasis [106,107,112]. Sculpting of the secondary site is also undertaken by tumor cells, wherein paracrine signaling can result in a transformation of the secondary tissue or activate stromal cells, and generate a microenvironment conducive to metastases seeding [100]. Insights into the role of these extra-tumoral cellular components have been reviewed elsewhere [84,85]. Thus, a definitive comprehension of the metastatic cascade necessitates an approach wherein the aforementioned processes can be integrated into informative protocols.
4. Visualization of Metastatic Modalities with Real-Time Approaches
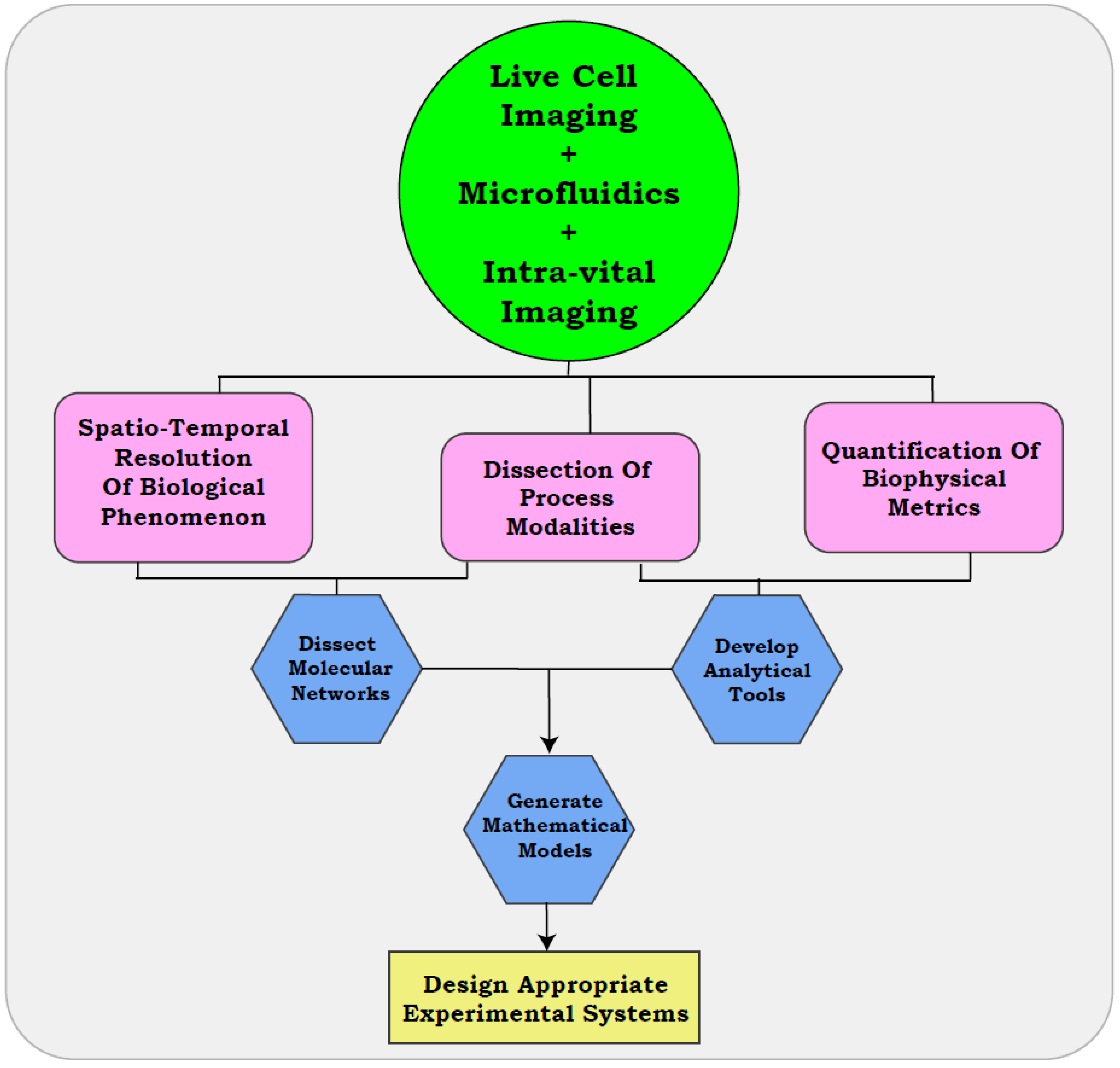
As discussed in previous sections, comprehending the biological heterogeneity of metastasis is often limited by the poor resolution of snapshot assays. Existing assays can provide an enhanced resolution of the cellular functionalities, when coupled with microfluidic approaches, real-time visualization, and intra-vital imaging. These methods can further reform traditional ideologies by including physiologically relevant variables and the development of quantitative parameters to delineate the metastatic cascade (Figure 3).
4.1. Use of Microfluidics
Microfluidic devices are recent advances in automation that have been applied in order to elucidate the distinct ECM components, widths of migratory paths, cell adhesion forces associated with collective migration, and the effect of extrinsic fluid pressures, amongst other metrics associated with metastasis [103,123,124,125]. In a recent study, an artificial circulatory system was generated on PDMS (polydimethylsiloxane) micro-capillaries lined with functional endothelial cells. This system was employed to model extra-/intra-vasation modalities under influences of extrinsic fluid pressure [102]. Label-free isolation and characterization of viable CTC clusters from blood samples has been achieved with Cluster-Chip, a microchip technology employing bifurcation traps. CTCs released from the microchip could be subjected to functional and molecular characterization in vitro, to comprehend their role in the metastatic cascade [126]. Similarly, an organ chip bioengineered to mimic the 3D microvascular networks has been effective in capturing and quantifying trans-endothelial migration, seeding, and micro-metastases formation [127]. Similar approaches have also used 3D bio-matrices to generate tissue models for studies pertaining to the interaction and infiltration of cancer cells [128,129,130]. Recently, the co-culture of breast cancer cells and CAFs on a microfluidic device successfully established a 3D-organotypic model to mimic stroma-driven tumor cell invasion. Live cell imaging coupled with transcriptome analysis of the co-cultured cells further discerned the novel molecular targets associated with invasion [131]. The recapitulation of spatio-temporal metabolic adaptations encountered by tumor cells was also achieved with a microfluidic organotypic breast cancer model, and employed to devise therapeutic strategies for targeting hypoxic cells [132].
4.2. In Vitro Resolution of Metastatic Modalities with Live Cell Imaging
Independent of microfluidic devices, live cell imaging has enabled the quantification of various functional properties by visually monitoring routine in vitro protocols. The spontaneous detachment of cells from 3D culture models of ovarian cancer was recently visualized with live cell imaging. These observations allowed for the development of in vivo lineage tracing methods, and discerned a collective dissociation of cells from the primary tumor, indicating the onset of metastasis [37]. A recent study from our group, coupling live cell imaging with the in vitro scratch assay, not only discerned the distinct migratory modalities in ovarian carcinoma (CCM versus EMT), but also resolved subtle variations within CCM; this defined CCM as being mediated either through proliferation (passive CCM) or sheet-like migration (active CCM). We quantified the altered migratory modalities in response to extrinsic and intrinsic stimuli, further establishing an improved prototype of the scratch assay [99]. In a separate study, CCM was assessed in the context of durotaxis, wherein live cell imaging successfully quantified the extent and pattern of migration in response to ECM variations [133]. Similar approaches were applied to invasion assays, so as to determine the morphological variations of patient derived spheroids invading into a carboxy–methyl cellulose matrix [134]. Separately, live cell imaging of the migrating cells in a confined micro-pillar array permitted the derivation of distinct quantitative parameters for distinguishing the individual and collective modes of migration [135].
Real-time studies can thus offer an improved comprehension of the bio-mechanical features associated with metastasis that are often overlooked by snapshot assays (Table S3). The corroboration of time-lapse microscopy with trans-well invasion assays could distinguish the degradative, deformative, and amoeboid modes of cell invasion [98,136]. Similar approaches with co-culture systems can delineate the proliferation driven disruption of endothelial linings from active events of intra-/extra-vasation, and in suspension cultures, distinguish the mechanical forces associated with spheroids versus cellular aggregates [38,115,137]. The assay resolution can be further enhanced by employing fluorescent dyes or protein tags that differentially label cell membranes, ECM fibers, cytoskeletal components, and so on. Importantly, the labelling methods adopted for real-time imaging studies must be permissive of the assay systems, and contribute minimal hindrance to the biological process under study [138,139]. Such real-time analysis can also be implemented in high-throughput screens, along with traditional assay outputs, so as to address the effects of growth additives and pharmacological compounds on diverse biological modalities.
4.3. Intra-Vital Imaging Assisted Visualization of Cellular Properties
As discussed previously, the limited in vitro recapitulation of the heterogeneity associated with metastasis has compelled cancer biologists to use animal models suitable for varied experimental approaches. The in vivo dynamics of migration and invasion have been previously noted in C. elegans, fruit fly, and zebrafish [67]. Recent advances in microscopy and fluorophore chemistry have also permitted the intra-vital single-cell resolution in vertebrate animal models. In a mouse model, intra-vital imaging assisted approaches elucidated the role of Rac1-dependent membrane protrusions in the maintenance of 3D spatial positions of mouse dermal fibroblasts during wound healing [140]. In yet another report, the transplantation of tumor cells into the mouse cerebellopontine angle region coupled with intra-vital imaging successfully developed a model to study the disease progression of vestibular schwannoma [141]. Separately, the intra-vital imaging of early breast carcinoma lesions identified a Her2 driven dissemination program that enables the dispersal of dormant cancer cells to distant organs, prior to the development of a primary tumor [142]. Intra-vital imaging also successfully discerned the significance of orthotopic, as opposed to subcutaneous, tumor models in prostate cancer wherein drastic differences in the overall vascularization and distant metastasis were duly noted [143]. Intra-vital imaging has been aided by confetti mouse models, which express a combination of fluorescent proteins in response to the Cre-recombinase, and permit the visualization of the in vivo heterogeneity inherent to tissue systems [144,145]. Confetti mice have been instrumental in comprehending squamous cell carcinoma progression, wherein the recruitment of the adjacent epithelium by monoclonal papillomas and intra-tumoural invasion by newly generated clones were effectively visualized [146]. Cell of origin studies in castration resistance prostate cancer, employing confetti mice, have previously assigned tumor formation to a population of Bmi1+ve luminal cells in the proximal prostate [83]. The in vivo fate and properties of transplanted corneal epithelium were also resolved with intra-vital imaging, which tracked the fluorescence signals from the confetti tagged donor cells [147].
Other advances in imaging technologies include improved tomography scans, fluorescence lifetime imaging microscopy (FLIM), coherent anti-Stokes Raman microscopy (CARS), and so on, and have been reviewed elsewhere in detail [148,149]. The routine application of such high-resolution approaches may be beneficial towards the development of more informative protocols, the quantification of key biomechanical features, and could overcome several limitations harbored by snapshot studies.
5. Quantitative Resolution of Biological Modalities
Examples of real-time assays discussed in previous sections not only emphasize their visual impact, but also highlight the derivation of varied quantitative metrics. The integration of live cell imaging into routine approaches can quantify diverse physiological parameters and enhance assay read-outs. We briefly familiarize readers with recent approaches developed for high-resolution real-time imaging, user-friendly analytical tools, and quantitative variables generated from imaging datasets (Table S3).
Continued technological advances and automation in the field of optics have provided high-resolution microscopes that facilitate the real-time visualization of dynamic biological processes, including the migration of metastatic cancer cells in vivo [150]. Similarly, atomic force microscopy (AFM) and protrusion force microscopy (PFM) have been employed to discern the mechanical forces involved in leukocyte–endothelial adhesion and invadosome–assisted extravasation, respectively [151]. CARS coupled with two-photon imaging has also emerged as a promising approach to describe multiple aspects of the tumor niche and disease progression, with minimal photo-damage to the specimen [73]. Interested readers may refer to several recent articles elaborating on the advances in microscope design, with significant implications in real-time cancer research [148,149].
Datasets generated by real-time imaging present a significant challenge for analysis, thus necessitating familiarity with relevant software and tools. While the adaptability of imaging tools across model systems is beneficial, optimal quantification can only be ensured when high-resolution/-contrast images are provided as inputs. The availability of user-friendly interfaces, improvements with recent plugins, and the development of codes for the customization of outputs enhance the applicability of imaging tools. Currently, a variety of programs facilitate time-lapse image analysis, and are available either commercially, viz., Metamorph (Olympus), Imaris (Bitplane), Zen (Zeiss), Elements (Nikon), Volocity (PerkinElmer), and so on, or as open source platforms, viz., Micro-manager, ImageJ, MotilityLab, and so on. Imaris is specifically useful for 4D data analysis, and can be substituted with the ImageJ post-installation of plugins that facilitate drift correction, volume measurement, free rotation, and so on [152]. COMBImage, a recent computational framework, has facilitated the automated analyses of cell morphology and confluence in cell viability assays [153]. The management and analysis of multi-dimensional imaging data have also been aided by support vector machines (SVM), which have been applied to group pixels for image segmentation, to detect cellular/sub-cellular phenotypes, or to categorize developmental stages at the level of entire organisms [154]. Similarly, mathematical approaches for migration track analysis have been developed [155]. A more comprehensive list of the currently available image processing software and tools is available elsewhere [156,157].
Analytical tools can yield extensive information from imaging datasets, and often confound untrained individuals. The extraction of relevant quantitative metrics must consider the biophysical properties of a system and examine the associative trends with the cellular processes under study. Examples highlighting the selection of quantitative metrics are summarized in Table S3, and include the micro-pillar array-based study, wherein six quantitative parameters extracted from individual cells generated a binary solidification model for describing the interconversion between collective versus individual modes of migration [135], fractal analysis that described the developing dermis as a lattice structure resulting from the dynamic stroma [158], derivation of migration modalities in ovarian cancer cell lines assisted by coupling live cell imaging with the in vitro scratch assay [99], and so on. Additional studies highlight the vast array of quantitative measures derived from real-time imaging datasets, and signify their contributions to an improved understanding of the biological processes [11,151,159,160,161,162].
6. The Era of Live Cell Imaging
The simple corroboration of time-lapse microscopy with endpoint assays can reveal the modalities of several biological processes, indicate differential routes of metastasis taken by tumors, and identify the associated molecular pathways (Figure 3). Collation of these datasets can generate multi-parametric simulations that define the biological thresholds associated with diverse cellular functions, and their modulation in response to various cues [163,164,165]. Such models or simulations could aid in the design of appropriate experimental systems that encompass heterogeneous processes contributing to metastasis. Similarly, analytical tools developed with real-time approaches can be integrated into drug development pipelines [166]. In accordance with our statements, a recent study employing intra-vital imaging discerned the crucial role of CD44 driven cell aggregation of CTCs in metastatic dissemination. The study established CD44–PAK2 interaction as a driving event of the process, thus providing clinically relevant information with the aid of visual outputs [167]. While snap-shot assays will continue to provide high-throughput readouts for cell systems, it is crucial that experimentalists acknowledge their limitations when deriving clinically relevant inferences. With the right amount of caution, real-time approaches could herald an “era of live cell imaging” in cancer biology.
Real-time approaches, in conjunction with lineage tracing, may initiate the next phase of resolving cancer heterogeneity, which continues to elude comprehension and is responsible for therapy resistance. In view of recent reports, real-time approaches may decipher functional nuances and provide insight into the concept of “metastability” predicted during phenotypic switches. Real-time dynamics of molecular turn-over can further identify the spatiotemporal alterations of markers over the course of disease progression, and improve clinical interventions. Moreover, intra-vital imaging approaches present an exciting opportunity to tackle a residual minimal disease, which often results in an aggressive relapse and overall poor prognosis. Similarly, extra-tumoral cell populations crucial for disease progression in vivo can be identified and probed in vitro with micro-fluidic and real-time studies. More importantly, the stages of primary dissociation and colonization during metastasis represent a “grey area” of information, which could be effectively resolved with the previously discussed approaches. The delineation of these diverse biophysical features essentially nurtures the model of personalized medicine. A careful combination of appropriate existing assays with live cell imaging can reveal several nuances of the metastatic cascade and reiterate the “seeing is believing” ideology.
7. Conclusions
Recent studies reveal a functional uncoupling of several physiological processes previously deemed crucial for successful metastasis. Our own observations associated with the evaluation of in vitro metastasis reflect on the shortcomings of routine assays. For the detection of distinct modalities associated across the metastatic cascade, we suggest the adoption of time lapse imaging in routine assays. This approach has the capability to (i) make a visual impact on biological processes, (ii) identify different routes undertaken by tumor cells towards achieving these functions, and (iii) provide metrics for quantifying these processes. The information obtained from these approaches can be applied towards establishing mathematical models and intuitive inferences that tackle the heterogeneity associated with disease progression. Inherent cellular plasticity that contributes to tissue homeostasis can also be deciphered in the context of pathological conditions, by developing novel assays that combine the ease of traditional approaches with the enhanced resolution of real-time imaging. We also emphasize on the routine re-evaluation of functional read-outs, so as to incorporate recent conceptual developments that improve the comprehension of biological systems. Together, these features are likely to be useful in elucidating the poorly understood areas of disease progression in cancer, and deriving clinically relevant conclusions over the current assays.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2077-0383/8/7/941/s1: Table S1. Metastasis associated functional read-outs. Details pertaining to the setup, quantification, and biological read-out associated with the in vitro and in vivo methods employed to study the metastatic cascade are enlisted. Table S2. Applications and limitations of metastasis associated functional read-outs. Details pertaining to the cellular functionality gauged, stage of metastasis represented, and inherent limitations of the in vitro and in vivo methods employed to study the metastatic cascade are enlisted. Table S3. The real-time approach. Details pertaining to the model systems, relevant instrumentation, quantitative tools and variables, and their biological relevance for recent real-time studies are enlisted. The table includes a summary of a limited number of studies, interested readers may refer to the available literature cited in the text for more information.
Author Contributions
Conceptualization, S.A.B. and S.S.V.; investigation, S.S.V.; writing (original draft preparation), S.A.B. and S.S.V.; funding acquisition, S.A.B.
Funding
This work was supported by funding to SAB from NCCS, Pune (Intramural), and the Department of Biotechnology, Government of India, New Delhi (Extramural grant, BT/Indo-Aus/06/03/2011). The research fellowship was availed by SSV from Council of Scientific and Industrial Research, New Delhi, India.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chaffer, C.L.; San Juan, B.P.; Lim, E.; Weinberg, R.A. EMT, cell plasticity and metastasis. Cancer Metastasis Rev. 2016, 35, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Nieto, M.A.; Huang, R.Y.Y.J.; Jackson, R.A.A.; Thiery, J.P.P. Emt: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, T. To differentiate or not-routes towards metastasis. Nat. Rev. Cancer 2012, 12, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.; Casanova, J. A common framework for EMT and collective cell migration. Development 2016, 143, 4291–4300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastushenko, I.; Brisebarre, A.; Sifrim, A.; Fioramonti, M.; Revenco, T.; Boumahdi, S.; Van Keymeulen, A.; Brown, D.; Moers, V.; Lemaire, S.; et al. Identification of the tumour transition states occurring during EMT. Nature 2018, 556, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Jolly, M.K.; Mani, S.A.; Levine, H. Hybrid epithelial/mesenchymal phenotype(s): The ‘fittest’ for metastasis? Biochim. Biophys. Acta Rev. Cancer 2018, 1870, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.H.; Yang, J. Epithelial—Mesenchymal plasticity in carcinoma metastasis. Genes Dev. 2013, 27, 2192–2206. [Google Scholar] [CrossRef]
- Chung, W.; Eum, H.H.; Lee, H.O.; Lee, K.M.; Lee, H.B.; Kim, K.T.; Ryu, H.S.; Kim, S.; Lee, J.E.; Park, Y.H.; et al. Single-cell RNA-seq enables comprehensive tumour and immune cell profiling in primary breast cancer. Nat. Commun. 2017, 8, 15081. [Google Scholar] [CrossRef] [Green Version]
- MacLean, A.L.; Hong, T.; Nie, Q. Exploring intermediate cell states through the lens of single cells. Curr. Opin. Syst. Biol. 2018, 9, 32–41. [Google Scholar] [CrossRef]
- Bednarz-Knoll, N.; Alix-Panabières, C.; Pantel, K. Plasticity of disseminating cancer cells in patients with epithelial malignancies. Cancer Metastasis Rev. 2012, 31, 673–687. [Google Scholar] [CrossRef]
- Labernadie, A.; Kato, T.; Brugués, A.; Serra-Picamal, X.; Derzsi, S.; Arwert, E.; Weston, A.; González-Tarragó, V.; Elosegui-Artola, A.; Albertazzi, L.; et al. A mechanically active heterotypic E-cadherin/N-cadherin adhesion enables fibroblasts to drive cancer cell invasion. Nat. Cell Biol. 2017, 19, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Aman, A.; Piotrowski, T. Wnt/β-Catenin and Fgf Signaling Control Collective Cell Migration by Restricting Chemokine Receptor Expression. Dev. Cell 2008, 15, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Barham, W.; Chen, L.; Tikhomirov, O.; Onishko, H.; Gleaves, L.; Stricker, T.P.; Blackwell, T.S.; Yull, F.E. Aberrant activation of NF-κB signaling in mammary epithelium leads to abnormal growth and ductal carcinoma in situ. BMC Cancer 2015, 15, 647. [Google Scholar] [CrossRef] [PubMed]
- Köthe, S.; Müller, J.P.; Böhmer, S.-A.; Tschongov, T.; Fricke, M.; Koch, S.; Thiede, C.; Requardt, R.P.; Rubio, I.; Böhmer, F.D. Features of Ras activation by a mislocalized oncogenic tyrosine kinase: FLT3 ITD signals through K-Ras at the plasma membrane of acute myeloid leukemia cells. J. Cell Sci. 2013, 126, 4746–4755. [Google Scholar] [CrossRef] [PubMed]
- Celestini, V.; Tezil, T.; Russo, L.; Fasano, C.; Sanese, P.; Forte, G.; Peserico, A.; Lepore Signorile, M.; Longo, G.; De Rasmo, D.; et al. Uncoupling FoxO3A mitochondrial and nuclear functions in cancer cells undergoing metabolic stress and chemotherapy. Cell Death Dis. 2018, 9, 231. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.J.; Baker, J.G.; Rees, S. Reporter-gene systems for the study of G-protein-coupled receptors. Curr. Opin. Pharmacol. 2001, 1, 526–532. [Google Scholar] [CrossRef]
- Fan, F.; Wood, K.V. Bioluminescent Assays for High-Throughput Screening. Assay Drug Dev. Technol. 2007, 5, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Hirata, E.; Park, D.; Sahai, E. Retrograde flow of cadherins in collective cell migration. Nat. Cell Biol. 2014, 16, 621–623. [Google Scholar] [CrossRef]
- Peglion, F.; Llense, F.; Etienne-Manneville, S. Adherens junction treadmilling during collective migration. Nat. Cell Biol. 2014, 16, 639–651. [Google Scholar] [CrossRef]
- Zhang, L.; Luo, J.; Wan, P.; Wu, J.; Laski, F.; Chen, J. Regulation of cofilin phosphorylation and asymmetry in collective cell migration during morphogenesis. Development 2011, 138, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Henkel, L.; Rauscher, B.; Boutros, M. Context-dependent genetic interactions in cancer. Curr. Opin. Genet. Dev. 2019, 54, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Kosztyu, P.; Slaninová, I.; Valčíková, B.; Verlande, A.; Müller, P.; Paleček, J.J.; Uldrijan, S. A Single Conserved Amino Acid Residue as a Critical Context-Specific Determinant of the Differential Ability of Mdm2 and MdmX RING Domains to Dimerize. Front. Physiol. 2019, 10, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richelle, A.; Chiang, A.W.T.; Kuo, C.C.; Lewis, N.E. Increasing consensus of context-specific metabolic models by integrating data-inferred cell functions. PLoS Comput. Biol. 2019, 15, e1006867. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Wang, X.; Li, X.; Meng, L.; Xu, F.; Xu, Y.; Xie, X.; He, H.; Xu, D.; Wang, C.; et al. Apogossypolone acts as a metastasis inhibitor via up-regulation of E-cadherin dependent on the GSK-3/AKT complex. Am. J. Transl. Res. 2019, 11, 218–232. [Google Scholar] [PubMed]
- Menezes, S.V.; Fouani, L.; Huang, M.L.; Geleta, B.; Maleki, S.; Richardson, A.; Richardson, D.R.; Kovacevic, Z. The metastasis suppressor, NDRG1, attenuates oncogenic TGF-β and NF-κB signaling to enhance membrane E-cadherin expression in pancreatic cancer cells. Carcinogenesis 2018. [Google Scholar] [CrossRef]
- Bendardaf, R.; Sharif-Askari, F.S.; Sharif-Askari, N.S.; Syrjänen, K.; Pyrhönen, S. Cytoplasmic E-Cadherin Expression Is Associated With Higher Tumour Level of VEGFA, Lower Response Rate to Irinotecan-based Treatment and Poorer Prognosis in Patients With Metastatic Colorectal Cancer. Anticancer Res. 2019, 39, 1953–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiello, N.M.; Maddipati, R.; Norgard, R.J.; Balli, D.; Li, J.; Yuan, S.; Yamazoe, T.; Black, T.; Sahmoud, A.; Furth, E.E.; et al. EMT Subtype Influences Epithelial Plasticity and Mode of Cell Migration. Dev. Cell 2018, 45, 681–695.e4. [Google Scholar] [CrossRef] [Green Version]
- Reichert, M.; Bakir, B.; Moreira, L.; Pitarresi, J.R.; Feldmann, K.; Simon, L.; Suzuki, K.; Maddipati, R.; Rhim, A.D.; Schlitter, A.M.; et al. Regulation of Epithelial Plasticity Determines Metastatic Organotropism in Pancreatic Cancer. Dev. Cell 2018, 45, 696–711. [Google Scholar] [CrossRef]
- Khalil, A.A.; Ilina, O.; Gritsenko, P.G.; Bult, P.; Span, P.N.; Friedl, P. Collective invasion in ductal and lobular breast cancer associates with distant metastasis. Clin. Exp. Metastasis 2017, 34, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Denecker, G.; Vandamme, N.; Akay, Ö.; Koludrovic, D.; Taminau, J.; Lemeire, K.; Gheldof, A.; De Craene, B.; Van Gele, M.; Brochez, L.; et al. Identification of a ZEB2-MITF-ZEB1 transcriptional network that controls melanogenesis and melanoma progression. Cell Death Differ. 2014, 21, 1250–1261. [Google Scholar] [CrossRef] [Green Version]
- Krebs, A.M.; Mitschke, J.; Losada, M.L.; Schmalhofer, O.; Boerries, M.; Busch, H.; Boettcher, M.; Mougiakakos, Di.; Reichardt, W.; Bronsert, P.; et al. The EMT-activator Zeb1 is a key factor for cell plasticity and promotes metastasis in pancreatic cancer. Nat. Cell Biol. 2017, 19, 518–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caramel, J.; Papadogeorgakis, E.; Hill, L.; Browne, G.J.; Richard, G.; Wierinckx, A.; Saldanha, G.; Osborne, J.; Hutchinson, P.; Tse, G.; et al. A Switch in the Expression of Embryonic EMT-Inducers Drives the Development of Malignant Melanoma. Cancer Cell 2013, 24, 466–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, S.; Reichert, M.; Bakir, B.; Das, K.K.; Nishida, T.; Miyazaki, M.; Heeg, S.; Collins, M.A.; Marchand, B.; Hicks, P.D.; et al. Prrx1 isoform switching regulates pancreatic cancer invasion and metastatic colonization. Genes Dev. 2016, 30, 233–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocaña, O.H.; Córcoles, R.; Fabra, Á.; Moreno-Bueno, G.; Acloque, H.; Vega, S.; Barrallo-Gimeno, A.; Cano, A.; Nieto, M.A. Metastatic Colonization Requires the Repression of the Epithelial-Mesenchymal Transition Inducer Prrx1. Cancer Cell 2012, 22, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Keckesova, Z.; Donaher, J.L.; Shibue, T.; Tischler, V.; Reinhardt, F.; Itzkovitz, S.; Noske, A.; Zürrer-Härdi, U.; Bell, G.; et al. Slug and Sox9 cooperatively determine the mammary stem cell state. Cell 2012, 148, 1015–1028. [Google Scholar] [CrossRef] [PubMed]
- Taddei, M.L.; Giannoni, E.; Fiaschi, T.; Chiarugi, P. Anoikis: An emerging hallmark in health and diseases. J. Pathol. 2012, 226, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Al Habyan, S.; Kalos, C.; Szymborski, J.; McCaffrey, L. Multicellular detachment generates metastatic spheroids during intra-abdominal dissemination in epithelial ovarian cancer. Oncogene 2018, 37, 5127–5135. [Google Scholar] [CrossRef] [PubMed]
- Kramer, N.; Walzl, A.; Unger, C.; Rosner, M.; Krupitza, G.; Hengstschläger, M.; Dolznig, H. In vitro cell migration and invasion assays. Mutat. Res. Rev. Mutat. Res. 2013, 752, 10–24. [Google Scholar] [CrossRef]
- Choi, M.; Yu, S.J.; Choi, Y.; Lee, H.R.; Lee, E.; Lee, E.; Lee, Y.; Song, J.; Son, J.G.; Lee, T.G.; et al. Polymer Thin Film-Induced Tumor Spheroids Acquire Cancer Stem Cell-like Properties. Cancer Res. 2018, 78, 6890–6902. [Google Scholar] [CrossRef]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef]
- Das, A.M.; Eggermont, A.M.M.; Ten Hagen, T.L.M. A ring barrier-based migration assay to assess cell migration in vitro. Nat. Protoc. 2015, 10, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Dowling, C.M.; Herranz Ors, C.; Kiely, P.A. Using real-time impedance-based assays to monitor the effects of fibroblast-derived media on the adhesion, proliferation, migration and invasion of colon cancer cells. Biosci. Rep. 2014, 34, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Anderl, J.; Ma, J.; Armstrong, L. Fluorescent Gelatin Degradation Assays for Investigating Invadopodia Formation. Nat Methods 2012, 121007, 1–6. [Google Scholar]
- Dong, Y.; Stephens, C.; Walpole, C.; Swedberg, J.E.; Boyle, G.M.; Parsons, P.G.; McGuckin, M.A.; Harris, J.M.; Clements, J.A. Paclitaxel Resistance and Multicellular Spheroid Formation Are Induced by Kallikrein-Related Peptidase 4 in Serous Ovarian Cancer Cells in an Ascites Mimicking Microenvironment. PLoS ONE 2013, 8, e57056. [Google Scholar] [CrossRef] [PubMed]
- Nakai, K.; Tanaka, T.; Murai, T.; Ohguro, N.; Tano, Y.; Miyasaka, M. Invasive human pancreatic carcinoma cells adhere to endothelial tri-cellular corners and increase endothelial permeability. Cancer Sci. 2005, 96, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Narai, A.; Arai, S.; Shimizu, M. Rapid decrease in transepithelial electrical resistance of human intestinal Caco-2 cell monolayers by cytotoxic membrane perturbents. Toxicol. Vitr. 1997, 11, 347–354. [Google Scholar] [CrossRef]
- Francart, M.E.; Lambert, J.; Vanwynsberghe, A.M.; Thompson, E.W.; Bourcy, M.; Polette, M.; Gilles, C. Epithelial–mesenchymal plasticity and circulating tumor cells: Travel companions to metastases. Dev. Dyn. 2018, 247, 432–450. [Google Scholar] [CrossRef]
- Bardelli, A.; Pantel, K. Liquid Biopsies, What We Do Not Know (Yet). Cancer Cell 2017, 31, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Alpaugh, K.R.; Cristofanilli, M. Circulating tumor cells (CTCs) in breast cancer: A diagnostic tool for prognosis and molecular analysis. Chin. J. Cancer Res. 2012, 24, 388–398. [Google Scholar] [CrossRef]
- Jolly, M.K. Implications of the Hybrid Epithelial/Mesenchymal Phenotype in Metastasis. Front. Oncol. 2015, 5, 155. [Google Scholar] [CrossRef] [Green Version]
- Nel, I.; Gauler, T.C.; Bublitz, K.; Lazaridis, L.; Goergens, A.; Giebel, B.; Schuler, M.; Hoffmann, A.C. Circulating tumor cell composition in renal cell carcinoma. PLoS ONE 2016, 11, e0153018. [Google Scholar] [CrossRef] [PubMed]
- McInnes, L.M.; Jacobson, N.; Redfern, A.; Dowling, A.; Thompson, E.W.; Saunders, C.M. Clinical Implications of Circulating Tumor Cells of Breast Cancer Patients: Role of Epithelial–Mesenchymal Plasticity. Front. Oncol. 2015, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Alconada, L.; Muinelo-Romay, L.; Madissoo, K.; Diaz-Lopez, A.; Krakstad, C.; Trovik, J.; Wik, E.; Hapangama, D.; Coenegrachts, L.; Cano, A.; et al. Molecular profiling of circulating tumor cells links plasticity to the metastatic process in endometrial cancer. Mol. Cancer 2014, 13, 223. [Google Scholar] [CrossRef] [Green Version]
- Giussani, M.; Merlino, G.; Cappelletti, V.; Tagliabue, E.; Daidone, M.G. Tumor-extracellular matrix interactions: Identification of tools associated with breast cancer progression. Semin. Cancer Biol. 2015, 35, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, A.; Costa-Silva, B.; Shen, T.L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-metastatic niches: Organ-specific homes for metastases. Nat. Rev. Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Khalili, A.A.; Ahmad, M.R. A Review of cell adhesion studies for biomedical and biological applications. Int. J. Mol. Sci. 2015, 16, 18149–18184. [Google Scholar] [CrossRef] [PubMed]
- Borowicz, S.; Van Scoyk, M.; Avasarala, S.; Karuppusamy Rathinam, M.K.; Tauler, J.; Bikkavilli, R.K.; Winn, R.A. The Soft Agar Colony Formation Assay. J. Vis. Exp. 2014, 92, e51998. [Google Scholar] [CrossRef] [PubMed]
- Weeber, F.; Ooft, S.N.; Dijkstra, K.K.; Voest, E.E. Tumor Organoids as a Pre-clinical Cancer Model for Drug Discovery. Cell Chem. Biol. 2017, 24, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Weeber, F.; van de Wetering, M.; Hoogstraat, M.; Dijkstra, K.K.; Krijgsman, O.; Kuilman, T.; Gadellaa-van Hooijdonk, C.G.M.; van der Velden, D.L.; Peeper, D.S.; Cuppen, E.P.J.G.; et al. Preserved genetic diversity in organoids cultured from biopsies of human colorectal cancer metastases. Proc. Natl. Acad. Sci. USA 2015, 112, 13308–13311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patman, G. Pancreatic cancer: From normal to metastases-a whole gamut of pancreatic organoids. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 61. [Google Scholar] [CrossRef] [PubMed]
- Buske, P.; Przybilla, J.; Loeffler, M.; Sachs, N.; Sato, T.; Clevers, H.; Galle, J. On the biomechanics of stem cell niche formation in the gut—Modelling growing organoids. FEBS J. 2012, 279, 3475–3487. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M.; Shimokawa, M.; Date, S.; Takano, A.; Matano, M.; Nanki, K.; Ohta, Y.; Toshimitsu, K.; Nakazato, Y.; Kawasaki, K.; et al. A Colorectal Tumor Organoid Library Demonstrates Progressive Loss of Niche Factor Requirements during Tumorigenesis. Cell Stem Cell 2016, 18, 827–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Praharaj, P.P.; Bhutia, S.K.; Nagrath, S.; Bitting, R.L.; Deep, G. Circulating tumor cell-derived organoids: Current challenges and promises in medical research and precision medicine. Biochim. Biophys. Acta Rev. Cancer 2018, 1869, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Drost, J.; Clevers, H. Organoids in cancer research. Nat. Rev. Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef]
- Schumacher, L.J.; Kulesa, P.M.; McLennan, R.; Baker, R.E.; Maini, P.K. Multidisciplinary approaches to understanding collective cell migration in developmental biology. Open Biol. 2016, 6, 160056. [Google Scholar] [CrossRef]
- Stuelten, C.H.; Parent, C.A.; Montell, D.J. Cell motility in cancer invasion and metastasis: Insights from simple model organisms. Nat. Rev. Cancer 2018, 18, 296–312. [Google Scholar] [CrossRef]
- Lokman, N.A.; Elder, A.S.F.; Ricciardelli, C.; Oehler, M.K. Chick chorioallantoic membrane (CAM) assay as an in vivo model to study the effect of newly identified molecules on ovarian cancer invasion and metastasis. Int. J. Mol. Sci. 2012, 13, 9959–9970. [Google Scholar] [CrossRef]
- Stoletov, K.; Willetts, L.; Paproski, R.J.; Bond, D.J.; Raha, S.; Jovel, J.; Adam, B.; Robertson, A.E.; Wong, F.; Woolner, E.; et al. Quantitative in vivo whole genome motility screen reveals novel therapeutic targets to block cancer metastasis. Nat. Commun. 2018, 9, 2343. [Google Scholar] [CrossRef]
- Lengyel, E.; Burdette, J.E.; Kenny, H.A.; Matei, D.; Pilrose, J.; Haluska, P.; Nephew, K.P.; Hales, D.B.; Stack, M.S. Epithelial ovarian cancer experimental models. Oncogene 2014, 33, 3619–3633. [Google Scholar] [CrossRef]
- Hou, W.; Ji, Z. Generation of autochthonous mouse models of clear cell renal cell carcinoma: Mouse models of renal cell carcinoma. Exp. Mol. Med. 2018, 50, 30. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Cuadrado, L.; Tracey, N.; Ma, R.; Qian, B.; Brunton, V.G. Mouse models of metastasis: Progress and prospects. Dis. Model. Mech. 2017, 10, 1061–1074. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Downes, A.; Chau, Y.; Serrels, B.; Hastie, N.; Elfick, A.; Brunton, V.; Frame, M.; Serrels, A. In vivo imaging of the tumor and its associated microenvironment using combined CARS/2-photon microscopy. IntraVital 2015, 4, e1055430. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.; Chagani, S.; Kwong, L.N. The path to metastatic mouse models of colorectal cancer. Oncogene 2018, 37, 2481–2489. [Google Scholar] [CrossRef] [PubMed]
- Beck, B.; Lapouge, G.; Rorive, S.; Drogat, B.; Desaedelaere, K.; Delafaille, S.; Dubois, C.; Salmon, I.; Willekens, K.; Marine, J.C.; et al. Different levels of Twist1 regulate skin tumor initiation, stemness, and progression. Cell Stem Cell 2015, 16, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Sauer, B. Inducible gene targeting in mice using the Cre/lox system. Methods 1998, 14, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; Huch, M.; Kujala, P.; van de Wetering, M.; Snippert, H.J.; van Es, J.H.; Sato, T.; Stange, D.E.; Begthel, H.; van den Born, M.; et al. Lgr5+veStem Cells Drive Self-Renewal in the Stomach and Build Long-Lived Gastric Units In Vitro. Cell Stem Cell 2010, 6, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Proia, T.A.; Keller, P.J.; Gupta, P.B.; Klebba, I.; Jones, A.D.; Sedic, M.; Gilmore, H.; Tung, N.; Naber, S.P.; Schnitt, S.; et al. Genetic predisposition directs breast cancer phenotype by dictating progenitor cell fate. Cell Stem Cell 2011, 8, 149–163. [Google Scholar] [CrossRef]
- Sikandar, S.S.; Kuo, A.H.; Kalisky, T.; Cai, S.; Zabala, M.; Hsieh, R.W.; Lobo, N.A.; Scheeren, F.A.; Sim, S.; Qian, D.; et al. Role of epithelial to mesenchymal transition associated genes in mammary gland regeneration and breast tumorigenesis. Nat. Commun. 2017, 8, 1669. [Google Scholar] [CrossRef]
- Hsu, Y.-C. The Theory and Practice of Lineage Tracing. Stem Cells 2015, 33, 3197–3204. [Google Scholar] [CrossRef]
- Tekeli, I.; Aujard, I.; Trepat, X.; Jullien, L.; Raya, A.; Zalvidea, D. Long-term in vivo single-cell lineage tracing of deep structures using three-photon activation. Light Sci. Appl. 2016, 5, e16084. [Google Scholar] [CrossRef] [PubMed]
- Marx, V. Stem cells: Lineage tracing lets single cells talk about their past. Nat. Methods 2018, 15, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Yoo, Y.A.; Roh, M.; Naseem, A.F.; Lysy, B.; Desouki, M.M.; Unno, K.; Abdulkadir, S.A. Bmi1 marks distinct castration-resistant luminal progenitor cells competent for prostate regeneration and tumour initiation. Nat. Commun. 2016, 7, 12943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, M.; Barker, T.H.; Gibbons, D.L.; Kurie, J.M. The fibrotic tumor stroma. J. Clin. Investig. 2018, 128, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.B.; Yeh, E.S.; Soloff, A.C. Tumor-associated macrophages: Unwitting accomplices in breast cancer malignancy. NPJ Breast Cancer 2016, 2, 15025. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, T.; Kalluri, R.; Nieto, M.A.; Weinberg, R.A. EMT in cancer. Nat. Rev. Cancer 2018, 18, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.G.; Sanders, A.J.; Katoh, M.; Ungefroren, H.; Gieseler, F.; Prince, M.; Thompson, S.K.; Zollo, M.; Spano, D.; Dhawan, P.; et al. Tissue invasion and metastasis: Molecular, biological and clinical perspectives. Semin. Cancer Biol. 2015, 35, S244–S275. [Google Scholar] [CrossRef] [PubMed]
- Abu, M.; Muhamad, M.; Hassan, H.; Zakaria, Z.; Ali, S.A.M. Proximity coupled antenna with star geometry pattern amc ground plane. ARPN J. Eng. Appl. Sci. 2016, 11, 8822–8828. [Google Scholar]
- Zhong, X.; Rescorla, F.J. Cell surface adhesion molecules and adhesion-initiated signaling: Understanding of anoikis resistance mechanisms and therapeutic opportunities. Cell. Signal. 2012, 24, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Livas, T.; Kyprianou, N. Anoikis and EMT: Lethal “Liaisons” during Cancer Progression. Crit. Rev. Oncog. 2016, 21, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Nasrollahi, S.; Walter, C.; Loza, A.J.; Schimizzi, G.V.; Longmore, G.D.; Pathak, A. Past matrix stiffness primes epithelial cells and regulates their future collective migration through a mechanical memory. Biomaterials 2017, 146, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Ng, C.; Jana, A.; Padhi, A.; Szymanski, P.; Lee, J.S.H.; Behkam, B.; Nain, A.S. Aligned fibers direct collective cell migration to engineer closing and nonclosing wound gaps. Mol. Biol. Cell 2017, 28, 2579–2588. [Google Scholar] [CrossRef]
- Symowicz, J.; Adley, B.P.; Gleason, K.J.; Johnson, J.J.; Ghosh, S.; Fishman, D.A.; Hudson, L.G.; Stack, M.S. Engagement of collagen-binding integrins promotes matrix metalloproteinase-9-dependent E-cadherin ectodomain shedding in ovarian carcinoma cells. Cancer Res. 2007, 67, 2030–2039. [Google Scholar] [CrossRef] [PubMed]
- Drifka, C.R.; Loeffler, A.G.; Mathewson, K.; Keikhosravi, A.; Eickhoff, J.C.; Liu, Y.; Weber, S.M.; Kao, W.J.; Eliceiri, K.W. Highly aligned stromal collagen is a negative prognostic factor following pancreatic ductal adenocarcinoma resection. Oncotarget 2016, 7, 76197–76213. [Google Scholar] [CrossRef] [PubMed]
- Han, M.K.L.; de Rooij, J. Converging and Unique Mechanisms of Mechanotransduction at Adhesion Sites. Trends Cell Biol. 2016, 26, 612–623. [Google Scholar] [CrossRef]
- Alaseem, A.; Alhazzani, K.; Dondapati, P.; Alobid, S.; Bishayee, A.; Rathinavelu, A. Matrix Metalloproteinases: A challenging paradigm of cancer management. Semin. Cancer Biol. 2017, 56, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Cantelli, G.; Orgaz, J.L.; Rodriguez-Hernandez, I.; Karagiannis, P.; Maiques, O.; Matias-Guiu, X.; Nestle, F.O.; Marti, R.M.; Karagiannis, S.N.; Sanz-Moreno, V. TGF-β-Induced Transcription Sustains Amoeboid Melanoma Migration and Dissemination. Curr. Biol. 2015, 25, 2899–2914. [Google Scholar] [CrossRef]
- O’Neill, P.R.; Castillo-Badillo, J.A.; Meshik, X.; Kalyanaraman, V.; Melgarejo, K.; Gautam, N. Membrane Flow Drives an Adhesion-Independent Amoeboid Cell Migration Mode. Dev. Cell 2018, 46, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Varankar, S.S.; Bapat, S.A. Migratory Metrics of Wound Healing: A Quantification Approach for in vitro Scratch Assays. Front. Oncol. 2018, 8, 633. [Google Scholar] [CrossRef]
- Celià-Terrassa, T.; Kang, Y. Metastatic niche functions and therapeutic opportunities. Nat. Cell Biol. 2018, 20, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Reid, S.E.; Kay, E.J.; Neilson, L.J.; Henze, A.; Serneels, J.; McGhee, E.J.; Dhayade, S.; Nixon, C.; Mackey, J.B.; Santi, A.; et al. Tumor matrix stiffness promotes metastatic cancer cell interaction with the endothelium. EMBO J. 2017, 36, 2373–2389. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.D.; Searson, P.C. Live-cell imaging of invasion and intravasation in an artificial microvessel platform. Cancer Res. 2014, 74, 4937–4945. [Google Scholar] [CrossRef] [PubMed]
- Katt, M.E.; Placone, A.L.; Wong, A.D.; Xu, Z.S.; Searson, P.C. In Vitro Tumor Models: Advantages, Disadvantages, Variables, and Selecting the Right Platform. Front. Bioeng. Biotechnol. 2016, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Kersten, K.; de Visser, K.E.; van Miltenburg, M.H.; Jonkers, J. Genetically engineered mouse models in oncology research and cancer medicine. EMBO Mol. Med. 2017, 9, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Savagner, P. The epithelial-mesenchymal transition (EMT) phenomenon. Ann. Oncol. 2010, 21, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Broggini, T.; Piffko, A.; Hoffmann, C.J.; Harms, C.; Vajkoczy, P.; Czabanka, M. Passive entrapment of tumor cells determines metastatic dissemination to spinal bone and other osseous tissues. PLoS ONE 2016, 11, e0162540. [Google Scholar] [CrossRef] [PubMed]
- Yeung, T.-L.; Leung, C.S.; Yip, K.-P.; Au Yeung, C.L.; Wong, S.T.C.; Mok, S.C. Cellular and molecular processes in ovarian cancer metastasis. A Review in the Theme: Cell and Molecular Processes in Cancer Metastasis. Am. J. Physiol. Cell Physiol. 2015, 309, C444–C456. [Google Scholar] [CrossRef] [Green Version]
- Blaue, C.; Kashef, J.; Franz, C.M. Cadherin-11 promotes neural crest cell spreading by reducing intracellular tension—Mapping adhesion and mechanics in neural crest explants by atomic force microscopy. Semin. Cell Dev. Biol. 2018, 73, 95–106. [Google Scholar] [CrossRef]
- Alonso-Alconada, L.; Eritja, N.; Muinelo-Romay, L.; Barbazan, J.; Lopez-Lopez, R.; Matias-Guiu, X.; Gil-Moreno, A.; Dolcet, X.; Abal, M. ETV5 transcription program links BDNF and promotion of EMT at invasive front of endometrial carcinomas. Carcinogenesis 2014, 35, 2679–2686. [Google Scholar] [CrossRef]
- Rhim, A.D.; Mirek, E.T.; Aiello, N.M.; Maitra, A.; Bailey, J.M.; McAllister, F.; Reichert, M.; Beatty, G.L.; Rustgi, A.K.; Vonderheide, R.H.; et al. EMT and dissemination precede pancreatic tumor formation. Cell 2012, 148, 349–361. [Google Scholar] [CrossRef]
- Schlegel, N.C.; von Planta, A.; Widmer, D.S.; Dummer, R.; Christofori, G. PI3K signalling is required for a TGFβ-induced epithelial-mesenchymal-like transition (EMT-like) in human melanoma cells. Exp. Dermatol. 2015, 24, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Haeger, A.; Wolf, K.; Zegers, M.M.; Friedl, P. Collective cell migration: Guidance principles and hierarchies. Trends Cell Biol. 2015, 25, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Lou, E.; Gholami, S.; Romin, Y.; Thayanithy, V.; Fujisawa, S.; Desir, S.; Steer, C.J.; Subramanian, S.; Fong, Y.; Manova-Todorova, K.; et al. Imaging Tunneling Membrane Tubes Elucidates Cell Communication in Tumors. Trends Cancer 2017, 3, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Combedazou, A.; Choesmel-Cadamuro, V.; Gay, G.; Liu, J.; Dupré, L.; Ramel, D.; Wang, X. Myosin II governs collective cell migration behaviour downstream of guidance receptor signalling. J. Cell Sci. 2017, 130, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Iwanicki, M.P.; Davidowitz, R.A.; Ng, M.R.; Besser, A.; Muranen, T.; Merritt, M.; Danuser, G.; Ince, T.; Brugge, J.S. Ovarian cancer spheroids use myosin-generated force to clear the mesothelium. Cancer Discov. 2011, 1, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Wang, X.; Auwerx, J. Analysis of Mitochondrial Respiratory Chain Supercomplexes Using Blue Native Polyacrylamide Gel Electrophoresis (BN-PAGE). Curr. Protoc. Mouse Biol. 2016, 6, 1–14. [Google Scholar] [PubMed]
- Melzer, C.; von der Ohe, J.; Hass, R. Breast Carcinoma: From Initial Tumor Cell Detachment to Settlement at Secondary Sites. BioMed Res. Int. 2017, 2017, 8534371. [Google Scholar] [CrossRef]
- Desgrosellier, J.; David, C. Integrins in cancer: Biological implications in therapeutic opportunities. Cancer Nat. Rev. 2015, 10, 9–22. [Google Scholar] [CrossRef]
- Yilmaz, M.; Christofori, G. Mechanisms of Motility in Metastasizing Cells. Mol. Cancer Res. 2010, 8, 629–642. [Google Scholar] [CrossRef] [Green Version]
- Yuzhalin, A.E.; Gordon-Weeks, A.N.; Tognoli, M.L.; Jones, K.; Markelc, B.; Konietzny, R.; Fischer, R.; Muth, A.; O’Neill, E.; Thompson, P.R.; et al. Colorectal cancer liver metastatic growth depends on PAD4-driven citrullination of the extracellular matrix. Nat. Commun. 2018, 9, 4783. [Google Scholar] [CrossRef]
- Jia, D.; Jolly, M.K.; Kulkarni, P.; Levine, H. Phenotypic plasticity and cell fate decisions in cancer: Insights from dynamical systems theory. Cancers 2017, 9, 70. [Google Scholar] [CrossRef] [PubMed]
- Jolly, M.K.; Kulkarni, P.; Weninger, K.; Orban, J.; Levine, H. Phenotypic Plasticity, Bet-Hedging, and Androgen Independence in Prostate Cancer: Role of Non-Genetic Heterogeneity. Front. Oncol. 2018, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.-H.V.; Middleton, K.; You, L.; Sun, Y. A review of microfluidic approaches for investigating cancer extravasation during metastasis. Microsyst. Nanoeng. 2018, 4, 17104. [Google Scholar] [CrossRef] [Green Version]
- Kocal, G.C.; Güven, S.; Foygel, K.; Goldman, A.; Chen, P.; Sengupta, S.; Paulmurugan, R.; Baskin, Y.; Demirci, U. Dynamic Microenvironment Induces Phenotypic Plasticity of Esophageal Cancer Cells under Flow. Sci. Rep. 2016, 6, 38221. [Google Scholar] [CrossRef] [PubMed]
- Vedula, S.R.K.; Hirata, H.; Nai, M.H.; Brugués, A.; Toyama, Y.; Trepat, X.; Lim, C.T.; Ladoux, B. Epithelial bridges maintain tissue integrity during collective cell migration. Nat. Mater. 2014, 13, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Sarioglu, A.F.; Aceto, N.; Kojic, N.; Donaldson, M.C.; Zeinali, M.; Hamza, B.; Engstrom, A.; Zhu, H.; Sundaresan, T.K.; Miyamoto, D.T.; et al. A microfluidic device for label-free, physical capture of circulating tumor cell clusters. Nat. Methods 2015, 12, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Sandy, M.; Butler, A. Microbial iron acquisition: Marine and terrestrial siderophores. Chem. Rev. 2009, 109, 4580–4595. [Google Scholar] [CrossRef]
- Tanner, K.; Gottesman, M.M. Beyond 3D culture models of cancer. Sci. Transl. Med. 2015, 7, 283ps9. [Google Scholar] [CrossRef]
- Qiao, H.; Tang, T. Engineering 3D approaches to model the dynamic microenvironments of cancer bone metastasis. Bone Res. 2018, 6, 3. [Google Scholar] [CrossRef]
- Shang, M.; Soon, R.H.; Lim, C.T.; Khoo, B.L.; Han, J. Microfluidic modelling of the tumor microenvironment for anti-cancer drug development. Lab Chip 2019, 19, 369–386. [Google Scholar] [CrossRef]
- Truong, D.D.; Kratz, A.; Park, J.G.; Barrientos, E.S.; Saini, H.; Nguyen, T.; Pockaj, B.; Mouneimne, G.; LaBaer, J.; Nikkhah, M. A human organotypic microfluidic tumor model permits investigation of the interplay between patient-derived fibroblasts and breast cancer cells. Cancer Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ayuso, J.M.; Gillette, A.; Lugo-cintrón, K.; Acevedo-acevedo, S.; Gomez, I.; Morgan, M.; Heaster, T.; Wisinski, K.B.; Palecek, S.P.; Skala, M.C.; et al. EBioMedicine Organotypic micro fl uidic breast cancer model reveals starvation-induced spatial-temporal metabolic adaptations. EBioMedicine 2018, 37, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.S.; Schlenoff, J.B.; Keller, T.C.S. Collective epithelial cell sheet adhesion and migration on polyelectrolyte multilayers with uniform and gradients of compliance. Exp. Cell Res. 2016, 346, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Zajac, O.; Raingeaud, J.; Libanje, F.; Lefebvre, C.; Sabino, D.; Martins, I.; Roy, P.; Benatar, C.; Canet-Jourdan, C.; Azorin, P.; et al. Tumour spheres with inverted polarity drive the formation of peritoneal metastases in patients with hypermethylated colorectal carcinomas. Nat. Cell Biol. 2018, 20, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Wong, I.Y.; Javaid, S.; Wong, E.A.; Perk, S.; Haber, D.A.; Toner, M.; Irimia, D. Collective and individual migration following the epithelial-mesenchymal transition. Nat. Mater. 2014, 13, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.B.; ONeill, G.M. Cooperative cell invasion: Matrix metalloproteinase-mediated incorporation between cells. Mol. Biol. Cell 2016, 27, 3284–3292. [Google Scholar] [CrossRef]
- Klymenko, Y.; Johnson, J.; Bos, B.; Lombard, R.; Campbell, L.; Loughran, E.; Stack, M.S. Heterogeneous Cadherin Expression and Multicellular Aggregate Dynamics in Ovarian Cancer Dissemination. Neoplasia 2017, 19, 549–563. [Google Scholar] [CrossRef]
- Kilgore, J.A.; Dolman, N.J.; Davidson, M.W. A Review of Reagents for Fluorescence Microscopy of Cellular Compartments and Structures, Part II: Reagents for Non-Vesicular Organelles. In Current Protocols in Cytometry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 12.31.1–12.31.24. ISBN 0471142956. [Google Scholar]
- Kilgore, J.A.; Dolman, N.J.; Davidson, M.W. A Review of Reagents for Fluorescence Microscopy of Cellular Compartments and Structures, Part III: Reagents for Actin, Tubulin, Cellular Membranes, and Whole Cell and Cytoplasm. In Current Protocols in Cytometry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 12.32.1–12.32.17. ISBN 0471142956. [Google Scholar]
- Marsh, E.; Gonzalez, D.G.; Lathrop, E.A.; Boucher, J.; Marsh, E.; Gonzalez, D.G.; Lathrop, E.A.; Boucher, J.; Greco, V. Positional Stability and Membrane Occupancy Define Skin Fibroblast Homeostasis In Vivo Article Positional Stability and Membrane Occupancy Define Skin Fibroblast Homeostasis In Vivo. Cell 2018, 175, 1620–1633. [Google Scholar] [CrossRef]
- Chen, J.; Landegger, L.D.; Sun, Y.; Ren, J.; Maimon, N.; Wu, L.; Ng, M.R.; Chen, J.W.; Zhang, N.; Zhao, Y.; et al. A cerebellopontine angle mouse model for the investigation of tumor biology, hearing, and neurological function in NF2-related vestibular schwannoma. Nat. Protoc. 2019, 14, 541–555. [Google Scholar] [CrossRef]
- Harper, K.L.; Sosa, M.S.; Entenberg, D.; Hosseini, H.; Cheung, J.F.; Nobre, R.; Avivar-Valderas, A.; Nagi, C.; Girnius, N.; Davis, R.J.; et al. Mechanism of early dissemination and metastasis in Her2+ mammary cancer. Nature 2016, 540, 588–592. [Google Scholar] [CrossRef]
- Yang, Z.; Hoffman, R.M.; Ma, H.; Toneri, M.; Goto, Y.; Zhang, Y.; Bouvet, M.; Seki, N. Real-Time GFP Intravital Imaging of the Differences in Cellular and Angiogenic Behavior of Subcutaneous and Orthotopic Nude-Mouse Models of Human PC-3 Prostate Cancer. J. Cell. Biochem. 2016, 117, 2546–2551. [Google Scholar]
- Zomer, A.; Ellenbroek, S.I.J.; Ritsma, L.; Beerling, E.; Vrisekoop, N.; Van Rheenen, J. Brief report: Intravital imaging of cancer stem cell plasticity in mammary tumors. Stem Cells 2013, 31, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Ritsma, L.; Ellenbroek, S.I.J.; Zomer, A.; Snippert, H.J.; de Sauvage, F.J.; Simons, B.D.; Clevers, H.; van Rheenen, J. Intestinal crypt homeostasis revealed at single-stem-cell level by in vivo live imaging. Nature 2014, 507, 362–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, M.Q.; Kandyba, E.; Harris, S.; Del Rosario, R.; Balmain, A. Multicolour lineage tracing reveals clonal dynamics of squamous carcinoma evolution from initiation to metastasis. Nat. Cell Biol. 2018, 20, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.; Park, M.; Watson, S.L.; Wakefield, D.; Di Girolamo, N. Visualizing the Fate of Transplanted K14-Confetti Corneal Epithelia in a Mouse Model of Limbal Stem Cell Deficiency. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1630–1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobis, M.; Warren, S.C.; Lucas, M.C.; Murphy, K.J.; Herrmann, D.; Timpson, P. Molecular mobility and activity in an intravital imaging setting—Implications for cancer progression and targeting. J. Cell Sci. 2018, 131, jcs206995. [Google Scholar] [CrossRef]
- Chitty, J.L.; Filipe, E.C.; Lucas, M.C.; Herrmann, D.; Cox, T.R.; Timpson, P. Recent advances in understanding the complexities of metastasis. F1000Research 2018, 7, F1000. [Google Scholar] [CrossRef]
- Liu, T.-L.; Upadhyayula, S.; Milkie, D.E.; Singh, V.; Wang, K.; Swinburne, I.A.; Mosaliganti, K.R.; Collins, Z.M.; Hiscock, T.W.; Shea, J.; et al. Observing the cell in its native state: Imaging subcellular dynamics in multicellular organisms. Science 2018, 360, eaaq1392. [Google Scholar] [CrossRef]
- Labernadie, A.; Trepat, X. ScienceDirect Sticking, steering, squeezing and shearing: Cell movements driven by heterotypic mechanical forces. Curr. Opin. Cell Biol. 2018, 54, 57–65. [Google Scholar] [CrossRef]
- Kelley, L.C.; Wang, Z.; Hagedorn, E.J.; Wang, L.; Shen, W.; Lei, S.; Johnson, S.A.; Sherwood, D.R. Live-cell confocal microscopy and quantitative 4D image analysis of anchor-cell invasion through the basement membrane in Caenorhabditis elegans. Nat. Protoc. 2017, 12, 2081–2096. [Google Scholar] [CrossRef] [Green Version]
- Chantzi, E.; Jarvius, M.; Niklasson, M.; Segerman, A.; Gustafsson, M.G. COMBImage: A modular parallel processing framework for pairwise drug combination analysis that quantifies temporal changes in label-free video microscopy movies. BMC Bioinform. 2018, 19, 453. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.F.Z.; Fernandez-Gonzalez, R. (Machine-)Learning to analyze in vivo microscopy: Support vector machines. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 1719–1727. [Google Scholar] [CrossRef] [PubMed]
- Svensson, C.; Medyukhina, A.; Belyaev, I.; Al-Zaben, N.; Figge, M.T. Untangling cell tracks: Quantifying cell migration by time lapse image data analysis. Cytom. Part A 2018, 93, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Valm, A.M.; Lippincott-Schwartz, J. Multispectral Live-Cell Imaging. Curr. Protoc. Cell Biol. 2018, 79, e46. [Google Scholar] [CrossRef] [PubMed]
- Piltti, K.M.; Cummings, B.J.; Carta, K.; Manughian-Peter, A.; Worne, C.L.; Singh, K.; Ong, D.; Maksymyuk, Y.; Khine, M.; Anderson, A.J. Live-cell time-lapse imaging and single-cell tracking of in vitro cultured neural stem cells—Tools for analyzing dynamics of cell cycle, migration, and lineage selection. Methods 2018, 133, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Correa-Gallegos, D.; Christ, S.; Stefanska, A.; Liu, J.; Ramesh, P.; Rajendran, V.; De Santis, M.M.; Wagner, D.E.; Rinkevich, Y. Two succeeding fibroblastic lineages drive dermal development and the transition from regeneration to scarring. Nat. Cell Biol. 2018, 20, 422–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Guo, P.; Moses, M.A. A time-lapse, label-free, quantitative phase imaging study of dormant and active human cancer cells. J. Vis. Exp. 2018, 132, e57035. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.M.; Stuelten, C.H.; Parent, C.A.; Losert, W. Collective cell migration over long time scales reveals distinct phenotypes. Converg. Sci. Phys. Oncol. 2016, 2, 025001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chepizhko, O.; Giampietro, C.; Mastrapasqua, E.; Nourazar, M.; Ascagni, M.; Sugni, M.; Fascio, U.; Leggio, L.; Malinverno, C.; Scita, G.; et al. Bursts of activity in collective cell migration. Proc. Natl. Acad. Sci. USA 2016, 113, 11408–11413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, C.D.; Mistriotis, P.; Konstantopoulos, K. Cancer cell motility: Lessons from migration in confined spaces. Nat. Rev. Cancer 2017, 17, 131–140. [Google Scholar] [CrossRef]
- Rogers, S.; McCloy, R.A.; Parker, B.L.; Gallego-Ortega, D.; Law, A.M.K.; Chin, V.T.; Conway, J.R.W.; Fey, D.; Millar, E.K.A.; O’Toole, S.; et al. MASTL overexpression promotes chromosome instability and metastasis in breast cancer. Oncogene 2018, 37, 4518–4533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetmanski, J.H.R.; Zindy, E.; Schwartz, J.M.; Caswell, P.T. A MAPK-Driven Feedback Loop Suppresses Rac Activity to Promote RhoA-Driven Cancer Cell Invasion. PLoS Comput. Biol. 2016, 12, e1004909. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.; Kneissl, J.; Grabher-Meier, V.; Heindl, S.; Hasenauer, J.; Maier, D.; Mattes, J.; Winter, P.; Luber, B. Evaluation of epidermal growth factor receptor signaling effects in gastric cancer cell lines by detailed motility-focused phenotypic characterization linked with molecular analysis. BMC Cancer 2017, 17, 845. [Google Scholar] [CrossRef] [PubMed]
- Shafqat-Abbasi, H.; Kowalewski, J.M.; Kiss, A.; Gong, X.; Hernandez-Varas, P.; Berge, U.; Jafari-Mamaghani, M.; Lock, J.G.; Strömblad, S. An analysis toolbox to explore mesenchymal migration heterogeneity reveals adaptive switching between distinct modes. Elife 2016, 5, e11384. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Taftaf, R.; Kawaguchi, M.; Chang, Y.F.; Chen, W.; Entenberg, D.; Zhang, Y.; Gerratana, L.; Huang, S.; Patel, D.B.; et al. Homophilic CD44 interactions mediate tumor cell aggregation and polyclonal metastasis in patient-derived breast cancer models. Cancer Discov. 2019, 9, 96–113. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Functional assays for the metastatic cascade. Metastasis encompasses three distinct stages, viz. primary dissociation, physical translocation, and colonization. The interplay of complex processes severs cells from the primary tumor; these cells proliferate, migrate, and invade through the tissue matrix to initiate hematogenous or lymphatic dissemination. Circulating tumor cells then overcome hydrostatic pressures and immune surveillance to extravasate and colonize distant tissues to seed micro-/macro-metastases. Diverse cellular functions activated during the metastatic cascade are evaluated experimentally by functional assays, and can be modified to accommodate multiple biological components (micro-environmental milieu, extra-cellular matrix–ECM, stromal cells, extrinsic physical pressures, immune cells, and so on). A list of the relevant assays employed across the metastatic cascade are listed and indicated in the schematic.
Figure 1.
Functional assays for the metastatic cascade. Metastasis encompasses three distinct stages, viz. primary dissociation, physical translocation, and colonization. The interplay of complex processes severs cells from the primary tumor; these cells proliferate, migrate, and invade through the tissue matrix to initiate hematogenous or lymphatic dissemination. Circulating tumor cells then overcome hydrostatic pressures and immune surveillance to extravasate and colonize distant tissues to seed micro-/macro-metastases. Diverse cellular functions activated during the metastatic cascade are evaluated experimentally by functional assays, and can be modified to accommodate multiple biological components (micro-environmental milieu, extra-cellular matrix–ECM, stromal cells, extrinsic physical pressures, immune cells, and so on). A list of the relevant assays employed across the metastatic cascade are listed and indicated in the schematic.

Figure 2.
Modalities of metastasis. Metastasis is achieved by a step wise progression of tumor cell dissociation, physical translocation, and colonization. (a.) Cell dissociation entails a combination of cell–cell junctional complex rearrangements; phenotypic transitions, like EMT, that result in a loss of epithelial junctions; or the passive severance of cell clusters by virtue of forces exerted by the interstitial fluids. (b.i.) Physical translocation involves a myriad of processes, amongst which cell migration is mediated by individual or collective clusters of cells. Individual cells migrate with the aid of ECM or stroma derived nanotubes, whereas collective migration is achieved by an active leading edge and/or by migration of cell sheets. (b.ii.) Migrating cells then undergo intra- and trans-tissue invasion by mediating distinct interactions with the ECM and non-transformed cell populations. Intra-tissue invasion involves the degradation and deformation of the ECM, either by the tumor or stromal cells. Alternatively, cells exhibit amoeboid invasion by altering their membrane fluidity so as to squeeze through the ECM with minimal disturbance to the surrounding architecture. (b.iii.) Trans-tissue invasion involves the disruption of endothelial linings by virtue of active intra-/extra-vasation or mechanical rupture because of an extensive tumor load. (b.iv.) Disseminating tumor cells in circulatory/lymphatic systems can exist either as platelet or tumor associated macrophage (TAMs) coated entities, mesenchymal cells along capillary linings, passively dispersed cell clusters, or exhibit an integrin mediated tumbling similar to cells of the immune system. (c.) The final stage of metastases establishment involves the colonization of tumor cells at distant sites mediated by phenotypic transitions, like mesenchymal to epithelial transition (MET), the selective seeding of epithelial cells from heterogeneous clusters, paracrine transformation of secondary site by the tumor cell secretome, passive deposition by interstitial fluids, and modulation of the secondary niche by activation of the tissue stromal compartment.
Figure 2.
Modalities of metastasis. Metastasis is achieved by a step wise progression of tumor cell dissociation, physical translocation, and colonization. (a.) Cell dissociation entails a combination of cell–cell junctional complex rearrangements; phenotypic transitions, like EMT, that result in a loss of epithelial junctions; or the passive severance of cell clusters by virtue of forces exerted by the interstitial fluids. (b.i.) Physical translocation involves a myriad of processes, amongst which cell migration is mediated by individual or collective clusters of cells. Individual cells migrate with the aid of ECM or stroma derived nanotubes, whereas collective migration is achieved by an active leading edge and/or by migration of cell sheets. (b.ii.) Migrating cells then undergo intra- and trans-tissue invasion by mediating distinct interactions with the ECM and non-transformed cell populations. Intra-tissue invasion involves the degradation and deformation of the ECM, either by the tumor or stromal cells. Alternatively, cells exhibit amoeboid invasion by altering their membrane fluidity so as to squeeze through the ECM with minimal disturbance to the surrounding architecture. (b.iii.) Trans-tissue invasion involves the disruption of endothelial linings by virtue of active intra-/extra-vasation or mechanical rupture because of an extensive tumor load. (b.iv.) Disseminating tumor cells in circulatory/lymphatic systems can exist either as platelet or tumor associated macrophage (TAMs) coated entities, mesenchymal cells along capillary linings, passively dispersed cell clusters, or exhibit an integrin mediated tumbling similar to cells of the immune system. (c.) The final stage of metastases establishment involves the colonization of tumor cells at distant sites mediated by phenotypic transitions, like mesenchymal to epithelial transition (MET), the selective seeding of epithelial cells from heterogeneous clusters, paracrine transformation of secondary site by the tumor cell secretome, passive deposition by interstitial fluids, and modulation of the secondary niche by activation of the tissue stromal compartment.

Figure 3.
Era of real-time analysis. Routine implementation of real-time approaches in cancer biology can spatio-temporally resolve cellular processes and biological modalities by permitting the extraction of quantitative metrics associated with these states. Outputs from real-time studies can be collated to identify the regulatory networks governing the biological processes, and can be further applied to develop analytical tools/mathematical models for implementation in therapeutic/clinical screens. Such analyses support the design of relevant animal models, recapitulating the in situ patho-physiological parameters of the disease. Corroboration with microfluidic devices further enhances the outputs of these pipelines.
Figure 3.
Era of real-time analysis. Routine implementation of real-time approaches in cancer biology can spatio-temporally resolve cellular processes and biological modalities by permitting the extraction of quantitative metrics associated with these states. Outputs from real-time studies can be collated to identify the regulatory networks governing the biological processes, and can be further applied to develop analytical tools/mathematical models for implementation in therapeutic/clinical screens. Such analyses support the design of relevant animal models, recapitulating the in situ patho-physiological parameters of the disease. Corroboration with microfluidic devices further enhances the outputs of these pipelines.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Varankar, S.S.; Bapat, S.A. Uncoupling Traditional Functionalities of Metastasis: The Parting of Ways with Real-Time Assays. J. Clin. Med. 2019, 8, 941. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8070941
AMA Style
Varankar SS, Bapat SA. Uncoupling Traditional Functionalities of Metastasis: The Parting of Ways with Real-Time Assays. Journal of Clinical Medicine. 2019; 8(7):941. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8070941
Chicago/Turabian StyleVarankar, Sagar S., and Sharmila A. Bapat. 2019. "Uncoupling Traditional Functionalities of Metastasis: The Parting of Ways with Real-Time Assays" Journal of Clinical Medicine 8, no. 7: 941. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8070941
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.