1. Introduction
Eggplant (
Solanum melongena L.) is commonly cultivated and consumed in Southern and Southeast Asia and has increased in popularity in Malaysia as a specialty vegetable [
1]. Globally, the eggplant is ranked as the third most important crop from the Solanaceae family after potato and tomato with an annual production of 55,197,878.00 million tons [
2]. The leading producing countries are China and India followed by Egypt, Turkey, Iran, Indonesia, Japan, Italy, Iraq and the Philippines [
2]. Rootstocks with good compatibility and tolerance or resistance to biotic and abiotic stresses are ideal for grafting vegetables and also encourage scion growth, increase production and do not decrease produce quality [
3]. Several studies have already been conducted to investigate various eggplant wild rootstocks.
Solanum paniculatum, for example, increased vigour and fruit yield while having no effect on fruit quality or composition [
4]. Tolerance to water and temperature stress was induced by
Solanum incanum [
5], while Fusarium oxysporum tolerance and Ralstonia solanacearum resistance were found in
Solanum aethiopicum and
Solanum macrocarpon [
6].
Solanum sisymbriifolium and
Solanum integrifolium have been shown to be effective in controlling bacterial wilt and yield increases [
7]. Although
Solanum torvum is the most commonly used rootstock for eggplant and has been reported to be resistant to soil-borne diseases, there is a need to find other alternative rootstocks due to a lack of rapid and homogeneous seed germination [
8]. According to Musa et al. [
9], grafting wild relatives’ rootstocks can have an impact on eggplant yield and apparent fruit quality. Grafting with resistant rootstocks proved to be an effective tool to overcome biotic and environmental stresses and increase yield and quality [
8,
10]. Presently,
Solanum torvum,
Solanum macrocarpon and
Solanum incanum have been proved as promising rootstocks effective for resistance to soil-borne diseases, tolerance to water and temperature stress, tolerance to abiotic stresses, tolerance to
Fusarium oxysporum and resistance to
Ralstonia solanacearum and are frequently used for eggplant grafting [
5,
8,
11]. The whole eggplant fruit has noticeable content of antioxidants and ranks amongst the leading vegetables in terms of antioxidant capacity [
12,
13].
Grafting vegetables with some certain rootstocks may increase total soluble solids, tataric acids and juice pH [
14,
15]. Proietti et al. [
16] reported that total vitamin C contents for grafted plants were higher than those from non-grafted plants. Research has shown the impact of genotypes on the amount and quality of the phenolic compounds found in eggplant [
17,
18]. Giorni et al. [
19] reported that eggplant grafted onto specific rootstocks had been demonstrated to increase total yield and quality of the fruit (including phytochemical content, for example, phenolic compound). Gisbert et al. [
20] reported that Cristal (eggplant) grafted onto
Solanum macrocarpon rootstocks showed a higher total phenolic content. Sabatino et al. [
21] also discovered that eggplant grafted onto
Solanum torvum enhanced total polyphenol fruit content in four landraces grown in open fields. This research was conducted to (i) assess the influence on eggplant vigour, yield, fruit proximate, physicochemical and antioxidant properties of grafted eggplant traits of wild relatives’ ST, SM and SI rootstocks and (ii) identify new potential rootstocks from eggplant wild relatives that may be useful for improving the quality of ‘ME, CE, NE and TE’ commercial varieties.
4. Discussion
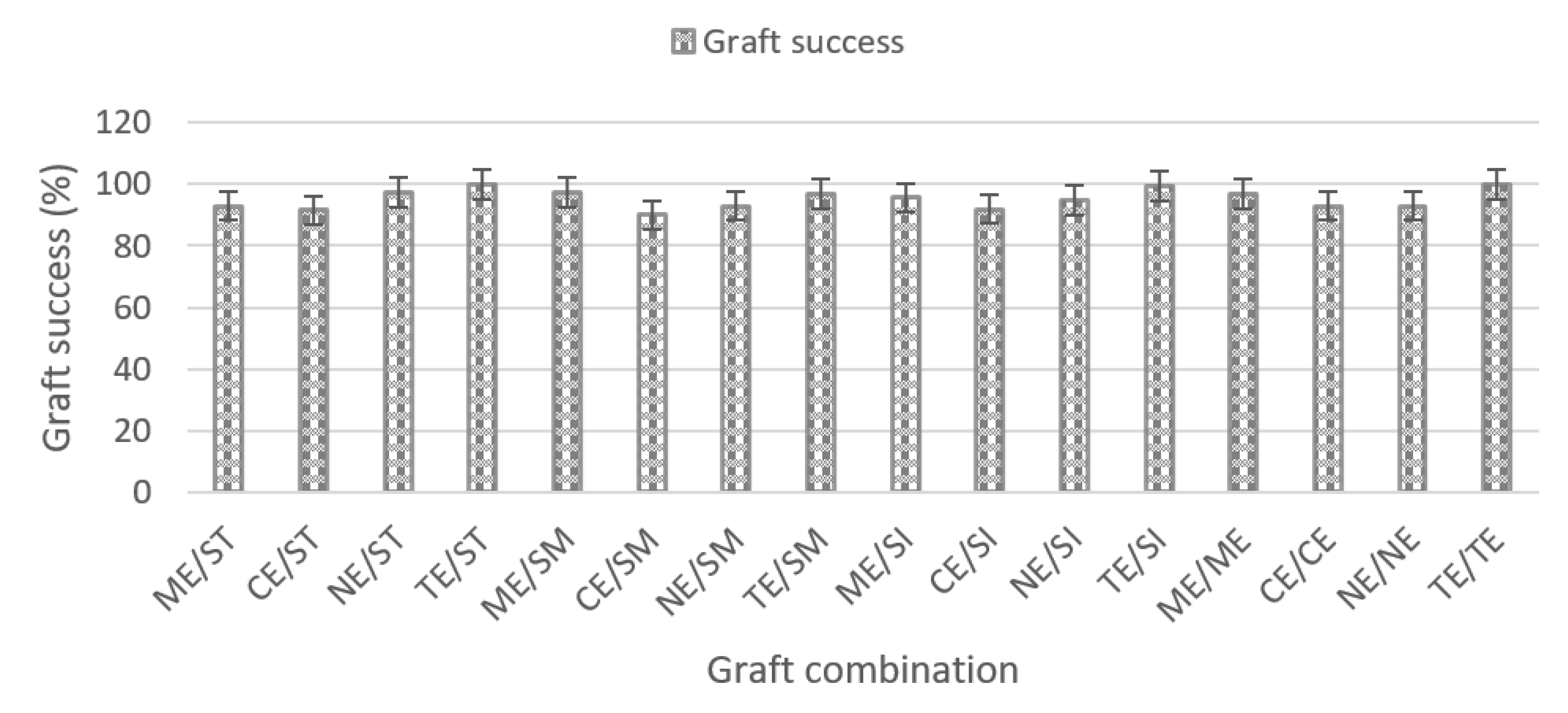
In this research, evidence showed that grafting with high-vigour rootstocks may increase the yield and quality of the fruit. The increased yield and fruit quality proven in this study is consistent with earlier reports on grafting [
9,
14,
21]. The success of grafting is determined by graft combination, rootstock and scion compatibility, temperature, humidity and light [
20,
29]. In this research, the results show that the cleft grafting technique was highly successful, with more than 90% graft success in all treatments. The result is supported by Musa et al. [
9], who reported significant graft success in MCV1, MCV2 and TCV scions grafted with wild eggplants using the cleft technique. As shown by our result, grafting with compatible rootstocks produced earlier flowers than self-rooted, for example, TE/SM, TE/ST and NE/SM. The results are consistent with the previous study of graft incompatibility and environmental stress [
9,
20]. At final harvest, plant height, which is usually regarded as a measure of vigour, was also affected by grafting. Improved vigour due to rootstock grafting in CE/ST, CE/SM and TE/SI was also evident in the values observed for plant height and number of branches, as a result of improved water uptake and greater capacity for assimilation, absorption and nutrient uptake [
9].
According to this study and earlier research, the increase in total yield of grafted plants compared with non-grafted and self-grafted plants was frequently due to the higher weight of the fruit or number of fruits. The scions CE and NE grafted onto ST rootstocks and TE scions grafted onto SM rootstocks outperformed non-grafted and self-grafted plants in terms of total yield, without any negative effects on the number of fruits per plant, since the increase in yield was determined by the number of fruits and fruit weight. Our findings are congruent with that of Sabatino et al. [
21], who observed that grafted plants had consistently more fruits per plant than non-grafted and self-grafted plants.
Higher yields in grafted plants were linked to increased water and nutrient absorption, according to Lee [
30] and Colla et al. [
31]. The result agreed with Khah et al. [
32], who observed that fruit weight was higher in grafted plants than in their non-grafted counterparts. Fruit quality is essential for marketability, and grafting can influence fruit quality attributes [
9,
14,
15,
20,
33]. Our research found that rootstocks had an impact on fruit shape parameters such as fruit length, width and fruit shape index (ME/ST, CE/ST and ME/SM, respectively). The findings are consistent with those of Gisbert et al. [
20], who showed that fruit shape changes are most likely due to the changes in the concentration of growth regulators generated by the rootstocks. In this study, Brix significantly increased in some eggplants grafted with ST, SM and SI rootstocks. ME/ST, NE/SM, TE/SM and CE/SI had higher Brix than plants from the non-grafted and self-grafted counterpart ‘ME, NE, TE and CE’ treatment, which is in accord with existing literature where a similar effect was reported by Martinez-Ballesta et al. [
34], Mohammed et al. [
35] and Flores et al. [
36]. Our result also shows that some graft combinations with vigorous rootstocks did not consistently result in increased fruit quality. These findings demonstrate that it is possible to record an increase, decrease or no changes in quality parameters of fruits from grafted plants. For example, grafted CE/SM was lower than non-grafted and self-grafted ‘CE’ treatments. This agrees with the result reported by existing literature [
14,
20,
36]. The result on fruit firmness shows a significant increase in eggplants grafted with the SM and SI rootstocks, which is in accord with existing literature reported by Lee [
30], Nkansah et al. [
37] and El-Wani et al. [
38]. The pH tends to prolong the shelf life of fresh fruits by inhibiting the multiplication of microorganisms and increasing resistance to microbial attack [
39]. The result on pH shows a significant increase in eggplants grafted with certain ST, SM and SI rootstocks and ME and CE onto ST. Improvement of fruit pH on grafting with certain rootstocks has been documented [
14,
32,
40]. Previous research has also produced inconclusive results regarding changes in fruit flavour and nutritional attributes as a result of grafting with specific rootstocks. Grafting with specific rootstocks improved some quality attributes while having neutral or negative effects on others [
36,
40,
41]. Our findings indicate that rootstocks had no effect on fruit dry matter when compared to grafting with self-grafted and non-grafted plants which produced the highest dry matter content (CE and TE), though self-grafted CE/CE was not statistically different from grafted plants CE/SI. The dry matter performance was inconsistent, depending on the rootstock and scion used, and was better in non-grafted and self-grafted plants [
20], whereas lower dry matter was recorded in grafted plants.
The findings are in accord with those obtained by Davis et al. [
14] and Turhan et al. [
42], who reported decreased dry matter in grafted fruits. However, Sabatino et al. [
33] reported a contrary observation stating that fruit dry matter produced by grafted plants was higher, compared to self- and non-grafted plants. Grafting with wild rootstocks may affect fruit fat content. Our result shows that plants grafted with wild rootstocks produced the highest fat content (ME/SI) which may be due to the selection of vigorous rootstocks, improved scions and the growing environment. Generally, changes in the proximate compositions were observed, suggesting that grafting with vigorous rootstocks may enhance the uptake of water and nutrients and growth hormone regulator balance [
41]. Fibre is one of the most important indigestible parts of food crops, such as vegetables. Results from our study indicate that the crude fibre decreased in grafted fruits, compared to control (CE/CE, NE/NE, TE/TE and TE). Based on our result, it can be concluded that crude fibre may be influenced by the selection of scions and rootstocks. Our result shows that rootstocks may not affect fruit ash content, which may be due to high mineral content in the non-grafted and self-grafted plants (TE and TE/TE). Our result is contrary to Sabatino et al. [
33] (2013), who noted that fruits harvested from grafted plants had higher ash content compared to non-grafted plants. Changes in the proximate compositions were observed, indicating that grafting may induce modifications associated with growth regulator balance.
Grafting with wild relative rootstock SM could affect fruit proximate traits such as crude protein. Results from this study are similar to those reported by Sabatino et al. [
33] and Gisbert et al. [
43], who claimed that grafted plants had higher amounts of protein in comparison with their non-grafted counterparts. Rouphael et al. [
40] also reported that fruit quality traits of grafted plants may be influenced by the scions and rootstocks selected. Changes in proximate compositions were generally observed, indicating that grafting vigorous rootstocks may induce modifications that are associated with growth regulator balance. This claim was supported by Khan et al. [
32], Gisbert et al. [
43] and Tabitha [
44] who reported that grafting with vigorous rootstocks may affect the quality of the final product [
45]. The result obtained in this study shows that the carbohydrate content decreased in grafted fruits, compared to non-grafted fruits (ME). However, the result is contrary to Gherghi et al. [
46], who reported that carbohydrate content was slightly higher in fruits produced from grafted plants in comparison to those from non-grafted plants. Similarly, Gheorghita et al. [
47] in an experiment on hybrids noted that ‘Kaiser F1′ rootstock directly influenced the content of carbohydrates.
The result on ascorbic acid showed a significantly higher level of ascorbic acid in eggplants grafted with wild rootstocks, and this was attributed to the effect of the wild rootstocks ST and SI used. This result is corroborated by Sanchez-Rodriguez et al. [
48] and Chavez-Mendoza et al. [
49], who reported that grafted fruits had higher ascorbic acid content compared with non- and self-grafted fruits. However, contrary results by Qaryouti et al. [
50] and Di Gioia et al. [
51] suggest that ascorbic acid traits were lower in grafted plants compared to un-grafted and self-grafted. The result on antioxidant activity (DPPH) shows a significant increase in the DPHH of eggplants grafted with ST and SI rootstocks. This finding is in contrast with past studies reported by Vinkovic Vrcek et al. [
52], who stated that the antioxidant activities of SIo grafts ‘Efialto’ and ‘Maxifort’ grafted onto ‘Tamaris’ were significantly lower compared to their respective rootstocks and scions. Martínez-Valverde et al. [
53] and Raffo et al. [
54] reported that antioxidant properties depend largely on lycopene content, phenolic compounds and ascorbic acid. Total flavonoid content is an antioxidant and health-related compound. Results on total flavonoid content show significantly higher flavonoid content in eggplants grafted with wild SM and ST rootstocks. This is in accord with a previous report by Nicoletto et al. [
55] who reported that under water stress, the use of a drought-tolerant rootstock (cv. Zarina) increased the concentration of total flavonoids, hydroxycinnamic acids and rutin compared to non-grafted or self-grafted ‘Zarina’.
Furthermore, some graft combinations with vigorous rootstocks showed an inconsistent increase in nutrient concentrations. Riga et al. [
56], in their study, noted that a decrease or increase in flavonoids was influenced by the choice of rootstocks when a similar scion cultivar was grafted. As shown by the drought study, rootstocks that are well adapted to stress conditions responsible for higher flavonoid production may lead to an improvement in total flavonoids in the entire plant [
48]. The phenolic compounds present in eggplant fruit made it one of the leading vegetables in terms of antioxidants [
57,
58]. Higher phenolic concentration may be a further indication of stress in the rootstock/scion combination, as stress conditions enhance the build-up of phenolics [
59,
60]. The result on total phenolic content showed more phenolics in eggplants grafted with ST and SM rootstocks. This result is similar to those obtained by Gisbert et al. [
20], Sabatino et al. [
21] and Kumar et al. [
61], who reported in their studies on fruit composition traits that higher fruit phenolic content was found in grafted plants compared to their un-grafted counterparts.


 ,
, 


{kind=link}