The Screening of Resistance against Meloidogyne graminicola in Oats


1
Department of Plant Pathology and Microbiology, National Taiwan University, Taipei 10617, Taiwan
2
Department of Agronomy, National Taiwan University, Taipei 10617, Taiwan
*
Author to whom correspondence should be addressed.
Agriculture 2020, 10(8), 352; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10080352
Submission received: 25 July 2020
/
Revised: 11 August 2020
/
Accepted: 11 August 2020
/
Published: 13 August 2020
(This article belongs to the Section Crop Protection, Diseases, Pests and Weeds)
Abstract
:Including pest resistance elements against the major local concern is naturally important in the breeding process. Oat (Avena sativa L.) has been recently reintroduced into Taiwan as a winter alternative forage crop, and its agronomic performance has been evaluated at different locations in the country. This study examined the resistance to root-knot nematode, Meloidogyne graminicola, in four oat (Avena sativa L.) breeding lines of mass planting potential for winter in Taiwan. The host attraction level to the nematode, and the penetration and reproduction ability of nematode towards host roots were evaluated by chemotaxis assay, root staining assay, root galling, and nematode extraction. Based on the gall index (GI) and multiplication factor (R), the resistance of each oat line was evaluated. At 24 h postinoculation, second-stage juvenile (J2) nematodes appeared most attracted by oat breeding lines UFRGS136104-3 and UFRGS136119-2. The number of J2s successfully penetrated into the two breeding lines were also high. However, at 40 days postinoculation, observation of the oats in the newly developed culture bag nematode-inoculation system revealed that the amount of root galls and 2nd generation nematodes were significantly higher in line LA08085BS-T2 than in other lines. In sum, oat breeding line UFRGS136104-3 was highly resistant to M. graminicola by inhibiting the gall formation and nematode reproduction, while UFRGS136106-3 and UFRGS136119-2 showed relatively weak resistance and oat line LA08085BS-T2 would be a moderately susceptible host to M. graminicola, with high numbers of root gall formation. The outcome of this study provides ground information for nematode-resistant oat cultivar breeding.
1. Introduction
The rice root-knot nematode, Meloidogyne graminicola, is a common plant-parasitic nematode to the Poaceae crops globally. The main hosts of M. graminicola in the family of Poaceae includes rice, wheat, barley, oat, and sorghum [1]. Research has shown that this nematode can result in up to 73–80% yield loss on rice whether in upland or intermittently flooded condition when the nematode population is high [2,3]. M. graminicola is widely distributed in all agricultural regions in Taiwan. It completes one life cycle within 19–21 days at 28–30 °C and causes an average of 80% disease rate in rice paddy fields [4]. The aboveground symptoms of M. graminicola infections often appear as patch patterns in a field, with the infected plants showing growth reduction, less vigor, yellowing and curling of leaves, wilting, and poor yield [5]. Underground, the 2nd juvenile stage of the nematode invades rice roots tips and the infected root tips would swell and form horseshoe-like hooks, the characteristic disease symptom of this nematode, as the female develops and lays eggs within the roots. Management of the M. graminicola in Taiwan is difficult since no nematicide is currently approved on the Poaceae crops for M. graminicola because of concerns for human and environment health [6]. Long-term crop rotation or bare fallow without any weed in field were shown to be effective in nematode control with the use of proper crop sequence [7] but is very impractical to most growers. Although recent progress had been made in biological control research [8], planting M. graminicola-resistant cultivars still remains the most promising option for future effective and economic control.
Oat (Avena sativa L.) is an annual grass grown worldwide. It is mainly cultivated for grain use in temperate regions between 45–65° of the northern hemisphere and 20–46° in the southern hemisphere [8]. Meanwhile, in the tropics and subtropics regions, oats are grown as forage for local use in some mountainous area or during cooler seasons [9]. As forage, oat is a high biomass crop with good nutrition composition profile that contains high crude protein and water-soluble carbohydrate and could be used as fresh feed, hay, or plant material for silage. Recent years, under the government’s support and promotion, oat has been reintroduced to Taiwan as a winter forage crop [10,11].
Natural nematode resistance has been reported in oats to some root-knot nematodes [12,13] and to cereal cyst nematode Heterodera avenae [14]. Three oat cultivars, Sniper, Tachiibuki, and Haeibuki, have been tested against three root-knot nematode species, M. incognita, M. arenaria, and M. hapla. Oat cultivars Sniper and Tachiibuki showed suppressive ability in the reproduction and development of these 3 Meloidogyne species in contrast to cultivar Haeibuki [12,13]. However, no breeding or experiments in oats for M. graminicola resistance have been reported. Given the fact that oat planting has gradually gained popularity in Taiwan, along with the raising economic risk caused by nematode infection, the aim of this study was to examine the disease resistance to M. graminicola among the putative oat lines that are currently used for winter forage crop breeding in Taiwan.
2. Materials and Methods
2.1. Nematode Inoculum
The nematode M. graminicola was first collected from a field with rice cropping history in Taiwan and identified morphologically through microscopic anatomy observation and body measurements with the deMan formula [15] and confirmed molecularly with ribosome gene sequences [8]. A single egg population was maintained on rice (cultivar Taoyuan No. 3) at 28 °C in a growth chamber. Prior to each experiment, eggs were extracted by macerating oats roots for 2 min in 1% NaClO [16] and placed in a Petri dish that contains water for hatching. The 2nd stage juveniles (J2) were collected on the same day of experiment.
2.2. Plant Materials
Four oat breeding lines (LA08085BS-T2, UFRGS136104-3, UFRGS136106-3, and UFRGS136119-2) from Quaker Oat International Nursery (QION) 2015 and 2016 were tested in this study. LA08085BS-T2 was a breeding line from the oat breeding program of the University of Florida, derived from a cross of a Brazilian line and a Florida line (UFRGS 046048-1 F6/FL0206FSB-34-S1-B-S1). The UFRGS series were from the breeding program of Universidade Federal do Rio Grande do Sul, Brazil, where UFRGS136104-3 and UFRGS136106-3 had the same pedigree but were from two independent crosses and UFRGS136119-2 was derived from a cross between UFRGS 988012-1 and UFRGS 995088-3 (Supplementary Table S1). We have selected these four oat lines for their good adaptation to subtropical climates. Thirty-five seedlings of each oat line were used in this study.
2.3. Attraction and Penetration Study
To quickly examine the oat lines for potential resistance to M. graminicola, the chemotaxis assay was conducted with oat seedlings. The seed of selected oat lines were sterilized with 1% sodium hypochlorite (NaClO) solution for 3 min and then thoroughly washed with distilled water. After soaking in distilled water for 2 h, the seeds were set to germinate in the dark for 24 h at 27 °C. The seedlings were then moved to 24 °C with 12–12 light/dark condition for 2 days before use. For evaluating the attraction of oats to M. graminicola, 200 J2 was added together with 18 mL of 23% Pluronic gel (Pluronic F-127, Sigma-Aldrich, St. Louis, MO, USA) in a petri dish (9 cm diameter) by shaking gently for uniform distribution. Then, one oat seedling was put in the gel in each petri dish. Throughout the process, the petri dishes were placed on ice to retain the liquid state of gel. The petri dishes were then transferred to room temperature for the gel to set. The number of nematodes within 1 mm diameter of the oat root tip were recorded at 8 and 24 h after inoculation. Five replications were set up for each tested oat line.
To determine the penetration rate of an oat breeding line, the total number of J2 which had completed root penetration at 24 h after inoculation were counted after staining with acid fuchsin, as described by Bybd et al. [17]. In brief, the roots were washed and placed in 1.5% NaClO solution for 4 min with occasional agitation. After removing residual NaClO, the segments were stained by boiling for 30 s in staining solution. The staining solution contains 1 mL stain (3.5 g acid fuchsin, 250 mL acetic acid, and 750 mL distilled water) with 30 mL water. The roots were rinsed in running water and then placed in acidified glycerin, heated to boiling, and then cooled to room temperature. The stained J2 inside the roots were counted under a stereoscopic binocular microscope. Five replications were set up for each treatment.
2.4. Inoculation Study
In order to observe and compare the disease development difference among oat breeding lines, a nematode-inoculation culture bag system was used. Three fresh germinated seeds of each breeding line were placed in the folding line of one culture bag and incubated in the growth chamber at 24 °C for 3 days. 200 J2 was inoculated to the 3-day-old seedlings as described by Lasserre et al. [18]. The disease severity and nematode reproduction ability were used to evaluate the resistance of oat genotypes. Plants with no nematode inoculation were used as controls. Five replicates were set up for each treatment.
The numbers of root galls represent the disease severity in this trail. The gall index (GI) was rated at 40 days postinoculation (dpi) with a five-scales system: 1 = no galling; 2 = 1–10 galls; 3 = 11–30 galls; 4 = 31–100 galls; and 5 = >100 galls/root system [18]. In addition, the culture bags were observed every day to record the occurrence of the first gall on each root system. All root was freshly weighted at the end of the experiment.
The number of eggs and the juveniles of the 2nd generation were used to evaluate the nematode reproduction ability. At the end of the trial, the total amount of J2 and eggs were collected and counted. With the initial population density (Pi) as 200, the J2 amount at the beginning of the inoculation, the J2 collected from the water in culture bags were used to calculate the final population density (Pf). Then, the multiplication factors (R) were calculated (R = Pf/Pi). Eggs were harvested by macerating oat roots in 1% NaClO for 2 mins, and the eggs were transferred into sterile water for counting as previously described [19].
2.5. Evaluation of Resistance and Susceptibility of Oat Lines
2.6. Data Analysis
For attraction and penetration study, the numbers of J2 were analyzed through the analysis of variance (ANOVA) and followed with the Duncan’s Multiple Range Test (DMRT) to measure the differences between means. For inoculation study, all data of root gall formation, nematode reproduction, and plant weight were analyzed by ANOVA and means were separated by DMRT. All the statistical analyses were performed using SAS 9.4 software (SAS Institute Inc., Cary, NC, USA). Significant differences were determined by p < 0.05.
3. Results
3.1. Attraction and Penetration Study
Test oat lines showed variable attraction to J2. After 8 h, there were the most amounts of J2 observed around the root tips of UFRGS136104-3, which were significantly (p < 0.05) greater than the number of J2 around the root tips of lines LA08085BS-T2 and UFRGS136106-3, while no significant difference was found between UFRGS136119-2 and the other three genotypes (Figure 1A). After 24 h, significantly fewer J2 were found around UFRGS136119-2, followed by UFRGS136104-3, LA08085BS-T2, and UFRGS136106-3 (Figure 1B).
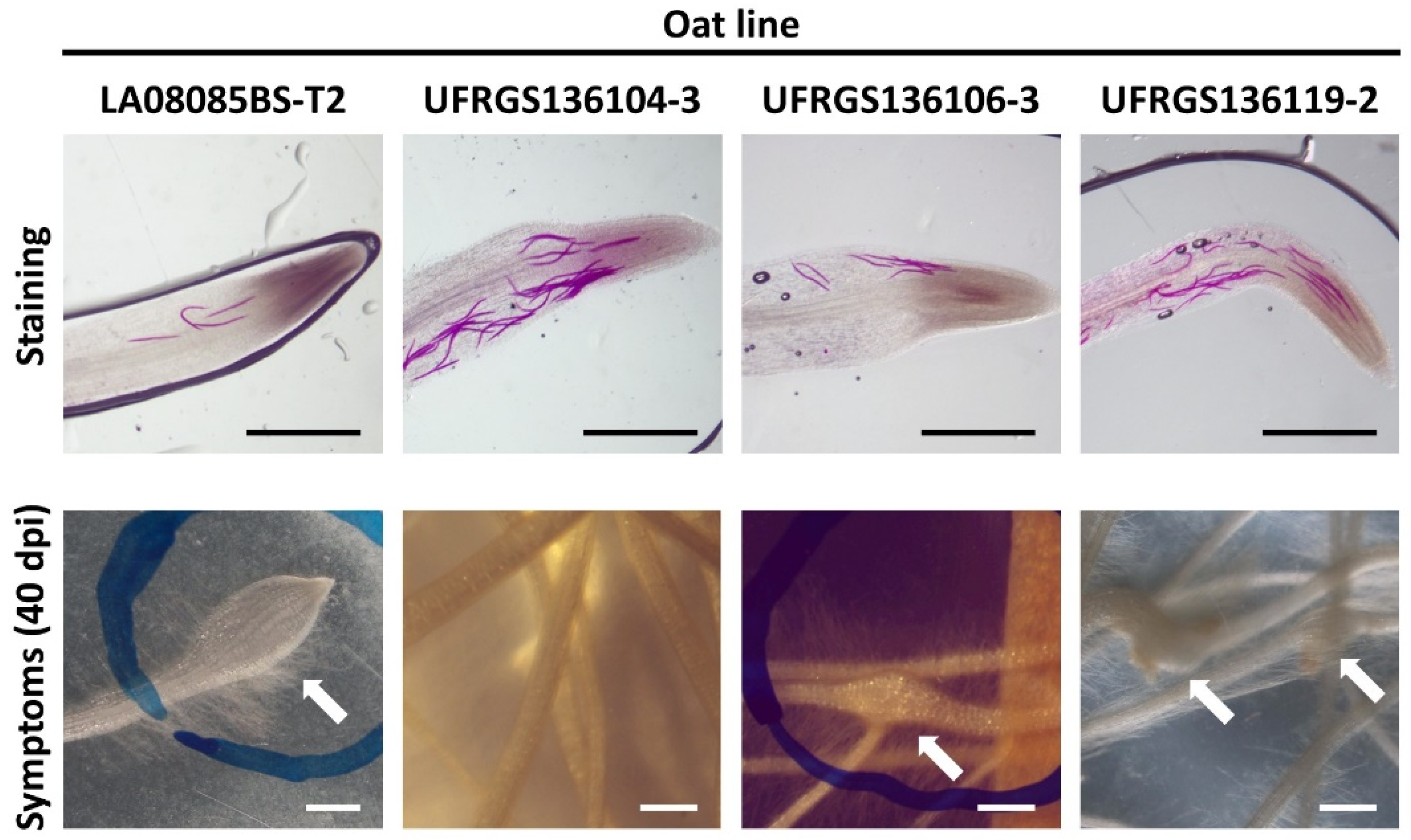
After acid fuchsin staining, J2 could be clearly found inside the root tips of oats at 24 h after inoculation (Figure 2). There were significant differences (p < 0.05) in J2 penetration observed after 24 h (Figure 1C). Four to eight times the number of J2 were found in the root tips of lines UFRGS136104-3 and UFRGS136119-2 compared to lines LA08085BS-T2 and UFRGS136106-3 (Figure 1C). Combining the number of J2 around and inside the root tips of each oat line after 24 h, a significantly (p < 0.05) greater number of J2 were attracted and penetrated to the root tips of lines UFRGS136104-3 and UFRGS136119-2 than those of lines LA08085BS-T2 and UFRGS136106-3 (Figure 1D).
3.2. Inoculation Study
The first gall formation of lines LA08085BS-T2, UFRGS136106-3, and UFRGS136119-2 could be observed at 10 dpi, 3 dpi, and 6 dpi, respectively. At 40 dpi, the typical symptoms caused by M. graminicola such as swellings and horseshoe-like hooks were observed on several root tips of tested oat lines, except UFRGS136104-3 (Figure 2). No gall was observed in UFRGS136104-3 during the trial time (Table 2). The total number of root galls, incompletely developed root galls, and completely developed root galls of line LA08085BS-T2 were all significantly higher (p < 0.05) when compared with UFRGS136106-3 and UFRGS136119-2 (Table 2). At 40 dpi, M. graminicola produced a significantly (p < 0.05) high number of eggs in LA08085BS-T2 (Table 2). Also, the number of J2 harvested in LA08085BS-T2 were more than UFRGS136104-3, UFRGS136106-3, and UFRGS136119-2 (Table 2). The fresh root weight of oats was increased in lines LA08085BS-T2, UFRGS136106-3, and UFRGS136119-2 at 40 dpi, but there were no significant differences in fresh root and leaf weight of inoculated oat lines compared with control plants (Table 3).
3.3. Resistance/Susceptibility of Oats Evaluation
The resistance/susceptibility of tested oat lines are summarized in Table 4. LA08085BS-T2 expressed as a moderately susceptible host to M. graminicola, which might not be easily invaded (Figure 1) initially but the nematodes population could successfully establish feeding sites and form many root galls in the later stage. In contrast, UFRGS136104-3 was highly resistant to M. graminicola; Even with some J2 penetration in the roots (Figure 1), the nematodes did not survive, no feeding sites could be developed within, and therefore no gall was observed. UFRGS136106-3 showed resistance to M. graminicola, and the nematode population within was the smallest among the tested oat lines. Finally, UFRGS136119-2 exhibited resistance by possessed fewer root gall formation and led to ineffective nematode reproduction.
4. Discussion
Meloidogyne graminicola, rice root-knot nematode, is one of the limiting factors for the production of the Poaceae crops, yet little is known about the pathogenicity of M. graminicola to oats. In the present study, the resistance level of oat lines from forage crop breeding lines in Taiwan were revealed using petri dish chemotaxis assay, root-tip penetration assay, and the newly developed culture bag nematode-inoculation system. The experiment results not only filled the important gap knowledge for M. graminicola host range and disease development on root but also revealed the limitation of using the results of petri dish chemotaxis and root-tip penetration assays as resistance/susceptibility indicators. In the case of oat line UFRGS136104-3, many J2s were attracted to the root tip and successfully entered the root, but a strong resistance occurred at a later stage in plants that led to no gall formation. On the other hand, on oat line UFRGS136106-3, even though the first gall was observed very early in the trail, only gentle damage was caused by the nematode at the end of the experiment.
The plant-parasitic nematodes can recognize specific chemical substances from the hosts by their sensory organs (e.g., amphids and phasmids), and it allows the nematodes to seek for food [22]. We found that the attraction to M. graminicola J2 of tested oat lines UFRGS136104-3 and UFRGS136119-2 were much higher than lines LA08085BS-T2 and UFRGS136106-3. However, the level of host attraction to J2 did not associate with the plant resistance level. Cabasan et al. [23] have observed the equal attraction to M. graminicola J2 between resistant and susceptible rice genotypes.
Host penetration is a key step of root-knot nematodes to complete their life cycles, and the defense responses that they may encounter from host plants could be pre-penetration, post-penetration, or both [24]. Pre-penetration defenses include physical root barriers and plant defensive biochemicals to inhibit Meloidogyne spp. penetration [25,26]. Two types of post-penetration defenses of M. graminicola-resistant rice genotypes were previously described [27]. First, an early hypersensitive-response (HR)-like reaction caused cell necrosis to prevent nematode feeding. Second, a late response resulted in poor developments of giant cells. Our results indicated that there are different defense response expressions in oat lines UFRGS136104-3, UFRGS136106-3, and UFRGS136119-2. The reproduction results suggest that oat lines UFRGS136104-3 and UFRGS136106-3 expressed pre-penetration or early stage post-penetration resistance, while line UFRGS136119-2 exhibited late post-penetration resistance (Figure 3). Further gene expression studies on nematode associated molecular pattern (NAMP)-induced basal defense associated host genes or markers of effector triggered host defense pathways would support the observation.
The resistance mechanism that lies in the M. graminicola oat line may not resemble the ones previously reported from other Poaceae plants. To date, 19 resistant genes to Heterodera avenae have been identified in wheat, barley, and oat [28,29,30,31,32]. Three flavone-C-glycoside compounds in methanolic root and shoot extracts of oats were found to be induced by Pratylenchus neglectus and H. avenae invasion and by methyl jasmonate [33]. Treatment with methyl jasmonate reduced invasion of both nematodes and increased plant mass, compensating for damage caused by the nematodes, and is attributed to the active flavone-C-glycoside [33]. In M. graminicola-resistant rice cultivars, the amount of nematode J2 with successful penetration was significantly lower, the nematode development was slower, their reproduction in the host was slower, significantly fewer galls were observed, and the size of mature females were significantly smaller [23]. However, in the inoculation experiment of this study, M. graminicola successfully penetrated the root of oat line UFRGS136104-3 with relatively high J2 amounts at 24 hours postinoculation but was not able to induce any gall or to reproduce. Based on the data obtained in the present research, there might be certain genetic properties for the resistances to M. graminicola existing in oat lines UFRGS136104-3, UFRGS136106-3, and UFRGS136119-2. Previous studies showed that the resistance genes in peppers (N, Me1, Me2, Me3, Me4, Me5, Me6, Mech1, and Mech2) and in tomato (Mi) regulated the plants abilities to defend against the penetration of Meloidogyne spp. and inhibited nematode reproduction [34,35,36,37]. Therefore, further plant physiological experiments along with genomic studies would be needed to reveal the M. graminicola-resistant mechanism in oats.
The management of M. graminicola currently relies heavily on nematicides application. For long-term sustainable farming, incorporating the nematode-resistance quality into oat cultivar selection would be ideal to prevent disease occurrence in fields. This study demonstrated and provided essential information on the natural M. graminicola resistance for future crop breeding.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2077-0472/10/8/352/s1, Table S1: The pedigree of oat breeding lines used in this study.
Author Contributions
Conceptualization, J.-I.Y. and Y.-F.H.; methodology, J.-I.Y.; software, S.-M.Y.; validation, J.-I.Y. and Y.-F.H.; formal analysis, S.-M.Y.; resources, Y.-F.H.; data curation, S.-M.Y.; writing—original draft preparation, S.-M.Y.; writing—review and editing, J.-I.Y. and Y.-F.H.; visualization, S.-M.Y.; supervision, J.-I.Y.; project administration, J.-I.Y.; funding acquisition, Y.-F.H. and J.-I.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science and Technology of Taiwan (grant number: 106-2311-B-002-026-MY3) and the Council of Agriculture, Executive Yuan of Taiwan (grant number: 105AS–2.3.4–AD–U1 and 106AS–2.3.4–AD–U1).
Acknowledgments
We are grateful to Stephen A. Harrison and Allison Lunos Harding from the Louisiana State University AgCenter for coordinating the Quaker International Oat Nursery germplasm exchange and to the many oat breeders who have contributed their materials to the nursery. The authors also thank Zi-Ying Chuang for her assistance on the experiments.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Vaish, S.; Pandey, S. Root-knot disease caused by Meloidogyne graminicola: A limiting factor for growth and yield of barley (Hordeum vulgare L.). Curr. Nematol. 2012, 23, 7–12. [Google Scholar]
- Plowright, R.A.; Bridge, J. Effect of Meloidogyne graminicola (Nematoda) on the establishment, growth and yield of rice cv. IR36. Nematologica 1990, 36, 81–89. [Google Scholar] [CrossRef]
- Soriano, I.R.S.; Prot, J.-C.; Matias, D.M. Expression of tolerance for Meloidogyne graminicola in rice cultivars as affected by soil type and flooding. J. Nematol. 2000, 32, 309–317. [Google Scholar]
- Duan, H.W. The Survey, Identification and Etiology Studies of Rice Root-Knot Nematode, Meloidogyne graminicola, in Taiwan. Master’s Thesis, National Chung Hsing University, Taichung, Taiwan, 2015. [Google Scholar]
- Mantelin, S.; Bellafiore, S.; Kyndt, T. Meloidogyne graminicola: A major threat to rice agriculture. Mol. Plant. Pathol. 2017, 18, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bureau of Animal and Plant Health Inspection and Quarantine (BAPHIQ). Available online: pesticide.baphiq.gov.tw (accessed on 5 July 2020).
- Rao, Y.S.; Prasad, J.S.; Panwar, M.S. Nematode problems in rice: Crop losses, symptomatology and management. In Plant Parasitic Nematodes of India: Problems and Progress; Swarup, G., Dasgupta, D.R., Eds.; Indian Agricultural Research Institute: New Delhi, India, 1986; pp. 279–299. [Google Scholar]
- Yang, J.I.; Stadler, M.; Chuang, W.Y.; Wu, S.; Ariyawansa, H.A. In vitro inferred interactions of selected entomopathogenic fungi from Taiwan and eggs of Meloidogyne graminicola. Mycol. Prog. 2020, 19, 97–109. [Google Scholar] [CrossRef]
- Suttie, J.M.; Reynolds, S.G. Fodder Oats: A World Overview; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004. [Google Scholar]
- Huang, C.W.; Liang, W.H.; Esvelt Klos, K.; Chen, C.S.; Huang, Y.F. Evaluation of agronomic performance and exploratory genome-wide association study of a diverse oat panel for forage use in Taiwan. Grassl. Sci. 2020. [Google Scholar] [CrossRef]
- Wang, S.H.; Shy, Y.M.; Chen, Y.H.; Chang, C.T.; Lee, K.H.; Jea, Y.S. Effect of oat silage substitute on lactation performance of Holstein lactating cows. J. Taiwan Livest. Res. 2018, 51, 109–115. [Google Scholar]
- Tateishi, Y.; Iwahori, H.; Uesugi, K.; Katsura, M. Invasion, development, and reproduction of 3 Meloidogyne species on oat cultivar Tachiibuki, a nematode-suppressive fall crop. Jpn. J. Nematol. 2012, 41, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Tateishi, Y.; Uesugi, K.; Iwahori, H. Host suitability of a common oat cultivar Sniper for five plant-parasitic nematode species. Jpn. J. Nematol. 2016, 46, 21–24. [Google Scholar] [CrossRef] [Green Version]
- Andersen, S.; Andersen, K. Suggestions for determination and terminology of pathotypes and genes for resistance in cyst-forming nematodes, especially Heterodera avenae 1. EPPO Bull. 1982, 12, 379–386. [Google Scholar] [CrossRef]
- Mai, W.; Mullin, P.G. Plant.-Parasitic Nematodes: A Pictorial Key to Genera, 5th ed.; Cornell University Press: Ithaca, NY, USA, 1996. [Google Scholar]
- Padgham, J.; Sikora, R. Biological control potential and modes of action of Bacillus megaterium against Meloidogyne graminicola on rice. Crop. Prot. 2007, 26, 971–977. [Google Scholar] [CrossRef]
- Bybd, D.W., Jr.; Kirkpatrick, T.; Barker, K.R. An improved technique for clearing and staining tissues for detection of nematodes. J. Nematol. 1983, 15, 142–143. [Google Scholar] [PubMed]
- Lasserre, F.; Rivoal, R.; Cook, R. Interactions between Heterodera avenae and Pratylenchus neglectus on wheat. J. Nematol. 1994, 26, 336. [Google Scholar] [PubMed]
- Dongre, M.; Simon, S. Infestation and host studies of Meloidogyne graminicola in rice: Nurseries of Allahabad. Int. J. Bot. Res. 2013, 3, 27–34. [Google Scholar]
- Taylor, A.L.; Sasser, J.N. Biology, Identification and Control. of Root-Knot Nematodes (Meloidogyne Species); Department of Plant Pathology, North Carolina State University Graphics and the USAID: Raleigh, NC, USA, 1978; p. 111. [Google Scholar]
- Sasser, J.C.; Carter, C.C.; Hartman, K.M. Standardization of Host Suitability Studies and Reporting of Resistance to Root-Knot Nematode; United States Agency for International Development (USAID): Raleigh, NC, USA, 1985; p. 7.
- Reynolds, A.M.; Dutta, T.K.; Curtis, R.H.C.; Powers, S.J.; Gaur, H.S.; Kerry, B.R. Chemotaxis can take plant-parasitic nematodes to the source of a chemoattractant via the shortest possible routes. J. R. Soc. Interface 2011, 8, 568–577. [Google Scholar] [CrossRef] [Green Version]
- Cabasan, M.T.N.; Kumar, A.; De Waele, D. Comparison of migration, penetration, development and reproduction of Meloidogyne graminicola on susceptible and resistant rice genotypes. Nematology 2012, 14, 405–415. [Google Scholar] [CrossRef]
- Pedrosa, E.M.R.; Hussey, R.S.; Boerma, H.R. Penetration and post-infectional development and reproduction of Meloidogyne arenaria races 1 and 2 on susceptible and resistant soybean genotypes. J. Nematol. 1996, 28, 343–351. [Google Scholar]
- Andwar, S.A.; McKenry, M.V. Penetration, development and reproduction of Meloidogyne arenaria on two new resistant Vitis spp. Nematropica 2000, 30, 9–17. [Google Scholar]
- Pegard, A.; Brizzard, G.; Fazari, A.; Soucaze, O.; Abad, P.; Djian-Caporalino, C. Histological characterization of resistance to different root-knot nematode species related to phenolics accumulation in Capsicum annuum. Phytopathology 2005, 95, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Cabasan, M.T.N.; Kumar, A.; Bellafiore, S.; De Waele, D. Histopathology of the rice root-knot nematode, Meloidogyne graminicola, on Oryza sativa and O. glaberrima. Nematology 2014, 16, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Andersen, S. Resistens mod Havreål Heterodera avenae; I Kommission hos Dansk Videnskabs Forlag: Copenhagen, Denmark, 1961. [Google Scholar]
- Clamot, G.; Rivoal, R. Genetic resistance to cereal cyst nematode Heterodera avenae WOLL. in wild oat Avena sterilis I. 376. Euphytica 1984, 33, 27–32. [Google Scholar] [CrossRef]
- Ireholm, A. Characterization of pathotypes of cereal cyst nematodes, Heterodera spp., in Sweden. Nematologica 1994, 40, 399–411. [Google Scholar] [CrossRef]
- Barr, A.R.; Chalmers, K.J.; Karakousis, A.; Kretschmer, J.M.; Manning, S.; Lance, R.C.M.; Lewis, J.; Jeffries, S.P.; Langridge, P. RFLP mapping of a new cereal cyst nematode resistance locus in barley. Plant. Breed. 1998, 117, 185–187. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Yamazaki, Y.; Dubcovsky, J.; Rogers, J.; Morris, C.; Somers, D.J.; Appels, R.; Devos, K.M. Catalogue of gene symbols for wheat. National BioResource Project, Komugi-Wheat Genetic Resources Database. Available online: https://shigen.nig.ac.jp/wheat/komugi/genes/download.jsp (accessed on 11 November 2019).
- Soriano, I.R.; Asenstorfer, R.E.; Schmidt, O.; Riley, I.T. Inducible flavone in oats (Avena sativa) is a novel defense against plant-parasitic nematodes. Phytopathology 2004, 94, 1207–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendy, H.; Dalmasso, A.; Cardin, M.C. Differences in resistant Capsicum annuum attacked by different Meloidogyne species. Nematologica 1985, 31, 72–78. [Google Scholar] [CrossRef]
- Kaloshian, I.; Yaghoobi, J.; Liharska, T.; Hontelez, J.; Hanson, D.; Hogan, P.; Jesse, T.; Wijbrandi, J.; Simons, G.; Vos, P.; et al. Genetic and physical localization of the root-knot nematode resistance locus Mi in tomato. Mol. Genet. Genom. 1998, 3, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Djian-Caporalino, C.; Fazari, A.; Arguel, M.J.; Vernie, T.; VandeCasteele, C.; Faure, I.; Brunoud, G.; Pijarowski, L.; Palloix, A. Root-knot nematode (Meloidogyne spp.) Me resistance genes in pepper (Capsicum annuum L.) are clustered on the P9 chromosome. Theor. Appl. Genet. 2007, 114, 473–486. [Google Scholar] [CrossRef]
- Hajihassani, A.; Rutter, W.B.; Luo, X. Resistant Pepper Carrying N, Me1, and Me3 have Different Effects on Penetration and Reproduction of Four Major Meloidogyne species. J. Nematol. 2019, 51, 1–9. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Numbers of M. graminicola J2 in a 1 mm diameter area around the root tips and in the roots of oat lines at 8 h and 24 h after inoculation: (A) Number of J2 around the oat root tips at 8 h after inoculation, (B) number of J2 around the oat root tips at 24 h after inoculation, (C) number of J2 in the oat roots at 24 h after inoculation, and (D) total number of J2 around the oat root tips and inside the oat roots after 24 h. Data points are mean ± standard error (SE) of 5 replicates. Means followed by the same letter are not significantly different (p ≥ 0.05).
Figure 1.
Numbers of M. graminicola J2 in a 1 mm diameter area around the root tips and in the roots of oat lines at 8 h and 24 h after inoculation: (A) Number of J2 around the oat root tips at 8 h after inoculation, (B) number of J2 around the oat root tips at 24 h after inoculation, (C) number of J2 in the oat roots at 24 h after inoculation, and (D) total number of J2 around the oat root tips and inside the oat roots after 24 h. Data points are mean ± standard error (SE) of 5 replicates. Means followed by the same letter are not significantly different (p ≥ 0.05).

Figure 2.
Penetration of M. graminicola observed by acid fuchsin staining at 24 h after inoculation and root symptoms at 40 days postinoculation (dpi): The arrow sticks indicate the swollen and horseshoe-like hooked root tips. The scale bars represent 1 mm.
Figure 2.
Penetration of M. graminicola observed by acid fuchsin staining at 24 h after inoculation and root symptoms at 40 days postinoculation (dpi): The arrow sticks indicate the swollen and horseshoe-like hooked root tips. The scale bars represent 1 mm.

Figure 3.
Overview of different defenses of oats against root-knot nematodes observed in this study. H = high; L = low; N = (almost) not observed; EPoP = early post-penetration; PrP = pre-penetration; LPoP = late post-penetration.
Figure 3.
Overview of different defenses of oats against root-knot nematodes observed in this study. H = high; L = low; N = (almost) not observed; EPoP = early post-penetration; PrP = pre-penetration; LPoP = late post-penetration.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Resistance rating scale for Meloidogyne graminicola based on root gall index (GI) and multiplication factor (R).
Table 1.
Resistance rating scale for Meloidogyne graminicola based on root gall index (GI) and multiplication factor (R).
| GI | R | |
|---|---|---|
| 1 | <0.1 | Highly resistant |
| 2 | ≤1 | Resistant |
| 3 | >1 | Moderately susceptible |
| 4 | >1 | Susceptible |
| 5 | >10 | Highly susceptible |
Table 2.
Comparison of root gall formation and nematode reproduction of M. graminicola on oat lines at 40 days postinoculation (dpi).
Table 2.
Comparison of root gall formation and nematode reproduction of M. graminicola on oat lines at 40 days postinoculation (dpi).
| Oat Line | Root Gall Formation | Nematode Reproduction | |||
|---|---|---|---|---|---|
| Total Number of Root Galls | Numbers of Incompletely Developed Root Galls | Numbers of Completely Developed Root Galls | Total Number of Eggs | Numbers of Harvested J2 | |
| LA08085BS-T2 | 17 ± 6.26 a | 11.6 ± 4.5 a | 5.4 ± 2.2 a | 325.27 ± 131.96 a | 203.4 ± 160.55 a |
| UFRGS136104-3 | 0.00 ± 0.00 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.00 ± 0.00 b | 11 ± 4.65 b |
| UFRGS136106-3 | 3 ± 0.71 b | 1.6 ± 0.7 b | 1.4 ± 0.9 b | 2.2 ± 1.16 b | 0.00 ± 0.00 b |
| UFRGS136119-2 | 2.8 ± 0.86 b | 1.4 ± 0.9 b | 1.4 ± 0.6 b | 58.4 ± 25.96 b | 125 ± 64.02 a |
Values are mean ± standard error (SE) of 5 replicates. Means within columns followed by the same letter are not significantly different (p ≥ 0.05).
Table 3.
Fresh leaf and root weight of oat lines at 40 days postinoculation (dpi) with M. graminicola.
Table 3.
Fresh leaf and root weight of oat lines at 40 days postinoculation (dpi) with M. graminicola.
| Oat Line | Leaf Weight (g) | Root Weight (g) | ||||
|---|---|---|---|---|---|---|
| Inoculated | Control | Inoculated | Control | |||
| LA08085BS-T2 | 0.138 ± 0.006 | 0.132 ± 0.008 | NS | 0.431 ± 0.017 | 0.365 ± 0.035 | NS |
| UFRGS136104-3 | 0.141 ± 0.005 | 0.151 ± 0.006 | NS | 0.589 ± 0.039 | 0.704 ± 0.051 | NS |
| UFRGS136106-3 | 0.197 ± 0.004 | 0.188 ± 0.013 | NS | 0.723 ± 0.053 | 0.701 ± 0.049 | NS |
| UFRGS136119-2 | 0.144 ± 0.015 | 0.151 ± 0.008 | NS | 0.523 ± 0.062 | 0.475 ± 0.034 | NS |
Data are mean ± standard error (SE) of 5 replicates. NS = no significant difference (p ≥ 0.05) between control and inoculated plants of each oat line.
Table 4.
Evaluation of the resistance and susceptibility of oat lines to M. graminicola based on root galls index and multiplication factor.
Table 4.
Evaluation of the resistance and susceptibility of oat lines to M. graminicola based on root galls index and multiplication factor.
| Oat Line | Root Galls Index 1 | Multiplication Factor 2 | Resistance/Susceptibility 3 |
|---|---|---|---|
| LA08085BS-T2 | 2.80 ± 0.37 a | 2.64 ± 1.36 a | MS |
| UFRGS136104-3 | 1.00 ± 0.00 b | 0.06 ± 0.02 b | HR |
| UFRGS136106-3 | 2.00 ± 0.00 b | 0.01 ± 0.01 b | R |
| UFRGS136119-2 | 1.80 ± 0.2 b | 0.92 ± 0.41 a | R |
Values are mean ± standard error (SE) of 5 replicates. Means within columns followed by the same letter are not significantly different (P ≥ 0.05); 1 Root galls index rated on a scale of 1 to 5: 1 = no gall; 2 = 1–10 galls; 3 = 11–30 galls; 4 = 31–100 galls; and 5 = more than 100 galls/root system; 2 Multiplication factor = (total number of eggs + numbers of harvested J2)/200, the initial numbers of J2 inoculation. 3 HR = highly resistant; R = resistant; MS = moderately susceptible; S = susceptible; HS = highly susceptible.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yao, S.-M.; Huang, Y.-F.; Yang, J.-I. The Screening of Resistance against Meloidogyne graminicola in Oats. Agriculture 2020, 10, 352. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10080352
AMA Style
Yao S-M, Huang Y-F, Yang J-I. The Screening of Resistance against Meloidogyne graminicola in Oats. Agriculture. 2020; 10(8):352. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10080352
Chicago/Turabian StyleYao, Shun-Min, Yung-Fen Huang, and Jiue-In Yang. 2020. "The Screening of Resistance against Meloidogyne graminicola in Oats" Agriculture 10, no. 8: 352. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10080352
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.