The Impact of Early Basal Leaf Removal at Different Sides of the Canopy on Aglianico Grape Quality




Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Grapevine Measurements
- N: 100% of fruit-zone leaves were removed from the north canopy side (bunches exposed to the afternoon sun);
- S: 100% of fruit-zone leaves were removed from the south canopy side (bunches exposed to the morning sun;
- N–S: 100% of fruit-zone leaves were removed from the north and the south sides of the canopy (bunches exposed to the morning and afternoon sun), respectively;
- A control thesis (non-defoliated vines, where all basal leaves were retained in each shoot) was used for comparison.
2.2. Yield Components and Grape Composition
2.3. Phenols Extraction
2.4. Phenols and Antioxidant Activity Analysis
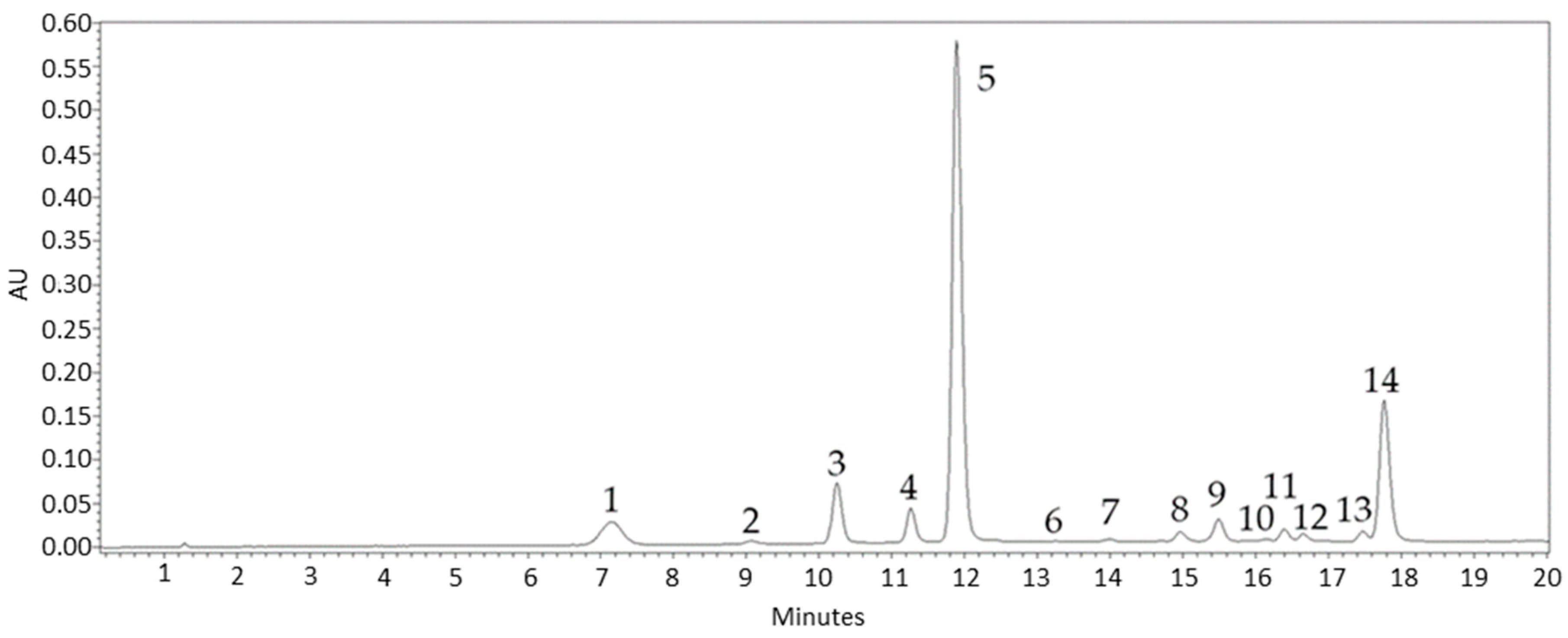
2.5. HPLC-DAD Anthocyanin Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. Growing Season Environmental Conditions and Grape Yields
3.2. Berry Composition
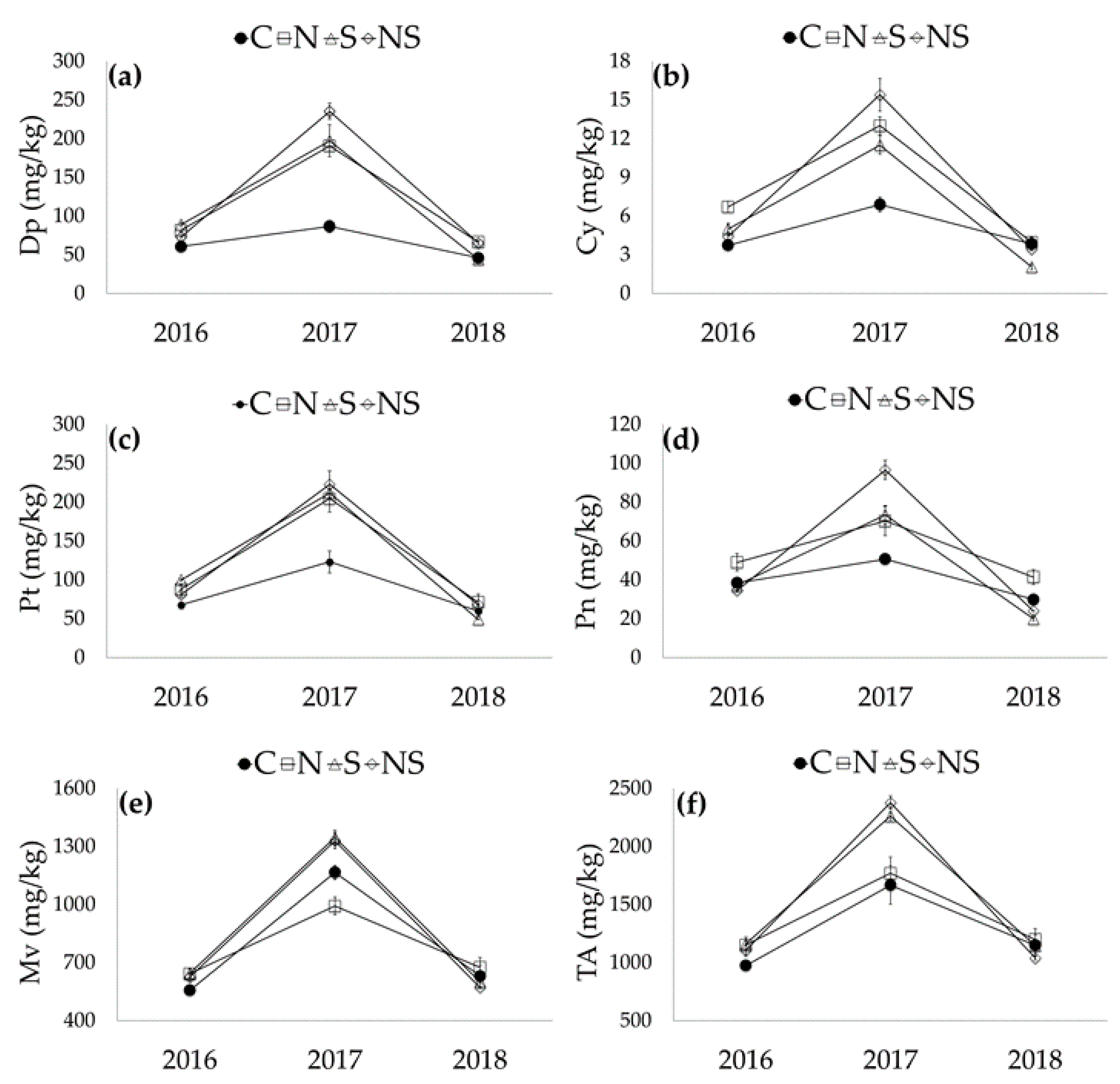
3.3. Anthocyanin Profile
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bergqvist, J.; Dokoozlian, N.; Ebisuda, N. Sunlight exposure and temperature effects on berry growth and composition of Cabernet Sauvignon and Grenache in the Central San Joaquin Valley of California. Am. J. Enol. Vitic. 2001, 52, 1–7. [Google Scholar]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J.C. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar]
- Drappier, J.; Thibon, C.; Rabot, A.; Geny-Denis, L. Relationship between wine composition and temperature: Impact on Bordeaux wine typicity in the context of global warming. Crit. Rev. Food Sci. Nutr. 2019, 59, 14–30. [Google Scholar] [CrossRef]
- Frioni, T.; Saracino, S.; Squeri, C.; Tombesi, S.; Palliotti, A.; Sabbatini, P.; Magnanini, E.; Poni, S. Understanding kaolin effects on grapevine leaf and whole-canopy physiology during water stress and re-watering. J. Plant Physiol. 2019, 242, 153020. [Google Scholar] [CrossRef]
- Cubasch, U.; Wuebbles, D.; Chen, D.; Facchini, M.C.; Frame, D.; Mahowald, N.; Winther, J.G. The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattenr, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Migley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 119–158. [Google Scholar] [CrossRef]
- Prodhomme, D.; Valls-Fonayet, J.; Hévin, C.; Franc, C.; Hilbert, G.; De Revel, G.; Richard, T.; Ollat, N.; Cookson, S.J. Metabolite profiling during graft union formation reveals the reprogramming of primary metabolism and the induction of stilbene synthesis at the graft interface in grapevine. BMC Plant. Biol. 2019, 19, 599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambacorta, G.; Antonacci, D.; La Gatta, M.; Faccia, M.; La Gatta, B.; Pati, S.; Coletta, A.; La Notte, E. Phenolic composition of Aglianico and Nero di Troia grapes and wines as affected by cover cropping and irrigation. Ital. J. Food Sci. 2011, 23, 381–394. [Google Scholar]
- Baiano, A.; De Gianni, A.; Previtali, M.A.; Del Nobile, M.A.; Novello, V.; de Palma, L. Effects of defoliation on quality attributes of Nero di Troia (Vitis vinifera L.) grape and wine. Food Res. Int. 2015, 75, 260–269. [Google Scholar] [CrossRef]
- Suriano, S.; Alba, V.; Di Gennaro, D.; Suriano, M.S.; Savino, M.; Tarricone, L. Genotype/rootstocks effect on the expression of anthocyanins and flavans in grapes and wines of Greco Nero n. (Vitis vinifera L.). Sci. Hortic. 2016, 209, 309–315. [Google Scholar] [CrossRef]
- Tarricone, L.; Alba, V.; Di Gennaro, D.; Amendolagine, A.M.; Gentilesco, G.; Masi, G. Grape and wine quality of Vitis vinifera ‘Nero di Troia’ in response to moderate deficit irrigation. Acta Hortic. 2017, 1150, 485–492. [Google Scholar] [CrossRef]
- Silvestroni, O.; Lanari, V.; Lattanzi, T.; Palliotti, A.; Vanderweide, J.; Sabbatini, P. Canopy managements strategies to control yield and grape composition of Montepulciano grapevines. Aust. J. Grape Wine Res. 2019, 25, 30–42. [Google Scholar] [CrossRef]
- Buesa, I.; Caccavello, G.; Basile, B.; Merli, M.C.; Poni, S.; Chirivella, C.; Intrigliolo, D.S. Delaying berry ripening of Bobal and Tempranillo grapevines by late leaf removal in a semi-arid and temperate-warm climate under different water regimes. Aust. J. Grape Wine Res. 2019, 25, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Bubola, M.; Lukić, I.; Radeka, S.; Sivilotti, P.; Grozi, K.; Vanzo, A.; Bavcar, D.; Lisjak, K. Enhancement of Istrian Malvasia wine aroma and hydroxycinnamate composition by hand and mechanical leaf removal. J. Sci. Food Agric. 2019, 99, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Petrie, P.R.; Trought, M.C.T.; Howell, G.S.; Buchan, G. The effect of leaf removal and canopy height on whole-vine gas exchange and fruit development of Vitis vinifera L. Sauvignon blanc. Funct. Plant Biol. 2003, 30, 711–717. [Google Scholar] [CrossRef] [PubMed]
- de Palma, L.; Tarricone, L.; Muci, G.; Limosani, P.; Savino, M.; Novello, V. Leaf removal, vine physiology and wine quality in cv Negramaro (Vitis vinifera L). In Proceedings of the 17th International Symposium GiESCO, Asti-Alba, Italy, 29 August–2 September 2011; pp. 231–234. [Google Scholar]
- Sivilotti, P.; Herrera, J.C.; Lisjak, K.; Bašaesnik, H.; Sabbatini, P.; Peterlunger, E.; Castellarin, S.D. Impact of leaf removal, applied before and after flowering, on anthocyanin, tannin, and methoxypyrazine concentrations in ‘Merlot’ (Vitis vinifera L.) grapes and wines. J. Agric. Food Chem. 2016, 64, 4487–4496. [Google Scholar] [CrossRef] [Green Version]
- Mosetti, D.; Herrera, J.C.; Sabbatini, P.; Green, A.; Alberti, G.; Peterlunger, E.; Lisjak, K.; Castellarin, S.D. Impact of leaf removal after berry set on fruit composition and bunch rot ‘Sauvignon blanc’. VITIS—J. Grapevine Res. 2016, 55, 57–64. [Google Scholar] [CrossRef]
- Wang, Y.; He, L.; Pan, Q.; Duan, C.; Wang, J. Effects of Basal Defoliation on Wine Aromas: A Meta-Analysis. Molecules 2018, 23, 779. [Google Scholar] [CrossRef] [Green Version]
- Diago, M.P.; Ayestaran, B.; Guadalupe, Z.; Poni, S.; Tardaguila, J. Impact of prebloom and fruit set basal leaf removal on the flavonol and anthocyanin composition of Tempranillo grapes. Am. J. Enol. Vitic. 2012, 63, 367–376. [Google Scholar] [CrossRef]
- Sternad Lemut, M.; Sivilotti, P.; Franceschi, P.; Wehrens, R.; Vrhovsek, U. Use of metabolic profiling to study grape skin polyphenol behavior as a result of canopy microclimate manipulation in a ‘Pinot noir’ vineyard. J. Agric. Food Chem. 2013, 61, 8976–8986. [Google Scholar] [CrossRef]
- Frioni, T.; Zhuang, S.; Palliotti, A.; Sivilotti, P.; Falchi, R.; Sabbatini, P. Leaf removal and cluster thinning efficiencies are highly modulated by environmental conditions in cool climate viticulture. Am. J. Enol. Vitic. 2017, 68, 325–335. [Google Scholar] [CrossRef] [Green Version]
- Šuklje, K.; Antalick, G.; Coetzee, Z.; Schmidtke, L.M.; Baša Cesnik, H.; Brandt, J.; du Toit, W.J.; Lisjak, K.; Deloire, A. Effect of leaf removal and ultraviolet radiation on the composition and sensory perception of Vitis vinifera L. cv. Sauvignon Blanc wine. Aust. J. Grape Wine Res. 2014, 20, 223–233. [Google Scholar] [CrossRef]
- Suriano, S.; Savino, M.; Basile, T.; Tarricone, L.; Di Gennaro, D. Management of malolactic fermentation and influence on chemical composition of Aglianico red wines. Ital. J. Food Sci. 2015, 3, 310–319. [Google Scholar]
- Iannini, C.; Mattii, G.B.; Rivelli, A.R.; Rotundo, A. Leaf removal and cluster thinning trials in Aglianico grapevine. Acta Hortic. 2007, 754, 241–248. [Google Scholar] [CrossRef]
- Caccavello, G.; Giaccone, M.; Scognamiglio, P.; Forlani, M.; Basile, B. Influence of intensity of post-veraison defoliation or shoot trimming on vine physiology, yield components, berry and wine composition in Aglianico grapevines. Aust. J. Grape Wine Res. 2016, 22, 1–14. [Google Scholar] [CrossRef]
- EEC 2676. Commission Regulation No 000/90 of 17 September 1990 Determining Community Methods for the Analysis of Wines. Off. J. Eur. Communities 1990, 81–83. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A01990R2676-20050813 (accessed on 22 October 2020).
- Di Stefano, R.; Cravero, M.C. Methods for grape phenolic compound study. Riv. Vitic. Enol. 2001, 2, 37–45. [Google Scholar]
- Trani, A.; Verrastro, V.; Punzi, R.; Faccia, M.; Gambacorta, G. Phenols, volatiles and sensory properties of Primitivo wines from the “Gioia Del Colle” PDO area. S. Afr. J. Enol. Vitic. 2016, 37, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Baiano, A.; Terracone, C.; Gambacorta, G.; La Notte, E. Phenolic content and antioxidant activity of Primitivo wine: Comparison among winemaking technologies. J. Food Sci. 2009, 74, C258–C267. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Miller, N. Measurement of the antioxidant status of dietary constituents, low density lipoprotein and plasma. Prostaglandins Leukot. Essent. Fat. Acids 1997, 57, 499–505. [Google Scholar] [CrossRef]
- Labanca, F.; Faraone, I.; Nolè, M.R.; Hornedo-Ortega, R.; Russo, D.; García-Parrilla, M.C.; Chiummiento, L.; Bonomo, M.G.; Milella, L. New Insights into the Exploitation of Vitis vinifera L. cv. Aglianico Leaf Extracts for Nutraceutical Purposes. Antioxidants 2020, 9, 708. [Google Scholar] [CrossRef]
- Revilla, E.; Ryan, L.M. Analysis of several phenolic compounds with potential antioxidant properties in grapes extracts and wines by high-performance liquid chromatography-photodiode array detection without sample preparation. J. Chrom. A 2000, 881, 461–469. [Google Scholar] [CrossRef]
- Frioni, T.; Acimovic, D.; Tombesi, S.; Sivilotti, P.; Palliotti, A.; Poni, S.; Sabbatini, P. Changes in within-shoot carbon partitioning in Pinot noir grapevines subjected to early basal leaf removal. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Poni, S.; Casalini, L.; Bernizzoni, F.; Civardi, S.; Intrieri, C. Effects of early defoliation on shoot photosynthesis, yield components, and grape composition. Am. J. Enol. Vitic. 2006, 57, 397–407. [Google Scholar]
- Ivanišević, D.; Kalajdžić, M.; Drenjančević, M.; Puškaš, V.; Korać, N. The impact of cluster thinning and leaf removal timing on the grape quality and concentration of monomeric anthocyanins in Cabernet-Sauvignon and Probus (Vitis vinifera L.) wines. OENO One 2020, 1, 63–74. [Google Scholar] [CrossRef]
- Casassa, L.F.; Harbertson, J.F. Extraction, evolution, and sensory impact of phenolic compounds during red wine maceration. Annu. Rev. Food Sci. Technol. 2014, 5, 83–109. [Google Scholar] [CrossRef] [PubMed]
- VanderWeide, J.; Tombesi, S.; Castellarin, S.D.; Sabbatini, P. Canopy architecture and fruit microclimate, not ripening-related phytohormones, control phenylpropanoid accumulation in response to early leaf removal in ‘Merlot’ (Vitis vinifera L.) grapevines. Plant Physiol. Biochem. 2020, 157, 291302. [Google Scholar] [CrossRef] [PubMed]
- Hunter, J.J.; Tarricone, L.; Volschenk, C.G.; Giacalone, C.; Melo, M.S.; Zorer, R. Grapevine physiological response to row orientation-induced spatial radiation and microclimate changes. OENO One 2020, 54, 411–433. [Google Scholar] [CrossRef]
- Chorti, E.; Theocharis, S.; Boulokostas, K.; Kallithraka, S.; Kotseridis, Y.; Koundouras, S. Row orientation and defoliation effects on grape composition of Vitis vinifera L. Agiorgitiko in Nemea (Greece). E3S Web Conf. 2018, 50, 01039. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Parameters | Average 1970–2000 | Annual | From Bud Break to Harvest (1 April to 31 October) | ||||
|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | 2016 | 2017 | 2018 | ||
| Mean annual T (°C) | 16.3 | 16.0 | 16.2 | 16.4 | 20.4 | 21.3 | 21.3 |
| Min mean T (°C) | −2.3 | −3.8 | −6.3 | −6.7 | 3.8 | 2.8 | 5.0 |
| Max mean T (°C) | 38.7 | 24.7 † | 22.8 † | 22.7 † | 40.4 | 44.2 | 39.9 |
| Rainfall (mm) | 563 | 669 | 349 | 633 | 455 | 128 | 306 |
| Accumulated GDD (°C) | 2330 | - | - | - | 2078 | 2311 | 2276 |
| Days daily max. T > 35 °C | 7 | - | - | - | 8 | 34 | 9 |
| ETr (mm) | 1063 | 1016 | 1187 | 1059 | 833 | 959 | 840 |
| Source of Variation | Cluster Number | Cluster Weight (g) | Yield (kg/vine) | Leaf Area per Vine (m2/vine) |
|---|---|---|---|---|
| Season (S) | ||||
| 2016 | 26.38 | 172.74 | 4.56 | 3.75 |
| 2017 | 25.86 | 177.21 | 4.30 | 3.50 |
| 2018 | 26.42 | 162.16 | 4.26 | 3.65 |
| Significance | ns | ns | ns | ns |
| Leaf removal (LR) | ||||
| Control | 24.25 | 172.86 | 4.08 | 4.75a |
| North | 26.39 | 154.98 | 4.09 | 3.12c |
| South | 28.50 | 160.51 | 4.52 | 3.76b |
| North-South | 25.61 | 166.56 | 4.89 | 2.90c |
| Significance | ns | ns | ns | *** |
| Interaction | ||||
| S*LR | ns | * | ns | ** |
| Source of Variation | Total Soluble Solids (Brix) | pH | Titratable Acidity (g/L) | Flavonoids (mg/kg) | Anthocyanins (mg/kg) | FRV (mg/kg) | Proantocyanidins (mg/kg) | FRV/P | TP (mg/kg) | Antioxidant Activity (mmol/kg) |
|---|---|---|---|---|---|---|---|---|---|---|
| Season (S) | ||||||||||
| 2016 | 23.37b † | 3.11b | 7.08a | 2925b | 1707b | 973b | 1809b | 0.54a | 2354b | 9.93b |
| 2017 | 25.43a | 3.14b | 7.45a | 4716a | 3098a | 1271a | 2282a | 0.55a | 4141a | 19.70a |
| 2018 | 23.58b | 3.30a | 5.27b | 2530c | 1600b | 691c | 1834b | 0.38b | 2009c | 10.76b |
| Significance | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Leaf removal | ||||||||||
| Control | 23.79 | 3.18 | 6.50 | 3517 | 2126 | 897b | 1826b | 0.49a | 2693b | 13.45ab |
| North | 24.16 | 3.14 | 6.95 | 3234 | 2115 | 1005ab | 1953ab | 0.50a | 2632b | 11.90b |
| South | 24.59 | 3.22 | 6.31 | 3474 | 2139 | 927b | 2047a | 0.45b | 3122a | 15.24a |
| North-South | 23.95 | 3.18 | 6.63 | 3335 | 2160 | 1084a | 2074a | 0.51a | 2891ab | 13.26ab |
| Significance | ns | ns | ns | ns | ns | *** | *** | * | *** | *** |
| Interaction | ||||||||||
| S*LR | ns | ns | ns | ** | *** | *** | *** | *** | *** | *** |
| Source of Variation | Dp | Cy | Pt | Pn | Mv | Dp-Ac | Pt-Ac | Pn-Ac | Mv-Ac | c-Mv-Cm | Mv-Cf | Pt-Cm | Pn-Cm | t-Mv-Cm | Total Anthocyanins |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Season (S) | |||||||||||||||
| 2016 | 76.1b | 5.0b | 84.1b | 39.9b | 617.7b | 3.1b | 13.5a | 19.5b | 31.3b | 1.1b | 13.7b | 6.9c | 12.6b | 178.7b | 1098.8b |
| 2017 | 177.5a | 11.8a | 190.6a | 72.8a | 1208.6a | 7.8a | 13.6a | 27.0a | 58.3a | 1.6a | 29.2a | 11.1a | 18.6a | 281.6a | 2019.0a |
| 2018 | 55.5b | 3.3b | 62.0b | 28.8b | 619.6b | 2.7b | 4.5b | 13.3b | 48.9a | 1.5a | 15.0b | 9.1b | 11.1b | 258.0a | 1133.4b |
| Significance | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Leaf removal | |||||||||||||||
| Control | 64.4b | 4.8b | 83.6b | 39.7b | 784.9b | 4.3b | 9.3c | 15.1b | 42.4b | 1.6a | 16.4c | 8.9 | 12.4b | 234.1b | 1264.0c |
| North | 112.9a | 7.9a | 121.2a | 53.6a | 770.2b | 4.4b | 10.4b | 22.6a | 43.5b | 1.4ab | 18.0b | 8.9 | 16.9a | 217.8b | 1373.7b |
| South | 110.2a | 6.2ab | 120.3a | 43.5b | 863.8a | 4.4b | 12.0a | 22.2a | 51.3a | 1.2b | 22.1a | 9.1 | 13.5b | 273.8a | 1523.0a |
| North-South | 124.6a | 7.8a | 123.9a | 51.7a | 842.4a | 5.0a | 10.4b | 19.8a | 47.4a | 1.4ab | 20.7a | 9.3 | 13.5b | 232.1b | 1507.5a |
| Significance | *** | *** | *** | ** | * | * | *** | *** | ** | *** | ** | ns | *** | ** | *** |
| Interaction | |||||||||||||||
| S*LR | *** | *** | *** | *** | *** | *** | ** | *** | ** | *** | ** | ns | *** | ** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarricone, L.; Faccia, M.; Masi, G.; Gambacorta, G. The Impact of Early Basal Leaf Removal at Different Sides of the Canopy on Aglianico Grape Quality. Agriculture 2020, 10, 630. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10120630
Tarricone L, Faccia M, Masi G, Gambacorta G. The Impact of Early Basal Leaf Removal at Different Sides of the Canopy on Aglianico Grape Quality. Agriculture. 2020; 10(12):630. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10120630
Chicago/Turabian StyleTarricone, Luigi, Michele Faccia, Gianvito Masi, and Giuseppe Gambacorta. 2020. "The Impact of Early Basal Leaf Removal at Different Sides of the Canopy on Aglianico Grape Quality" Agriculture 10, no. 12: 630. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10120630