Soil Respiration Dynamics in Bromus erectus-Dominated Grasslands under Different Management Intensities

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
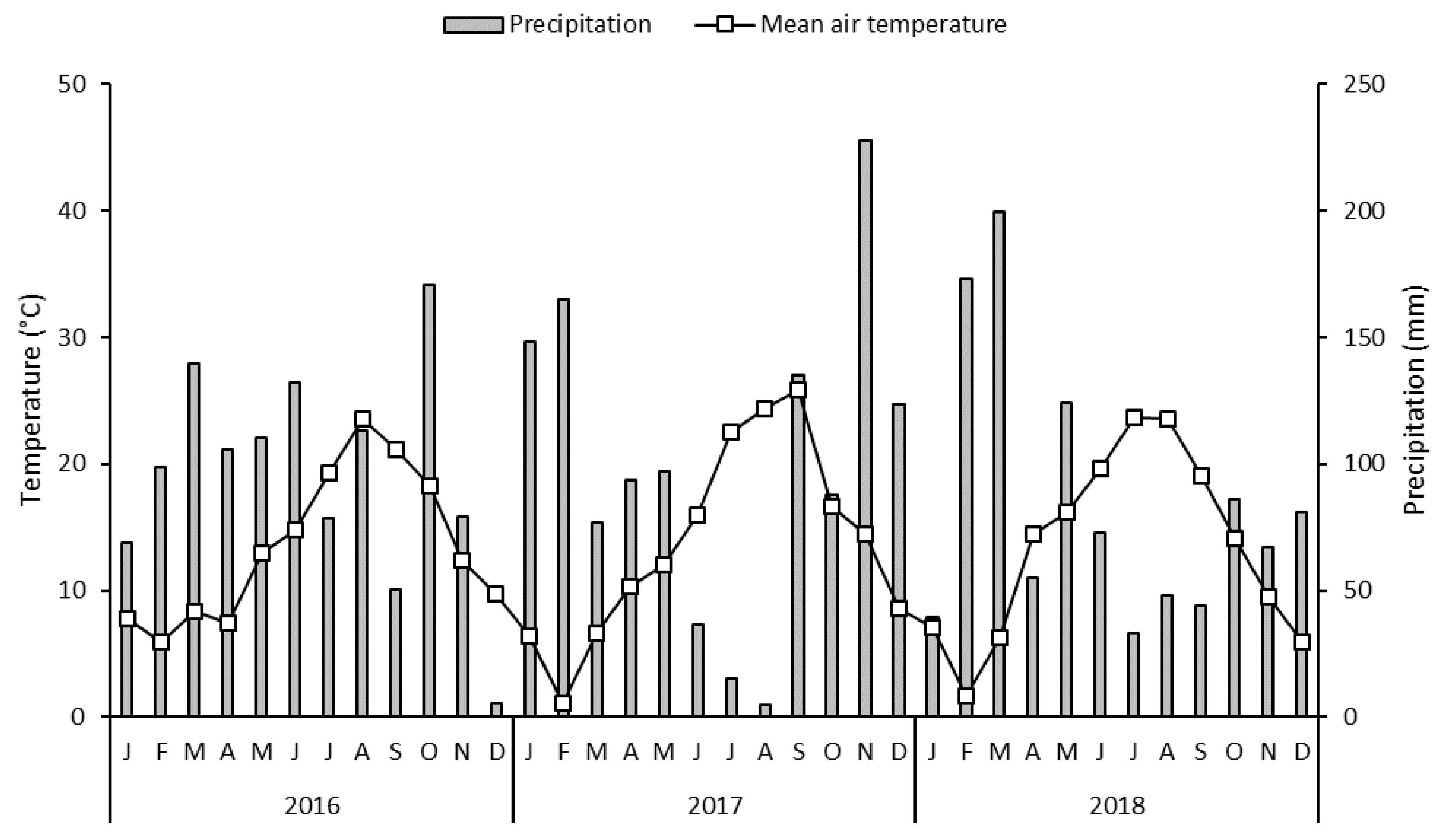
2.1. Study Area
2.2. Study Site and Experimental Design
2.3. Measurement Descriptions
2.4. Data Analysis
3. Results
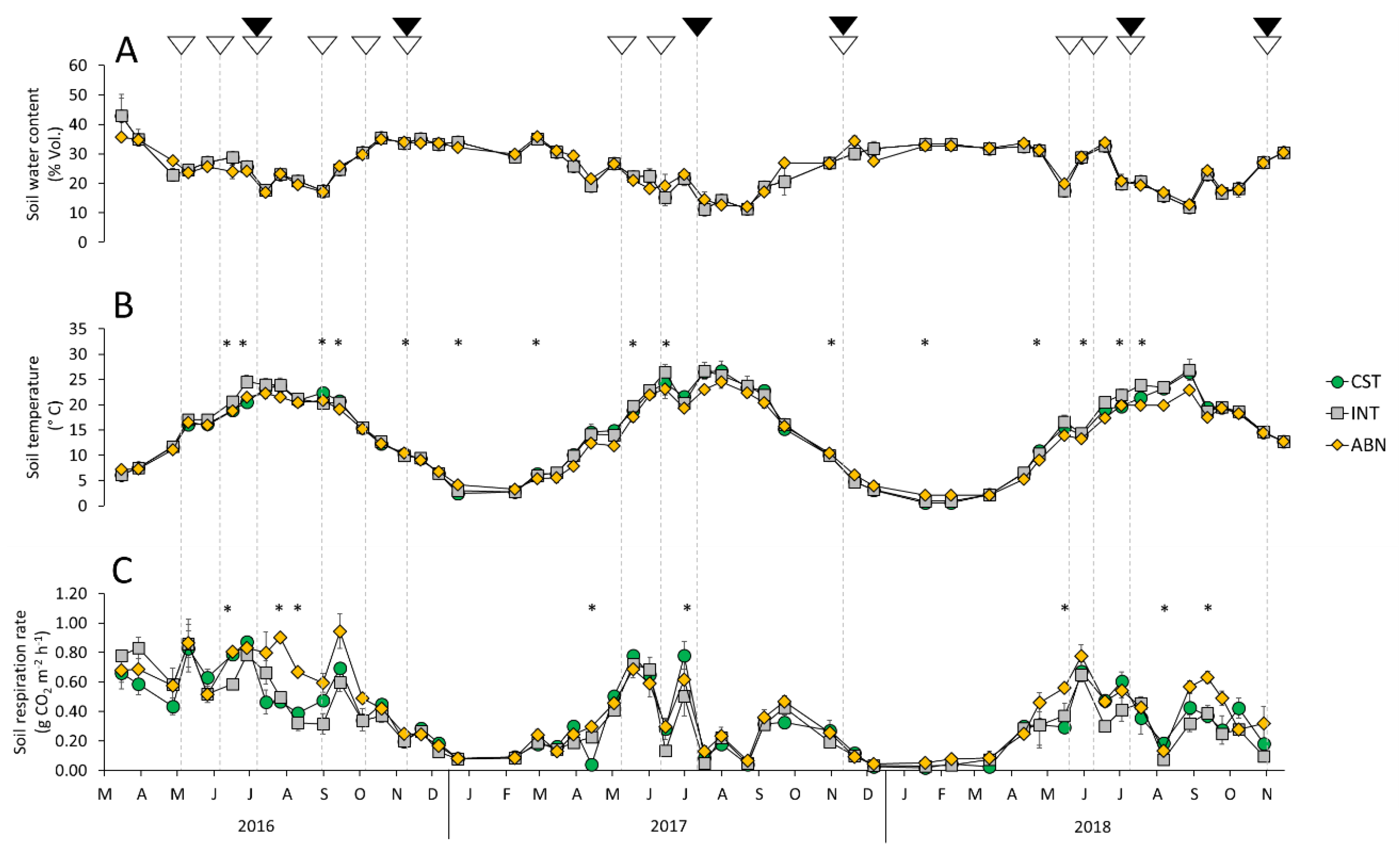
3.1. Soil Water Content and Temperature Dynamics as Affected by Mowing Intensity
3.2. Soil Respiration Dynamics as Affected by Mowing Intensity
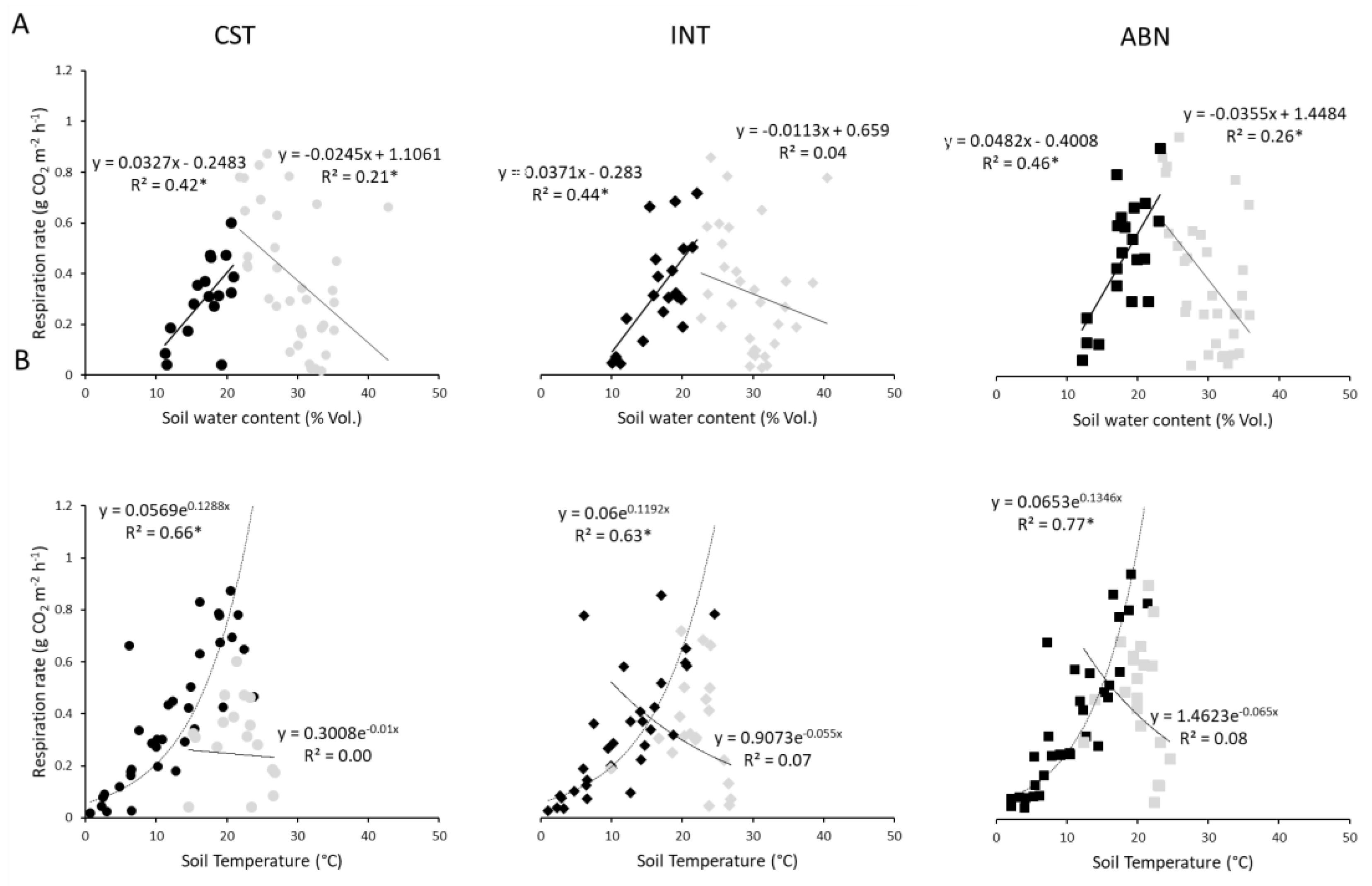
3.3. Relationships Between Soil Respiration, Soil Water Content, and Temperature
4. Discussion
4.1. Relationships Between Soil Respiration, Water Content and Temperature
4.2. The Effects of Management Practices on Soil Water Content and Temperature
4.3. The Effects of Management Practices on Soil Respiration
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, P.; Bustamante, M.; Ahammad, H.; Clark, H.; Dong, H.; Elsiddig, E.A.; Haberl, H.; Harper, R.; House, J.; Jafari, M.; et al. Agriculture, forestry and other land use (AFOLU). In Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 1992, 44, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Shang, H. Contribution of soil respiration to the global carbon equation. J. Plant Physiol. 2016, 203, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Rustad, L.; Huntington, T.G.; States, U.; Survey, G.; Boone, R.D. Controls on Soil Respiration: Implications for Climate Change Controls on soil respiration: Implications for climate change. Biogeochemistry 2000, 48, 1–6. [Google Scholar] [CrossRef]
- Hopkins, A.; Del Prado, A. Implications of climate change for grassland in Europe: Impacts, adaptations and mitigation options: A review. Grass Forage Sci. 2007, 62, 118–126. [Google Scholar] [CrossRef]
- Wang, W.; Fang, J. Soil respiration and human effects on global grasslands. Glob. Planet. Chang. 2009, 67, 20–28. [Google Scholar] [CrossRef]
- O’Mara, F.P. The role of grasslands in food security and climate change. Ann. Bot. 2012, 110, 1263–1270. [Google Scholar] [CrossRef] [Green Version]
- Smith, P. Land use change and soil organic carbon dynamics. Nutr. Cycl. Agroecosyst. 2008, 81, 169–178. [Google Scholar] [CrossRef]
- Li, W.; Wang, J.; Zhang, X.; Shi, S.; Cao, W. Effect of degradation and rebuilding of artificial grasslands on soil respiration and carbon and nitrogen pools on an alpine meadow of the Qinghai-Tibetan Plateau. Ecol. Eng. 2018, 111, 134–142. [Google Scholar]
- Hoffmann, I.; From, T.; Boerma, D. Ecosystem Services Provided by Livestock Species and Breeds, with Special Consideration to the Contributions of Small-Scale Livestock Keepers and Pastoralists. FAO Commission on Genetic Resources for Food and Agriculture. Background Study Paper 66(1). 2014. Available online: http://www.fao.org/3/a-at598e.pdf (accessed on 27 July 2019).
- Velthof, G.L.; Lesschen, J.P.; Schils, R.L.M.; Smit, A.; Elbersen, B.S.; Hazeu, G.W.; Mucher, C.A.; Oenema, O. Grassland Areas, Production and Use; Alterra Wageningen UR: Wageningen, The Netherlands, 2014. [Google Scholar]
- Almagro, M.; López, J.; Querejeta, J.I.; Martínez-Mena, M. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biol. Biochem. 2009, 41, 594–605. [Google Scholar] [CrossRef]
- Rong, Y.; Ma, L.; Johnson, D.A.; Yuan, F. Soil respiration patterns for four major land-use types of the agro-pastoral region of northern China. Agric. Ecosyst. Environ. 2015, 213, 142–150. [Google Scholar] [CrossRef]
- Calaciura, B.; Spinelli, O. Management of Natura 2000 Habitats. In 6210 Semi-Natural Dry Grasslands and Scrubland Facies on Calcareous Substrates (Festuco-Brometalia) (* Important Orchid Sites); European Commission: Brussels, Belgium, 2008; ISBN 978-92-79-08326-6. [Google Scholar]
- Wilson, J.W.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant species richness: The world records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- D’Ottavio, P.; Francioni, M.; Trozzo, L.; Sedić, E.; Budimir, K.; Avanzolini, P.; Trombetta, M.F.; Porqueddu, C.; Santilocchi, R.; Toderi, M. Trends and approaches in the analysis of ecosystem services provided by grazing systems: A review. Grass Forage Sci. 2018, 73, 15–25. [Google Scholar] [CrossRef]
- Alcamo, J.; Ash, N.J.; Butler, C.D.; Callicot, J.B.; Capistrano, D.; Carpenter, S.R. Ecosystems and Human Well-Being: A Framework for Assessment; Island Press: Washington, DC, USA, 2003; ISBN 1597260401. [Google Scholar]
- Toderi, M.; Francioni, M.; Seddaiu, G.; Roggero, P.P.; Trozzo, L.; D’Ottavio, P. Bottom-up design process of agri-environmental measures at a landscape scale: Evidence from case studies on biodiversity conservation and water protection. Land Use Policy 2017, 68, 295–305. [Google Scholar] [CrossRef]
- D’Ottavio, P.; Ziliotto, U. Effect of different management on the production characteristics of mountain permanent meadows. Ital. J. Anim. Sci. 2003, 2, 249–251. [Google Scholar]
- Bonanomi, G.; Caporaso, S.; Allegrezza, M. Effects of nitrogen enrichment, plant litter removal and cutting on a species-rich Mediterranean calcareous grassland. Plant Biosyst. 2009, 143, 443–455. [Google Scholar] [CrossRef]
- Tesei, G.; D’Ottavio, P.; Toderi, M.; Ottaviani, C.; Pesaresi, S.; Francioni, M.; Trozzo, L.; Allegrezza, M. Restoration strategies for grasslands colonized by Asphodel-dominant communities. Grassl. Sci. 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bahn, M.; Rodeghiero, M.; Anderson-Dunn, M.; Dore, S.; Gimeno, C.; Drösler, M.; Williams, M.; Ammann, C.; Berninger, F.; Flechard, C.; et al. Soil respiration in European grasslands in relation to climate and assimilate supply. Ecosystems 2008, 11, 1352–1367. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Ren, C.; Shelton, S.; Wang, Z.; Pang, G.; Chen, J.; Wang, J. Grazing intensity influence soil microbial communities and their implications for soil respiration. Agric. Ecosyst. Environ. 2017, 249, 50–56. [Google Scholar] [CrossRef]
- Wei, L.; Liu, J.; Su, J.; Jing, G.; Zhao, J.; Cheng, J.; Jin, J. Effect of clipping on soil respiration components in temperate grassland of Loess Plateau. Eur. J. Soil Biol. 2016, 75, 157–167. [Google Scholar] [CrossRef]
- Wan, S.; Luo, Y. Substrate regulation of soil respiration in a tallgrass prairie: Results of a clipping and shading experiment. Glob. Biogeochem. Cycles 2003, 17, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Zhang, Z.; Wang, C.; Jiang, F.; Xia, J. Effects of mowing and nitrogen addition on soil respiration in three patches in an oldfield grassland in Inner Mongolia. J. Plant Ecol. 2011, 5, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Luo, C.; Wang, S.; Zhang, Z.; Cui, S.; Bao, X.; Jiang, L.; Li, Y.; Li, X.; Wang, Q.; et al. Effects of warming, grazing/cutting and nitrogen fertilization on greenhouse gas fluxes during growing seasons in an alpine meadow on the Tibetan Plateau. Agric. For. Meteorol. 2015, 214–215, 506–514. [Google Scholar] [CrossRef]
- Francioni, M.; D’Ottavio, P.; Lai, R.; Trozzo, L.; Budimir, K.; Foresi, L.; Kishimoto-Mo, A.W.; Baldoni, N.; Allegrezza, M.; Tesei, G.; et al. Seasonal soil respiration Dynamics and Carbon-Stock Variations in Mountain Permanent Grasslands Compared to Arable Lands. Agriculture 2019, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Gavrichkova, O.; Moscatelli, M.C.; Kuzyakov, Y.; Grego, S.; Valentini, R. Influence of defoliation on CO2 efflux from soil and microbial activity in a Mediterranean grassland. Agric. Ecosyst. Environ. 2010, 136, 87–96. [Google Scholar] [CrossRef]
- Bahn, M.; Knapp, M.; Garajova, Z.; Pfahringer, N.; Cernusca, A. Root respiration in temperate mountain grasslands differing in land use. Glob. Chang. Biol. 2006, 12, 995–1006. [Google Scholar] [CrossRef]
- Cherwin, K.; Knapp, A. Unexpected patterns of sensitivity to drought in three semi-arid grasslands. Oecologia 2012, 169, 845–852. [Google Scholar] [CrossRef]
- Zhu, L.; Johnson, D.A.; Wang, W.; Ma, L.; Rong, Y. Grazing effects on carbon fluxes in a Northern China grassland. J. Arid Environ. 2015, 114, 41–48. [Google Scholar] [CrossRef]
- Caballero, R.; Fernández-gonzález, F.; Badia, R.; Molle, G.; Roggero, P.; Bagella, S.; D’Ottavio, P.; Papanastasis, V.; Fotiadis, G.; Sidiropoulou, A.; et al. Grazing Systems and Biodiversity in Mediterranean Areas: Spain, Italy and Greece. Pastos 2009, 39, 9–154. [Google Scholar]
- Allegrezza, M. Vegetazione e paesaggio vegetale della dorsale del Monte San Vicino (Appennino centrale). Fitosociologia 2003, 40, 3–118. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; Soil Conservation Services: Washington, DC, USA, 2014; Volume 12, p. 410.
- Jian, J.; Steele, M.K.; Day, S.D.; Quinn Thomas, R.; Hodges, S.C. Measurement strategies to account for soil respiration temporal heterogeneity across diverse regions. Soil Biol. Biochem. 2018, 125, 167–177. [Google Scholar] [CrossRef]
- Volpi, I.; Laville, P.; Bonari, E.; Nassi o Di Nasso, N.; Bosco, S. Nitrous oxide mitigation potential of reduced tillage and N input in durum wheat in the Mediterranean. Nutr. Cycl. Agroecosyst. 2018, 111, 189–201. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Soussana, J.; Loiseau, P.; Vuichard, N.; Ceschia, E.; Balesdent, J.; Chevallier, T.; Arrouays, D. Carbon cycling and sequestration opportunities in temperate grasslands. Soil Use Manag. 2004, 20, 219–230. [Google Scholar] [CrossRef]
- Balogh, J.; Papp, M.; Pintér, K.; Fóti, S.; Posta, K.; Eugster, W.; Nagy, Z. Autotrophic component of soil respiration is repressed by drought more than the heterotrophic one in dry grasslands. Biogeosciences 2016, 13, 5171–5182. [Google Scholar] [CrossRef] [Green Version]
- Francioni, M.; Lai, R.; D’Ottavio, P.; Trozzo, L.; Kishimoto-Mo, A.W.; Budimir, K.; Baldoni, N.; Toderi, M. Soil respiration dynamics in forage-based and cereal-based cropping systems in central Italy. Sci. Agric. 2020, 77, 1–10. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Z.; Wan, S. Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Glob. Chang. Biol. 2009, 15, 184–195. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | Source of Variation | p Values | Mowing Regime | Seasonal Mean ± Standard Error | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | 2016 | 2017 | 2018 | |||||||
| G | S | G | S | G | G | S | G | S | G | |||
| Soil water content (% Vol.) | Time | 0.01 | 0.01 | 0.01 | 0.06 | 0.01 | CST | 26.91 ± 0.49 | 32.11 ± 0.22 | 19.28 ± 0.88 | 31.92 ± 0.44 | 22.62 ± 0.63 |
| Mowing | 0.43 | 0.72 | 0.55 | 0.34 | 0.20 | INT | 26.13 ± 0.81 | 30.89 ± 0.70 | 19.29 ± 0.29 | 30.95 ± 0.41 | 22.21 ± 0.20 | |
| Time × Mowing | 0.49 | 0.30 | 0.29 | 0.08 | 0.95 | ABN | 25.94 ± 0.51 | 32.47 ± 0.50 | 19.98 ± 0.41 | 32.05 ± 0.61 | 23.16 ± 0.59 | |
| Soil temperature (°C) | Time | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | CST | 16.85 ± 0.52 | 6.77 ± 0.31 | 20.10 ± 0.85 ab | 3.45 ± 0.21 | 18.07 ± 0.60 |
| Mowing | 0.13 | 0.73 | 0.01 | 0.16 | 0.10 | INT | 17.34 ± 0.53 | 6.76 ± 0.50 | 20.18 ± 0.95 a | 3.54 ± 0.19 | 18.64 ± 0.83 | |
| Time × Mowing | 0.01 | 0.01 | 0.01 | 0.01 | 0.02 | ABN | 16.44 ± 0.28 | 6.58 ± 0.27 | 18.57 ± 0.73 b | 3.92 ± 0.05 | 16.80 ± 0.25 | |
| Soil respiration (g CO2 m−2 h−1) | Time | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | CST | 0.56 ± 0.01 ab | 0.19 ± 0.02 | 0.35 ± 0.02 | 0.05 ± 0.01 | 0.37 ± 0.05 |
| Mowing | 0.04 | 0.40 | 0.28 | 0.23 | 0.23 | INT | 0.54 ± 0.01 b | 0.16 ± 0.01 | 0.33 ± 0.02 | 0.06 ± 0.01 | 0.32 ± 0.04 | |
| Time × Mowing | 0.01 | 0.08 | 0.01 | 0.82 | 0.01 | ABN | 0.67 ± 0.05 a | 0.18 ± 0.01 | 0.37 ± 0.03 | 0.07 ± 0.01 | 0.45 ± 0.01 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francioni, M.; Trozzo, L.; Toderi, M.; Baldoni, N.; Allegrezza, M.; Tesei, G.; Kishimoto-Mo, A.W.; Foresi, L.; Santilocchi, R.; D’Ottavio, P. Soil Respiration Dynamics in Bromus erectus-Dominated Grasslands under Different Management Intensities. Agriculture 2020, 10, 9. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10010009
Francioni M, Trozzo L, Toderi M, Baldoni N, Allegrezza M, Tesei G, Kishimoto-Mo AW, Foresi L, Santilocchi R, D’Ottavio P. Soil Respiration Dynamics in Bromus erectus-Dominated Grasslands under Different Management Intensities. Agriculture. 2020; 10(1):9. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10010009
Chicago/Turabian StyleFrancioni, Matteo, Laura Trozzo, Marco Toderi, Nora Baldoni, Marina Allegrezza, Giulio Tesei, Ayaka Wenhong Kishimoto-Mo, Lucia Foresi, Rodolfo Santilocchi, and Paride D’Ottavio. 2020. "Soil Respiration Dynamics in Bromus erectus-Dominated Grasslands under Different Management Intensities" Agriculture 10, no. 1: 9. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10010009