
Rhizosphere Microbiome Regulates the Growth of Mustard under Organic Greenhouse Cultivation

1
Hualien District Agricultural Research and Extension Station, Council of Agriculture, Jian, Hualien 97365, Taiwan
2
Department of Biology, College of Staten Island, City University of New York, 2800 Victory Boulevard, Staten Island, NY 10314, USA
3
Biochemistry and Biology Ph.D. Program, Graduate Center, City University of New York, New York, NY 10016, USA
4
Institute for Macromolecular Assemblies, City University of New York, New York, NY 10031, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agriculture 2021, 11(10), 987; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11100987
Submission received: 6 September 2021
/
Revised: 4 October 2021
/
Accepted: 7 October 2021
/
Published: 10 October 2021
(This article belongs to the Special Issue The Next Generation of Fresh Vegetables Linking Sustainability and Produce Quality Design)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Organic cultivation can improve soil fertility and biodiversity through the preservation of soil organic matter. Meanwhile, greenhouse cultivation can provide a controlled environment and therefore enables the management of every aspect of plant growth. In recent years, the combination of organic and greenhouse cultivation has slowly become a popular option in tropical regions to prevent the unpredictable impact of weather. Although it is known that organic cultivation significantly increases the density and species of microorganisms, the impact of soil microbiome on short-term vegetable growth under organic greenhouse cultivation is still not elucidated. In this study, we examined soil physiochemical properties as well as the rhizosphere microbiome from healthy and diseased mustard plants under organic greenhouse cultivation. Through next generation sequencing (NGS) analysis, our results revealed that the rhizosphere microbiome structure of healthy mustard plants was significantly different from those of the diseased mustard plants under organic greenhouse cultivation. Our findings suggest that soil microbiome composition can influence the growth of the vegetable significantly. As such, we have shown the impact of soil microbiome on vegetable growth under organic greenhouse cultivation and provide a possible strategy for sustainable agriculture.
1. Introduction
Organic farming is the fastest growing agricultural system in recent years. The vital benefit of organic farming is the preservation of soil organic composition, and thus maintaining and improving fertility, soil structure and biodiversity, and reducing erosion. Furthermore, it can also reduce the risks of human, animal, and environmental exposure to toxic chemicals. For short-term vegetable cultivation, not using chemical pesticides and synthetic fertilizers has its merits. Currently, this has become more and more popular as consumers tend to buy organic products due to the fact that organic products are free of pesticide residues. Therefore, farmers are gradually adopting the organic farming method for short-term vegetable cultivation.
It is known that organic farming significantly increases the density and species of life that exists in soil. Suitable conditions for soil fauna and flora, as well as soil formation, conditioning, and nutrient cycling are enhanced by organic practices such as manipulation of crop rotations and strip-cropping, green manuring and organic fertilization, minimum tillage, and of course, avoidance of pesticide and herbicide use. Since the soil microbiome plays an important role in almost all soil processes, the abundance, activity, and composition of microorganisms will generally determine the productivity of agricultural land [1,2].
Research has shown that organic farming systems increased taxonomic, phylogenetic richness, diversity, and heterogeneity of soil microbiota compared with conventional farming methods [3,4]. However, when soil structure and pH were similar, microbial communities in conventional and organic farming systems revealed no major differences among the main bacterial phyla [5]. The soil microbiome surrounding roots or inside the root system is diverse and plays an important role in conferring tolerance to abiotic stresses of plants [6].
Although organic production offers farmers increased control over their crops, they face great challenges in pest management. For example, many pests and diseases such as the striped flea beetle (Phyllotreta striolata) or damping-off disease caused by Rhizoctonia solani have made organic farming difficult in open fields [7]. On the other hand, greenhouse cultivation gives farmers the chance to create their own growing environment, and therefore farmers can manage every aspect of production. Greenhouse cultivation protects crops from a host of pests and diseases, extreme weather conditions, and outdoor pollution such as pesticide drift. As such, greenhouse cultivation has become an alternative option to avoid pathogen reproduction and dispersal caused by increased humidity due to rainfall, and to prevents contacts with harmful pests.
In organic farming, pH is considered as one of major drivers in bacterial diversity, being highest in near neutral soils and lower in acidic soils [8,9]. A previous study demonstrated that continuous planting of vegetables in greenhouses can lead to soil acidification and salinization, thus reducing yield and quality of tomatoes significantly [10]. This may be due to the decrease of soil microbiome diversity and richness. Furthermore, bacterial diversity in the intensely cultivated greenhouse soils was also influenced by electrical conductivity which is associated with soil salinity [11]. Therefore, it is important to evaluate the impact of the soil microbiome on crop growth under organic greenhouse cultivation.
Although the soil microbiome plays an important role in agricultural production, most soil bacteria cannot be successfully cultivated in vitro [12]. We still have very limited knowledge about the complexity of the soil microbiome. Through next generation sequencing (NGS) analysis of soil microbiomes, individual bacterial taxa can be explored, including bacteria with growth-promoting or growth-compromising activities. In this study, we examined soil physiochemical properties as well as the soil microbiome from healthy and diseased mustard grown using organic greenhouse cultivation. This information will enable us to have a better understanding of soil microbial structure so that we can manage the soil environment by promoting beneficial microbes and suppressing detrimental microbes.
2. Materials and Methods
2.1. Place of Research and the Collection of Samples for Microbiota Profiling
The research was conducted in Hualien County, Taiwan (coordinates 23°53′54.92″, 121°31′22.80″) from November to December in 2019. Mustard (Brassica juncea cv. yuanjen) is a local fresh leafy vegetable variety. Normally, the suitable sowing time for mustard is between September and February of the following year while the temperature is 18–30 °C. For this research, mustard seeds were sown and germinated on 128-hole seedling trays (60 × 30 × 3.5 cm; 20 g soil/hole) under natural light until the seedlings reached four expanded leaves, and then transplanted into the greenhouse. The greenhouse is a manually operated greenhouse. Under normal circumstances, the natural light is used, and the sunshade net is used only when the temperature is above 32 °C During the period of this study, the average temperature for November 2019 and December 2019 were 22.08 °C and 19.44 °C, respectively. Meanwhile, the relative humidity for November 2019 and December 2019 were 78.5 and 82.0, respectively. Therefore, the greenhouse environment was not required to be controlled throughout the experiment.
Prior to planting the mustard, the soil was submerged with water and then drained completely twice to remove excessive salt from previous cultivation. Subsequently, organic fertilizer containing 80% organic matter, 5% nitrogen, 2.6% phosphorus, and 2% potassium was applied at 0.24 kg/m2 according to the manufacturer’s recommendations.
We collected root soil from the depth between 5 cm and 15 cm of three randomly chosen healthy (OK) areas’ and three randomly chosen diseased (NG) areas’ root systems at 7 days after transplanting. Some mustard plants showed symptoms within 7 days, including slow growth, yellowing and even death. These symptoms were not caused by pathogens or pests (data not shown). These areas were designated as NG areas (Figure S1). On the other hand, normal mustard growth areas where mustard plants showed no symptoms were OK areas. Therefore, we had 3 replications for each condition and 10 mustard roots were pooled in each replicate. Soil genomic DNA was extracted from 250 mg of each rhizosphere soil sample using the Qiagen DNeasy@ Powersoil kit (Qiagen, Hilden, Germany).
2.2. PCR Amplification, Sequencing, and Statistical Analysis
Targeted metagenomic profiling of the samples was carried out by sequencing the V4 region of the 16S rRNA gene for bacteria. Briefly, amplicon libraries were created using barcode-tagged primers for the primer pairs 515F/806R (515F: 5′-GTGCCAGCMGCCGCGGTAA-3′; 806R: 5′-GGACTACHVGGGTWTCTAAT-3′). These primers were used in quantitative PCR of DNA extracts. Sequencing was performed pair-end 2 × 250 bp by Illumina MiSeq Instrument. A total of 495,466 high quality reads were generated and the minimum frequency was 40,996, with an average read length of 253 bp. The high-throughput data was analyzed using Kraken 2 with MiniKraken v2 database for initial bacterial identification. Subsequently, all statistical analysis were performed in Qiime 2 (Ver. 2019.10) using Silva_138.
2.3. Soil Physicochemical Properties
For soil pH, air-dried rhizosphere soil sample was resuspended in deionized water (1:1, w/v) and the pH value was determined by TOA-DDK HM-25G pH Meter (Tokyo, Japan). The conductivity of soil suspension (1:5, w/v) filtrate was measured by conductivity meter (model CM-60S, TOA-DKK, Tokyo, Japan). The content of organic matter was assessed by the Vario Max C elemental analyzer (Elementar Analysensysteme GmbH, Hanau, Germany). Available P was determined by molybdenum blue colorimetric method [13,14]. Exchangeable potassium, calcium, and magnesium were quantified by inductively coupled plasma atomic emission spectrometry (ULTIMA-2000, HORIBA Jobin Yvon, Longjumeau, France).
3. Results
3.1. Microbial Biodiversity within Rhizosphere Soil Communities
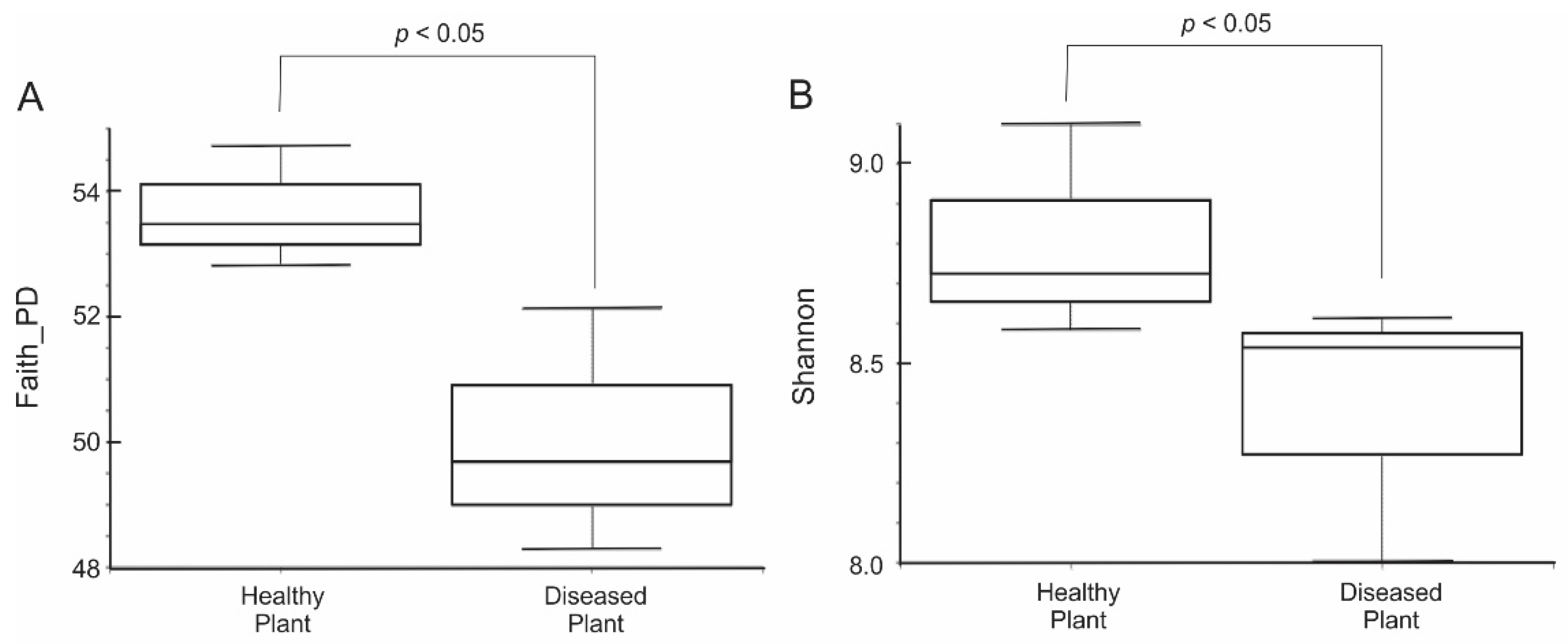
To examine whether the rhizosphere microbiome plays an important role in mustard (Brassica juncea cv. yuanjen) plant’s growth, rhizosphere soil samples were collected for DNA extraction, Illumina MiSeq NGS process and Qiime2 data analysis. Shannon rarefaction curve was calculated by randomly re-sampling each sample multiple times and validated the minimum number required for downstream analysis. The results showed that the minimum rarefied number is under 20,000 (Figure S2), and it demonstrated that the rhizosphere microbiome from diseased mustard (NG) is lower than that from the healthy mustard (OK). To retain maximum features for analysis, the sampling depth of 36,000 per sample was chosen to perform core metrics analysis. For alpha diversity analysis, both Faith’s phylogenetic diversity and Shannon diversity index were used to determine the within sample diversity. Both analyses showed that OK has a significantly higher bacterial diversity than NG (Figure 1A,B). This result suggests that the rhizosphere microbiome from healthy mustard plants had a higher number of operational taxonomic units (OTUs) and the species are more evenly distributed than the rhizosphere microbiome from diseased mustard plants.
3.2. The Rhizosphere MMicrobiomes of Healthy and Diseased Mustard Plant Communities
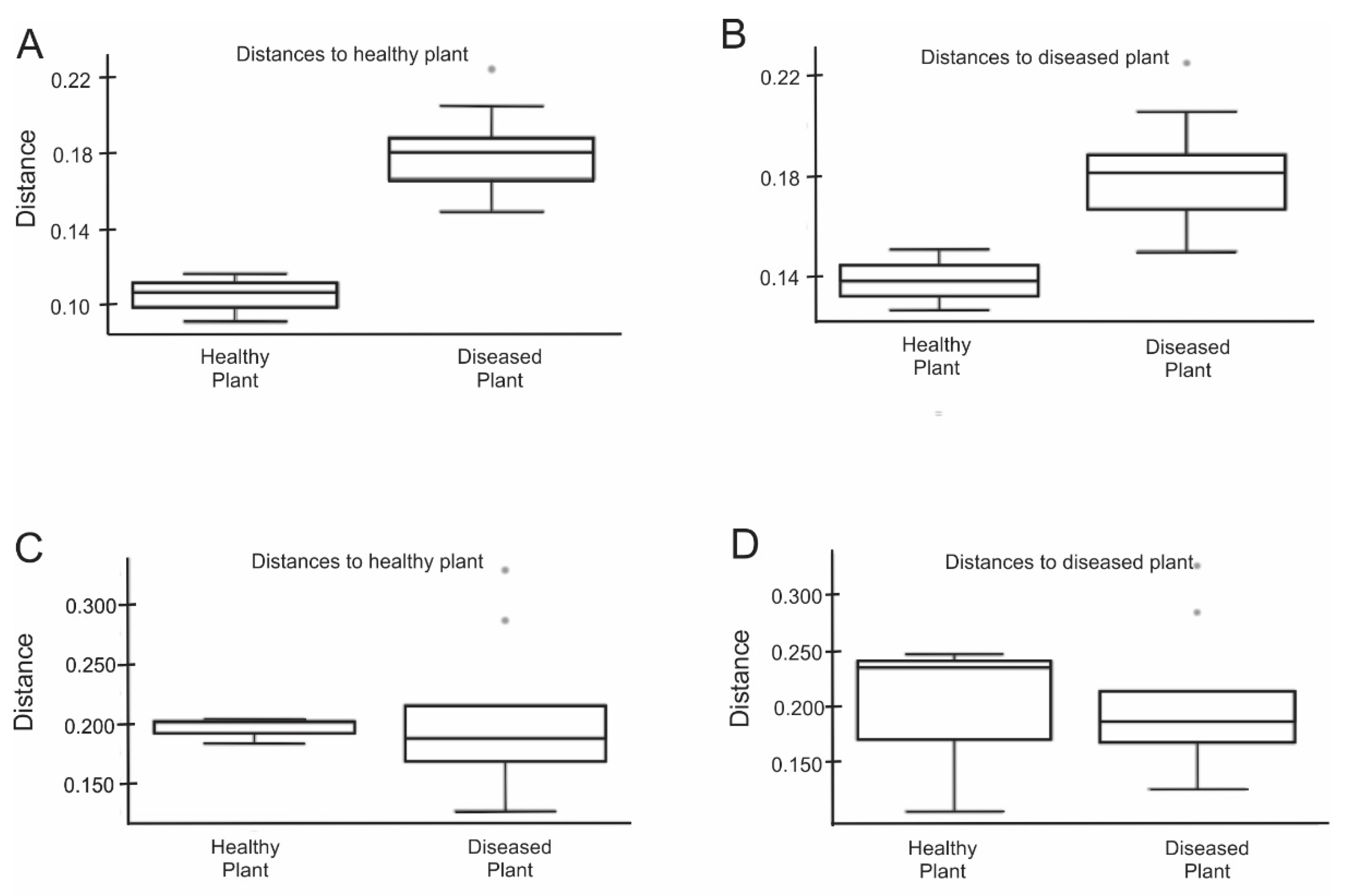
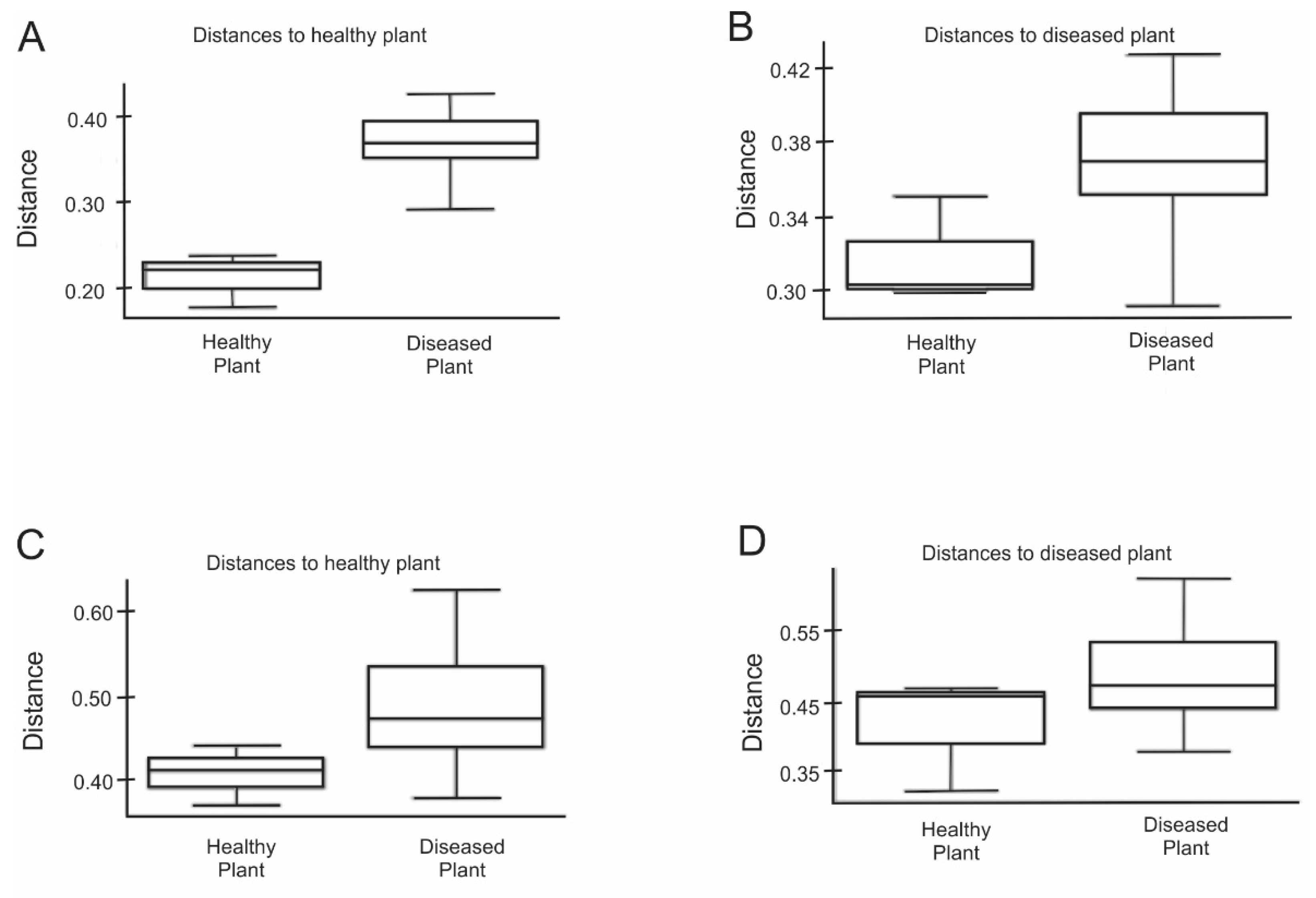
Next, we examined the biodiversity variation between the healthy rhizosphere microbiome and diseased rhizosphere microbiome. We found 308 unique OTUs and 84 unique OTUs in OK and NG rhizosphere, respectively, while 3811 OTUs were shared between both rhizospheres (Figure S3). Subsequently, principal coordinate analysis (PCoA) was used to examine global differences in microbial community structure. PCoA of unweighted UniFrac distances separated healthy microbiome from those of diseased microbiome. The unweighted UniFrac ANOSIM analysis displayed significant difference between two communities (R = 0.96, p < 0.1), indicating presence or absence of unique OTUs in either OK or NG (Figure 2A,B). PCoA of weighted UniFrac distances did not show significant difference between healthy rhizosphere microbiome and diseased rhizosphere microbiome. Furthermore, the weighted UniFrac ANOSIM analysis also revealed that the communities were not significantly different (R = 0.074, p > 0.1) (Figure 2C,D). However, both Jaccard analysis and Bray–Curtis PERMANOVA showed significant differences between the two communities (p < 0.1; Figure 3). Overall, beta diversity analysis suggested that rhizosphere microbiome structure of healthy mustard plants was remarkably different from those of the diseased mustard plants.
3.3. The Composition of Bacterial Taxa between the Healthy and Diseased Rhizosphere Communities
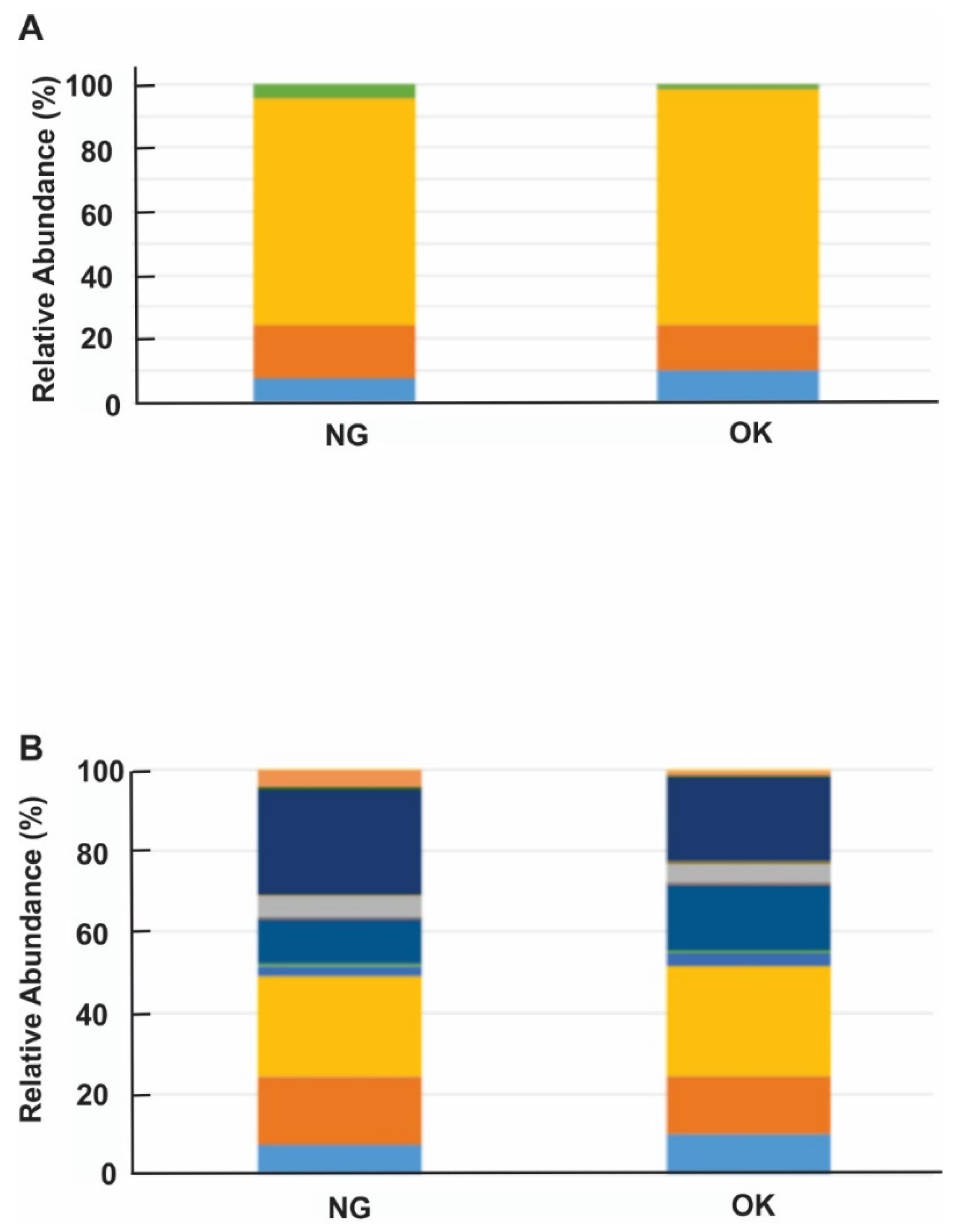
Since we found both healthy and diseased rhizosphere microbiome structures are significantly different, we next examined the bacterial composition in these two communities. In both OK and NG communities, the predominant bacterial phyla were Proteobacteria, Actinobacteria, Verrucomicrobia, Planctomycetes, and SBR1093, but with different relative abundance (Figure 4A; Figure S4). Within these phyla, the mean relative abundance of Proteobacteria and SBR1093 were significantly higher in OK rhizosphere than in NG rhizosphere. At order level, the most relative abundant orders in OK soil samples were Pseudomonadales, Desulfuromonadales, and Actinomycetales. The same distribution was also noted in NG soil, but Pseudomonadales appeared to be the most abundant order, followed by Actinomycetales and Desulfuromonadales (Figure 4B).
At lower taxonomic levels, excluding unknown, a total of 23 genera were significantly higher in OK soil than in NG soil, including Dyella and Pseudonocardia (Figure S5). In contrast, only three genera were significantly higher in diseased microbiome than in healthy microbiome. Interestingly, 14 genera were Proteobacteria, while 5 genera were Actinobacteria. At species level, 1128 species were at least identified once (Table S1). Analysis of a subset of OTUs revealed 53 bacterial species were significantly different between OK and NG in relative abundance (p < 0.05) and had absolute abundance greater or equal to 10 in at least one sample (Table S2). As such, our results clearly demonstrated that difference in the microbiome structure can have impact on mustard plants’ growth conditions.
4. Discussion
The major advantage of growing vegetables in greenhouses is to avoid the external environmental effects. For example, vegetables can be shaded in hot summer to reduce the direct exposure from the sun and thus protect vegetables from the heat damage. Furthermore, vegetables can be prevented from the impact of tropical storms, such as typhoons or hurricanes, which often occur during the summer in tropical regions. Greenhouses can also reduce chilling damage during the winter. In contrast, the disadvantage of greenhouse cultivation is the lack of rainwater washing. As a result, high soil salinity is usually a problem for greenhouse cultivation affecting the growth of the vegetables dramatically.
In the present study we cultivated mustard plants under the greenhouse conditions and examined the soil microbiome and soil physicochemical condition between the healthy and diseased mustard to further understand the relationship within the soil conditions, soil microbiome, and vegetable growth under organic greenhouse cultivation. Analysis of the physicochemical properties of soil samples revealed higher conductivity in soils with diseased plants (NG) than those growing healthy plants (OK) except NG2 soil (Table S3). We observed that NG1 had the highest electrical conductivity (EC(1:5)) at 2.02 dS/m, followed by NG3 with 1.32 dS/m. It is known that the salinity threshold (ECt) of most vegetable plants is low, ranging from 1 to 2.5 dS/m in saturated soil extracts [15]. When converted to EC(1:5), the threshold is approximately 0.2–0.5 dS/m depending on different vegetable crops. Therefore, it is possible that the poor growth of NG1 and NG3 may have resulted from the high electrical conductivity. From beta diversity correlation analysis, it showed weak significant correlation between the electrical conductivity and physiological growth states (Table S4). Nevertheless, EC(1:5) of NG2 was lower than OK1 and OK2, with a value of 0.25 dS/m, indicating other factors may contribute to plant growth.
In terms of the soil organic matter, we found that only NG3 had organic matter of 4.6% while others were less than 2%, and there was no significant difference between OK and NG mustard. Beta diversity correlation analysis did not show strong correlation between organic matter and mustard physiological growth states (Table S4). However, a significant difference between OK and NG was the abundance of Candidatus Koribacter versatilis found (p < 0.05; Table S2). Candidatus Koribacter versatilis contains abundant gene clusters encoding for modules of the carbohydrate-active enzyme family, as well as glycoside hydrolases and glycosyl transferases. These enzymes are involved in the breakdown, utilization and biosynthesis of diverse structural and storage polysaccharides and resistance to fluctuating temperature and nutrient deficient conditions [16]. Although it has been shown that the abundance of this taxon was correlated with the low nutrient inputs due to the adaption to a nutrient-limited environment [3,17], our results indicated that OK soil contains high abundance of Candidatus Koribacter versatilis, suggesting that the abundance of Candidatus Koribacter versatilis is irrelevant to the content of organic matter. Since healthy rhizosphere soil contains high abundance of Candidatus Koribacter versatilis, it might be one of the major factors contributing to the mustard plants’ growth under current study.
From beta diversity correlation analysis, we found a significant strong correlation between the pH and mustard growth states (Table S4). This finding suggests the mutual interaction between the soil microbiome and the soil pH, and this strong interaction contributes significantly to the mustard growth. To further explore the importance of this correlation, we examined the OUT collected from these two rhizospheres. OTUs classified at lower taxonomic level reveal in-depth information to infer the putative ecological role of the taxon. At the genus level, for example, we found that Dyella was significantly different between OK and NG soils (Figure S5). Dyella has been identified in rhizosphere of several plant species with the ability to modulate plant growth or to produce antibiotics. In soybean, Dyella was positively correlated with the nodule number, nodule biomass and nitrogenase activity, and negatively correlated with the available nitrogen [18]. Dyella also showed plant growth-promoting activities in nodules of both Lespedeza sp. and Phaseolus vulgaris [19,20]. During phosphate limitation in maize, D. japonica upregulates biosynthesis of phenazine, a natural bacterial antibiotic that can protect plants from disease [21]. As such, our finding indicates that Dyella might positively contribute to mustard’s growth under organic greenhouse cultivation.
From the assigned OTUs, we found 53 species that were significantly different between OK and NG in relative abundance (Table S2). These included 26 species of Proteobacteria and 9 species of Firmicutes. Among them, Bacillus is one of the genera in Firmicutes that has been studied intensively. Bacillus usually has many beneficial functions including phosphate solubilization, phytohormone production and protection against biotic and abiotic stresses [22]. Here, we show that OK had significantly higher Bacillus thuringiensis. It has been shown that Bacillus thuringiensis is commonly used in biological control against Lepidoptera larvae [23]. In addition, B. thuringiensis is a successful endophyte in many plants and can promote growth by suppressing diseases [24]. Another recent study also found that B. thuringiensis NEB17, isolated from soybean nodules, can produce a single small peptide thuricin 17 which can stimulate plant growth during abiotic stresses [25]. In fava bean, B. thuringiensis can exert indole acetic acid, produce exopolysaccharides, and fix nitrogen, thus is effective in promoting plant growth [26]. Therefore, our results coincide with these findings in which plant growth condition is positively related to the abundance of B. thuringiensis.
Within Firmicutes, Symbiobacterium thermophilum is a syntrophic bacterium whose growth depends on coculture with a Bacillus sp. Compounds 2,2-bis (3′-indolyl)indoxyl (BII) and 1,1-bis(3′-indolyl)ethane (BIE) were identified as self-growth inhibitors of S. thermophilum. It has been suggested that Bacillus benefits S. thermophilum by detoxifying BII and BIE [27]. In the current study, we found that S. thermophilum was significantly higher in OK than in NG. Furthermore, Bacillus spp. was also more abundant in OK than in NG. Therefore, our results suggest the importance of these bacteria in plant growth.
We found 29 Bacillus species in our analysis and the majority of these species were not significantly different between OK and NG, except Bacillus sp. Y1 and Bacillus thuringiensis. However, we did observe that B. cereus, B. megaterium, and B. coagulans were more abundant in OK than in NG despite being statistically insignificant (Table S5). In maize, B. cereus C1L acts as a plant growth-promoting rhizobacterium by eliciting induced systemic resistance against southern corn leaf blight [28]. In potato, B. megaterium showed not only antimicrobial activity against Streptomyces scabies and Erwinia carotovora subsp. atroseptica, but also inhibitory activity on the endotoxin solanine which is known to influence distribution of the endophytic bacterial community [29]. Some bacteria associated with arbuscular mycorrhizal fungi are referred to as mycorrhiza helper bacteria. Synergistic interactions between B. coagulans and arbuscular mycorrhizal fungi promoted plant growth compared to single inoculation with either of them [22]. As such, plant growth-promoting bacterium can also contribute to the difference between healthy and diseased mustard.
The plant rhizosphere is a highly competitive environment. The relative abundance of each microorganism fluctuates dynamically as the environment changes. Although the population size of B. subtilis and B. amyloliquefaciens were very low in both OK and NG, these microorganisms may thrive when conditions become suitable. Members of B. subtilis and B. amyloliquefaciens have been developed as biocontrol agents against various plant diseases. In soybean, B. subtilis RSS-1 and B. amyloliquefaciens JDF3 can inhibit mycelial growth, the germination of the cysts, and the swimming of the motile zoospores of Phytophthora sojae. In a pot experiment, B. subtilis and B. amyloliquefaciens can enhance the resistance of soybean to P. sojae [30]. B. subtilis is a remarkably diverse bacterial species that can grow within myriad environments. Like all members of the genus Bacillus, B. subtilis can form endospores that are capable of surviving nutrient deprivation and environmental stresses [31]. The biocontrol mechanism of B. amyloliquefaciens FZB42 is likely by stimulating induced systemic resistance [32].
The root exudates produced by plants may change their growth environment, biotic or abiotic properties, and thereby affect the growth and health of current and future plant generations [33,34]. Furthermore, small initial variation in soil microbiome composition and function can predetermine whether plants survived or become diseased [35]. Therefore, it is very likely that microbiome structure from current cultivation can remain in the soil and influence the next cultivation. Indeed, we did observe such impact when we grew cabbage seedlings in the diseased mustard soil. We found those transplanted to NG diseased soil grew more poorly than in the control soil (data not shown). Thus, the microbiome in NG soil was likely to be one of the major factors that hindered the growth of cabbage, and the preexisting microbiome structure influenced the growth of the next cultivation.
5. Conclusions
In this study, we examined the effect of organic greenhouse cultivation on short-term vegetable growth through soil physicochemical analysis and rhizosphere microbiome NGS analysis. Although the soil physicochemical properties might not have had a significant impact on soil microbiome, our results indicate that the significant difference in rhizosphere microbiome contribute to the physiological growth state of vegetables. Furthermore, results suggest the microbiome structure remains in the soil under greenhouse environment and thus influences the growth state of the next cultivation. Our findings have provided the evidence that microbiome structure is an important factor to control vegetable growth under organic greenhouse cultivation.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/agriculture11100987/s1, Figure S1: Organic greenhouse cultivation of mustard (Brassica juncea cv. yuanjen), Figure S2: Rarefaction analysis, Figure S3: Differences in bacterial operational taxonomic units (OTUs) between roots from healthy (OK) and diseased (NG) plants, Figure S4: Heatmap showing the relative abundance of bacterial OTUs at phylum level, Figure S5: Significantly different genera between conditions (log2 fold change OK to NG), Table S1: NGS analysis of OUT abundance, Table S2: Statistical test on OUT relative abundance, Table S3: Physicochemical properties of rhizosphere soil, Table S4: Beta diversity correlation analysis, Table S5: Statistical analysis of abundance in Bacillus genus.
Author Contributions
T.-C.L. perceived the project, prepared all samples, performed biochemical and physicochemical analysis. C.-H.S., T.-C.L., H.-M.P. and S.W. conducted data analysis. S.W. developed software codes. C.-H.S., T.-C.L., and H.-M.P. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Science and Technology Development Fund of the Ministry of Science and Technology, Taiwan, (MOST 109-3111-Y-067E-001) to T.-C.L. The APC was funded by MOST 109-3111-Y-067E-001.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data is contained within the article or supplementary material.
Acknowledgments
This research was supported by the National Science and Technology Development Fund (MOST 109-3111-Y-067E-001) of the Ministry of Science and Technology, R.O.C. (Taiwan) to T.-C.L., and by PSC-CUNY awards (635450051 and 644680052), and CUNY Institute for Macromolecular Assemblies research support to C.-H.S.
Conflicts of Interest
The authors declare no competing interests.
References
- van der Heijden, M.G.A.; Bardgett, R.D.; Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Walters, K.E.; Martiny, J.B. Alpha-, beta-, and gamma-diversity of bacteria varies across habitats. PLoS ONE 2020, 15, e0233872. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [Green Version]
- Lupatini, M.; Korthals, G.W.; de Hollander, M.; Janssens, T.K.S.; Kuramae, E.E. Soil microbiome is more heterogeneous in organic than in conventional farming system. Front. Microbiol. 2017, 7, 2064. [Google Scholar] [CrossRef] [Green Version]
- Armalytë, J.; Skerniškytë, J.; Bakienë, E.; Krasauskas, R.; Šiugždinienë, R.; Kareivienë, V.; Kerzienė, S.; Klimienė, I.; Sužiedėlienė, E.; Ružauskas, M. Microbial diversity and antimicrobial resistance profile in microbiota from soils of conventional and organic farming systems. Front. Microbiol. 2019, 10, 892. [Google Scholar] [CrossRef]
- Hartman, K.; Tringe, S.G. Interactions between plants and soil shaping the root microbiome under abiotic stress. Biochem. J. 2019, 476, 2705–2724. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.C.; Yu, S.W. Occurrence and Management of Diseases and Pests of Cruciferous Vegetables (in Chinese); Taiwan Agricultural Chemicals and Toxic Substances Research Institute: Taichung, Taiwan, 2012; pp. 14–36.
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Cheng, Z.; Meng, H.; Ahmad, I.; Zhao, H. Growth, yield and quality of spring tomato and physicochemical properties of medium in a tomato/garlic intercropping system under plastic tunnel organic medium cultivation. Sci. Hortic. 2014, 170, 159–168. [Google Scholar] [CrossRef]
- Kim, J.M.; Roh, A.S.; Choi, S.C.; Kim, E.J.; Choi, M.T.; Ahn, B.K.; Kim, S.; Lee, Y.; Joa, J.; Kang, S.; et al. Soil pH and electrical conductivity are key edaphic factors shaping bacterial communities of greenhouse soils in Korea. J. Microbiol. 2016, 54, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Girard, R.A.; Martin, T.; Edwards, J.; Thrash, J.C. Status of the archaeal and bacterial census: An update. mBio 2016, 7, e00201–e00216. [Google Scholar] [CrossRef] [Green Version]
- He, Z.L.; Baligar, V.; Ritchey, K.; Martens, D. Determination of soluble phosphorus in the presence of organic ligands or fluoride. Soil Sci. Soc. Am. J. 1998, 62, 1538–1541. [Google Scholar] [CrossRef]
- Wei, L.L.; Chen, C.R.; Xu, Z.H. The effect of low-molecular-weight organic acids and inorganic phosphorus concentration on the determination of soil phosphorus by the molybdenum blue reaction. Biol. Fertil. Soils 2009, 45, 775–779. [Google Scholar] [CrossRef]
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Rawat, S.R.; Männistö, M.K.; Bromberg, Y.; Häggblom, M.M. Comparative genomic and physiological analysis provides insights into the role of Acidobacteria in organic carbon utilization in Arctic tundra soils. FEMS Microbiol. Ecol. 2012, 82, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Ward, N.L.; Challacombe, J.F.; Janssen, P.H.; Henrissat, B.; Coutinho, P.M.; Wu, M.; Xie, G.; Haft, D.H.; Sait, M.; Badger, J.; et al. Three genomes from the phylum Acidobacteria provide insight into the lifestyles of these microorganisms in soils. Appl. Environ. Microbiol. 2009, 75, 2046–2056. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Gu, C.; Liu, X.; Yang, C.; Li, W.; Wang, S. Impact of soybean nodulation phenotypes and nitrogen fertilizer levels on the rhizosphere bacterial community. Front. Microbiol. 2020, 11, 750. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, P.; Chauhan, P.S.; Saravanan, V.S.; Anandham, R.; Sa, T. Isolation and characterization of plant growth promoting endophytic bacterial isolates from root nodule of Lespedeza sp. Biol. Fertil. Soils 2010, 46, 807–816. [Google Scholar] [CrossRef]
- Tapia-García, E.Y.; Hernández-Trejo, V.; Guevara-Luna, J.; Rojas-Rojas, F.U.; Arroyo-Herrera, I.; Meza-Radilla, G.; Vásquez-Murrieta, M.S.; Los Santos, P.E. Plant growth-promoting bacteria isolated from wild legume nodules and nodules of Phaseolus vulgaris L. trap plants in central and southern Mexico. Microbiol. Res. 2020, 239, 126522. [Google Scholar] [CrossRef] [PubMed]
- Dar, D.; Thomashow, L.S.; Weller, D.M.; Newman, D.K. Global landscape of phenazine biosynthesis and biodegradation reveals species-specific colonization patterns in agricultural soils and crop microbiomes. eLife 2020, 9, e59726. [Google Scholar] [CrossRef] [PubMed]
- Nanjundappa, A.; Bagyaraj, D.J.; Saxena, A.K.; Kumar, M.; Chakdar, H. Interaction between arbuscular mycorrhizal fungi and Bacillus spp. in soil enhancing growth of crop plants. Fungal Biol. Biotechnol. 2019, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capella, T.; Christou, P. Bacillus thuringiensis: A century of research, development and commercial applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azizoglu, U. Bacillus thuringiensis as a biofertilizer and biostimulator: A mini-review of the little-known plant growth-promoting properties of Bt. Curr. Microbiol. 2019, 76, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M.; Smith, D.L. A PGPR-produced bacteriocin for sustainable agriculture: A review of thuricin 17 characteristics and applications. Front. Plant Sci. 2020, 11, 916. [Google Scholar] [CrossRef]
- Saad, M.M.G.; Kandil, M.; Mohammed, Y.M.M. Isolation and Identification of plant growth-promoting bacteria highly effective in suppressing root rot in fava beans. Curr. Microbiol. 2020, 77, 2155–2165. [Google Scholar] [CrossRef]
- Watsuji, T.; Yamada, S.; Yamabe, T.; Watanabe, Y.; Kato, T.; Saito, T.; Ueda, K.; Beppu, T. Identification of indole derivatives as self-growth inhibitors of Symbiobacterium thermophilum, a unique bacterium whose growth depends on coculture with a Bacillus sp. Appl. Environ. Microbiol. 2007, 73, 6159–6165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.J.; Yang, K.H.; Liu, Y.H.; Lin, Y.J.; Chen, C.Y. Suppression of southern corn leaf blight by a plant growth promoting rhizobacterium Bacillus cereus C1L. Ann. Appl. Biol. 2010, 157, 45–53. [Google Scholar] [CrossRef]
- Liu, J.; Wang, S.; Zheng, X.; Jin, N.; Lu, J.; Huang, Y.; Fan, B.; Wang, F. Antimicrobial activity against phytopathogens and inhibitory activity on solanine in potatoes of the endophytic bacteria isolated from potato tubers. Front. Microbiol. 2020, 11, 570926. [Google Scholar] [CrossRef]
- Liu, D.; Li, K.; Hu, J.; Wang, W.; Liu, X.; Gao, Z. Biocontrol and action mechanism of Bacillus amyloliquefaciens and Bacillus subtilis in soybean phytophthora blight. Int. J. Mol. Sci. 2019, 20, 2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, A.M.; Losick, R.; Kolter, R. Ecology and genomics of Bacillus subtilis. Trends Microbiol. 2008, 16, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42–a review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [Green Version]
- Kulmatiski, A.; Kardol, P. Getting Plant—Soil feedbacks out of the greenhouse: Experimental and conceptual approaches. In Progress in Botany; Lüttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin, Heidelberg, 2008; Volume 69, pp. 449–472. [Google Scholar] [CrossRef]
- Song, C.; Zhu, F.; Carrión, V.J.; Cordovez, V. Beyond plant microbiome composition: Exploiting microbial functions and plant traits via integrated approaches. Front. Bioeng. Biotechnol. 2020, 8, 896. [Google Scholar] [CrossRef]
- Wei, Z.; Gu, Y.; Friman, V.P.; Kowalchuk, G.A.; Xu, Y.; Shen, Q.; Jousset, A. Initial soil microbiome composition and functioning predetermine future plant health. Sci. Adv. 2019, 5, eaaw0759. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Low bacterial diversity in diseased field through alpha diversity analysis. (A) Faith’s phylogenetic diversity (H index: 3.85 and p < 0.05); (B) Shannon diversity index (H index: 3.86 and p < 0.05). Healthy plant: OK; diseased plant: NG.
Figure 1.
Low bacterial diversity in diseased field through alpha diversity analysis. (A) Faith’s phylogenetic diversity (H index: 3.85 and p < 0.05); (B) Shannon diversity index (H index: 3.86 and p < 0.05). Healthy plant: OK; diseased plant: NG.

Figure 2.
Significant differences in microbial community composition between healthy and diseased rhizosphere microbiome through UniFrac ANOSIM analysis. (A) Unweighted UniFrac distance to healthy plant; (B) unweighted UniFrac distance to diseased plant; (C) weighted UniFrac distance to healthy plant; (D) weighted UniFrac distance to diseased plant. *, **: sample outlier.
Figure 2.
Significant differences in microbial community composition between healthy and diseased rhizosphere microbiome through UniFrac ANOSIM analysis. (A) Unweighted UniFrac distance to healthy plant; (B) unweighted UniFrac distance to diseased plant; (C) weighted UniFrac distance to healthy plant; (D) weighted UniFrac distance to diseased plant. *, **: sample outlier.

Figure 3.
Significant differences in microbial community composition between healthy and diseased rhizosphere microbiome through Jaccard and Bray–Curtis PERMANOVA. (A) Jaccard distance to healthy plant (OK); (B) Jaccard distance to diseased plant (NG); (C) Bray–Curtis distance to healthy plant (OK); (D) Bray–Curtis distance to diseased plant (NG).
Figure 3.
Significant differences in microbial community composition between healthy and diseased rhizosphere microbiome through Jaccard and Bray–Curtis PERMANOVA. (A) Jaccard distance to healthy plant (OK); (B) Jaccard distance to diseased plant (NG); (C) Bray–Curtis distance to healthy plant (OK); (D) Bray–Curtis distance to diseased plant (NG).

Figure 4.
Stacked bar plots of relative abundance of bacterial (A) phyla and (B) order. (A) ![Agriculture 11 00987 i001]() : Verrucomicrobia;
: Verrucomicrobia; ![Agriculture 11 00987 i002]() : Proteobacteria;
: Proteobacteria; ![Agriculture 11 00987 i003]() : Actinobacteria;
: Actinobacteria; ![Agriculture 11 00987 i004]() : SBR1093;
: SBR1093; ![Agriculture 11 00987 i005]() : Planctomycetes;
: Planctomycetes; ![Agriculture 11 00987 i006]() : unknown. (B)
: unknown. (B) ![Agriculture 11 00987 i003]() : Actinobacteria_Actinobacteria_Actinomycetales;
: Actinobacteria_Actinobacteria_Actinomycetales; ![Agriculture 11 00987 i005]() : Planctomycetes_OM190_agg27;
: Planctomycetes_OM190_agg27; ![Agriculture 11 00987 i002]() : Proteobacteria;
: Proteobacteria; ![Agriculture 11 00987 i007]() : Proteobacteria_Deltaproteobacteria;
: Proteobacteria_Deltaproteobacteria; ![Agriculture 11 00987 i008]() : Proteobacteria_Deltaproteobacteria_Desulfuromonadales;
: Proteobacteria_Deltaproteobacteria_Desulfuromonadales; ![Agriculture 11 00987 i001]() : Proteobacteria_Deltaproteobacteria_Desulfovibrionales;
: Proteobacteria_Deltaproteobacteria_Desulfovibrionales; ![Agriculture 11 00987 i009]() : Proteobacteria_Deltaproteobacteria_NB1-j;
: Proteobacteria_Deltaproteobacteria_NB1-j; ![Agriculture 11 00987 i010]() : Proteobacteria_Epsilonproteobacteria_Campylobacteriales;
: Proteobacteria_Epsilonproteobacteria_Campylobacteriales; ![Agriculture 11 00987 i011]() : Proteobacteria_Gammaproteobacteria;
: Proteobacteria_Gammaproteobacteria; ![Agriculture 11 00987 i012]() : Proteobacteria_Gammaproteobacteria_Chromatiales;
: Proteobacteria_Gammaproteobacteria_Chromatiales; ![Agriculture 11 00987 i004]() : Proteobacteria_Gammaproteobacteria_Pseudomonadales;
: Proteobacteria_Gammaproteobacteria_Pseudomonadales; ![Agriculture 11 00987 i013]() : Proteobacteria_Gammaproteobacteria_Thiotrichales;
: Proteobacteria_Gammaproteobacteria_Thiotrichales; ![Agriculture 11 00987 i014]() : SBR1093_VHS-BS-50;
: SBR1093_VHS-BS-50; ![Agriculture 11 00987 i015]() : Verrucomicrobia_Pedosphaerae_Pedosphaerales;
: Verrucomicrobia_Pedosphaerae_Pedosphaerales; ![Agriculture 11 00987 i016]() : Verrucomicrobia_Spartobacteria_Chthoniobacterales;
: Verrucomicrobia_Spartobacteria_Chthoniobacterales; ![Agriculture 11 00987 i006]() : Unknown. Healthy plant: OK; diseased plant: NG.
: Unknown. Healthy plant: OK; diseased plant: NG.
 : Verrucomicrobia;
: Verrucomicrobia;  : Proteobacteria;
: Proteobacteria;  : Actinobacteria;
: Actinobacteria;  : SBR1093;
: SBR1093;  : Planctomycetes;
: Planctomycetes;  : unknown. (B) : Actinobacteria_Actinobacteria_Actinomycetales; : Planctomycetes_OM190_agg27; : Proteobacteria;
: unknown. (B) : Actinobacteria_Actinobacteria_Actinomycetales; : Planctomycetes_OM190_agg27; : Proteobacteria;  : Proteobacteria_Deltaproteobacteria;
: Proteobacteria_Deltaproteobacteria;  : Proteobacteria_Deltaproteobacteria_Desulfuromonadales; : Proteobacteria_Deltaproteobacteria_Desulfovibrionales;
: Proteobacteria_Deltaproteobacteria_Desulfuromonadales; : Proteobacteria_Deltaproteobacteria_Desulfovibrionales;  : Proteobacteria_Deltaproteobacteria_NB1-j;
: Proteobacteria_Deltaproteobacteria_NB1-j;  : Proteobacteria_Epsilonproteobacteria_Campylobacteriales;
: Proteobacteria_Epsilonproteobacteria_Campylobacteriales;  : Proteobacteria_Gammaproteobacteria;
: Proteobacteria_Gammaproteobacteria;  : Proteobacteria_Gammaproteobacteria_Chromatiales; : Proteobacteria_Gammaproteobacteria_Pseudomonadales;
: Proteobacteria_Gammaproteobacteria_Chromatiales; : Proteobacteria_Gammaproteobacteria_Pseudomonadales;  : Proteobacteria_Gammaproteobacteria_Thiotrichales;
: Proteobacteria_Gammaproteobacteria_Thiotrichales;  : SBR1093_VHS-BS-50;
: SBR1093_VHS-BS-50;  : Verrucomicrobia_Pedosphaerae_Pedosphaerales;
: Verrucomicrobia_Pedosphaerae_Pedosphaerales;  : Verrucomicrobia_Spartobacteria_Chthoniobacterales; : Unknown. Healthy plant: OK; diseased plant: NG.
: Verrucomicrobia_Spartobacteria_Chthoniobacterales; : Unknown. Healthy plant: OK; diseased plant: NG.
Figure 4.
Stacked bar plots of relative abundance of bacterial (A) phyla and (B) order. (A) ![Agriculture 11 00987 i001]() : Verrucomicrobia;
: Verrucomicrobia; ![Agriculture 11 00987 i002]() : Proteobacteria;
: Proteobacteria; ![Agriculture 11 00987 i003]() : Actinobacteria;
: Actinobacteria; ![Agriculture 11 00987 i004]() : SBR1093;
: SBR1093; ![Agriculture 11 00987 i005]() : Planctomycetes;
: Planctomycetes; ![Agriculture 11 00987 i006]() : unknown. (B)
: unknown. (B) ![Agriculture 11 00987 i003]() : Actinobacteria_Actinobacteria_Actinomycetales;
: Actinobacteria_Actinobacteria_Actinomycetales; ![Agriculture 11 00987 i005]() : Planctomycetes_OM190_agg27;
: Planctomycetes_OM190_agg27; ![Agriculture 11 00987 i002]() : Proteobacteria;
: Proteobacteria; ![Agriculture 11 00987 i007]() : Proteobacteria_Deltaproteobacteria;
: Proteobacteria_Deltaproteobacteria; ![Agriculture 11 00987 i008]() : Proteobacteria_Deltaproteobacteria_Desulfuromonadales;
: Proteobacteria_Deltaproteobacteria_Desulfuromonadales; ![Agriculture 11 00987 i001]() : Proteobacteria_Deltaproteobacteria_Desulfovibrionales;
: Proteobacteria_Deltaproteobacteria_Desulfovibrionales; ![Agriculture 11 00987 i009]() : Proteobacteria_Deltaproteobacteria_NB1-j;
: Proteobacteria_Deltaproteobacteria_NB1-j; ![Agriculture 11 00987 i010]() : Proteobacteria_Epsilonproteobacteria_Campylobacteriales;
: Proteobacteria_Epsilonproteobacteria_Campylobacteriales; ![Agriculture 11 00987 i011]() : Proteobacteria_Gammaproteobacteria;
: Proteobacteria_Gammaproteobacteria; ![Agriculture 11 00987 i012]() : Proteobacteria_Gammaproteobacteria_Chromatiales;
: Proteobacteria_Gammaproteobacteria_Chromatiales; ![Agriculture 11 00987 i004]() : Proteobacteria_Gammaproteobacteria_Pseudomonadales;
: Proteobacteria_Gammaproteobacteria_Pseudomonadales; ![Agriculture 11 00987 i013]() : Proteobacteria_Gammaproteobacteria_Thiotrichales;
: Proteobacteria_Gammaproteobacteria_Thiotrichales; ![Agriculture 11 00987 i014]() : SBR1093_VHS-BS-50;
: SBR1093_VHS-BS-50; ![Agriculture 11 00987 i015]() : Verrucomicrobia_Pedosphaerae_Pedosphaerales;
: Verrucomicrobia_Pedosphaerae_Pedosphaerales; ![Agriculture 11 00987 i016]() : Verrucomicrobia_Spartobacteria_Chthoniobacterales;
: Verrucomicrobia_Spartobacteria_Chthoniobacterales; ![Agriculture 11 00987 i006]() : Unknown. Healthy plant: OK; diseased plant: NG.
: Unknown. Healthy plant: OK; diseased plant: NG.
: Verrucomicrobia; : Proteobacteria; : Actinobacteria; : SBR1093; : Planctomycetes; : unknown. (B) : Actinobacteria_Actinobacteria_Actinomycetales; : Planctomycetes_OM190_agg27; : Proteobacteria; : Proteobacteria_Deltaproteobacteria; : Proteobacteria_Deltaproteobacteria_Desulfuromonadales; : Proteobacteria_Deltaproteobacteria_Desulfovibrionales; : Proteobacteria_Deltaproteobacteria_NB1-j; : Proteobacteria_Epsilonproteobacteria_Campylobacteriales; : Proteobacteria_Gammaproteobacteria; : Proteobacteria_Gammaproteobacteria_Chromatiales; : Proteobacteria_Gammaproteobacteria_Pseudomonadales; : Proteobacteria_Gammaproteobacteria_Thiotrichales; : SBR1093_VHS-BS-50; : Verrucomicrobia_Pedosphaerae_Pedosphaerales; : Verrucomicrobia_Spartobacteria_Chthoniobacterales; : Unknown. Healthy plant: OK; diseased plant: NG.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, T.-C.; Peng, H.-M.; Wollney, S.; Shen, C.-H. Rhizosphere Microbiome Regulates the Growth of Mustard under Organic Greenhouse Cultivation. Agriculture 2021, 11, 987. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11100987
AMA Style
Liu T-C, Peng H-M, Wollney S, Shen C-H. Rhizosphere Microbiome Regulates the Growth of Mustard under Organic Greenhouse Cultivation. Agriculture. 2021; 11(10):987. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11100987
Chicago/Turabian StyleLiu, Ting-Chung, Hui-Mei Peng, Seth Wollney, and Chang-Hui Shen. 2021. "Rhizosphere Microbiome Regulates the Growth of Mustard under Organic Greenhouse Cultivation" Agriculture 11, no. 10: 987. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11100987
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.