
Effect of Plant-Growth-Promoting Fungi on Eggplant (Solanum melongena L.) in New Reclamation Land

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sample Collection and Fungal Isolation
2.2. Determination of Phosphate-Solubilizing Ability
2.3. Determination of Siderophore Production
2.4. Indole Acetic Acid (IAA) Detection
2.5. Morphological Identification
2.6. Phylogenetic Analysis
2.7. Effect of Isolates on Plant Growth Promotion in New Reclamation Land
2.8. Statistical Analysis
3. Results
3.1. Fungal Isolation and Screening of Phosphate-Solubilizing Ability
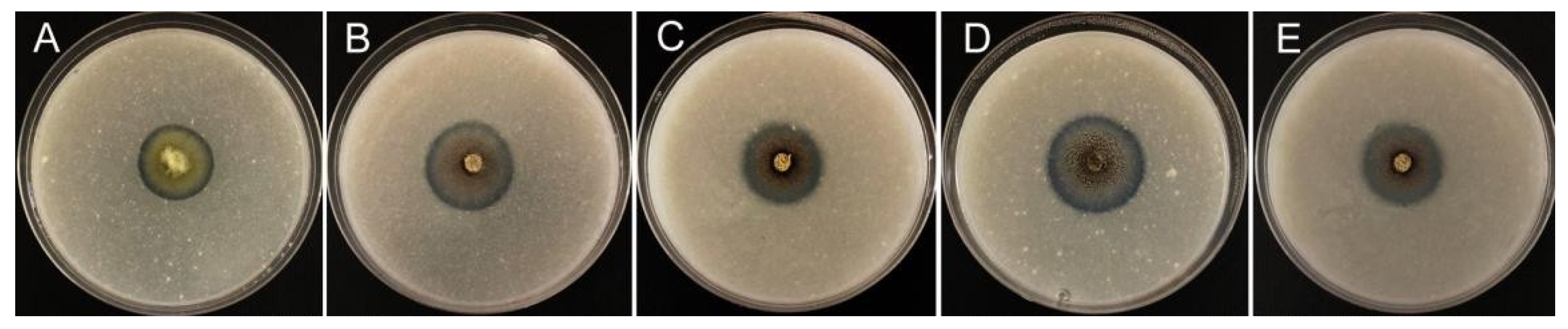
3.2. Siderophore Production
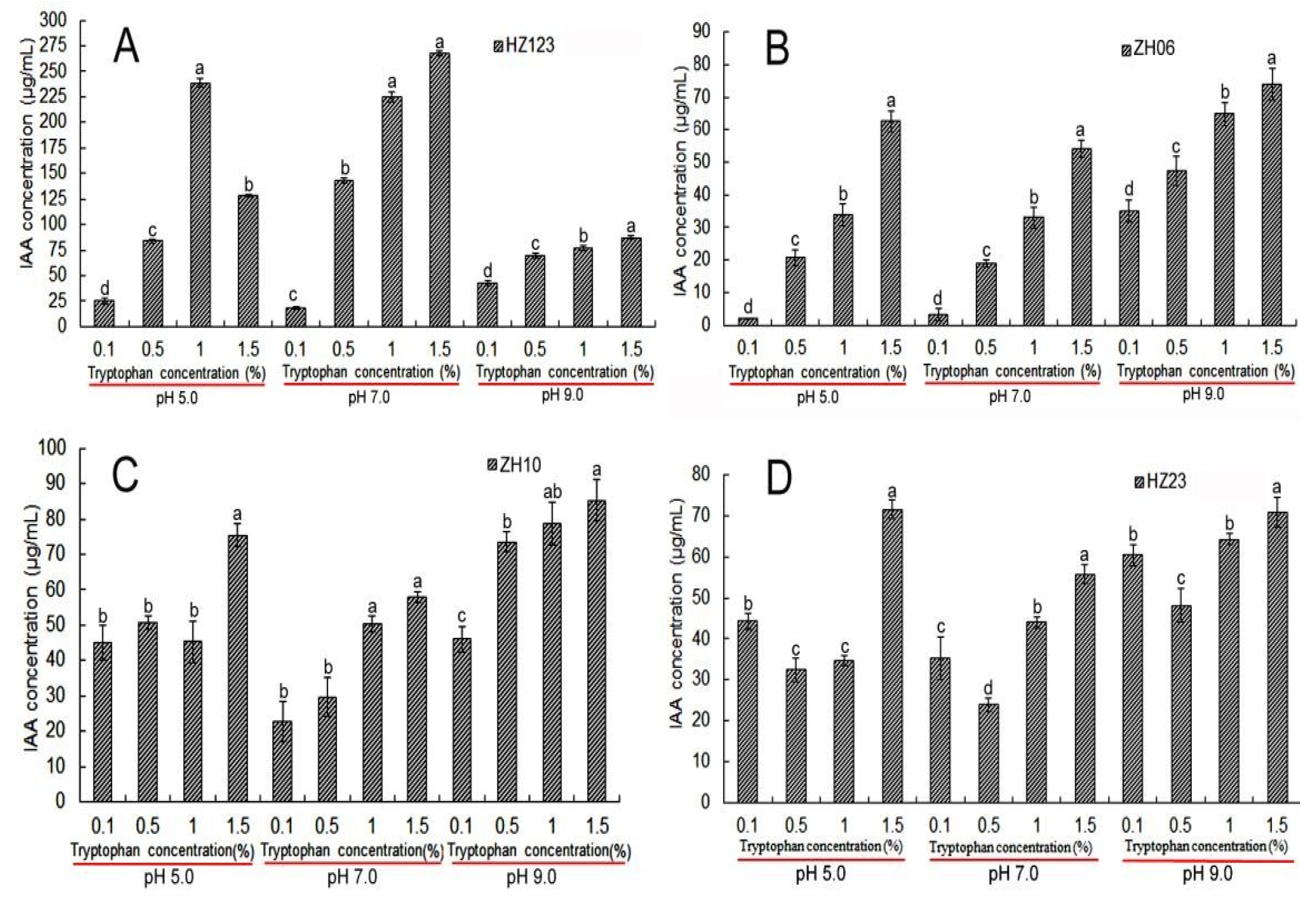
3.3. Indole Acetic Acid (IAA) Detection
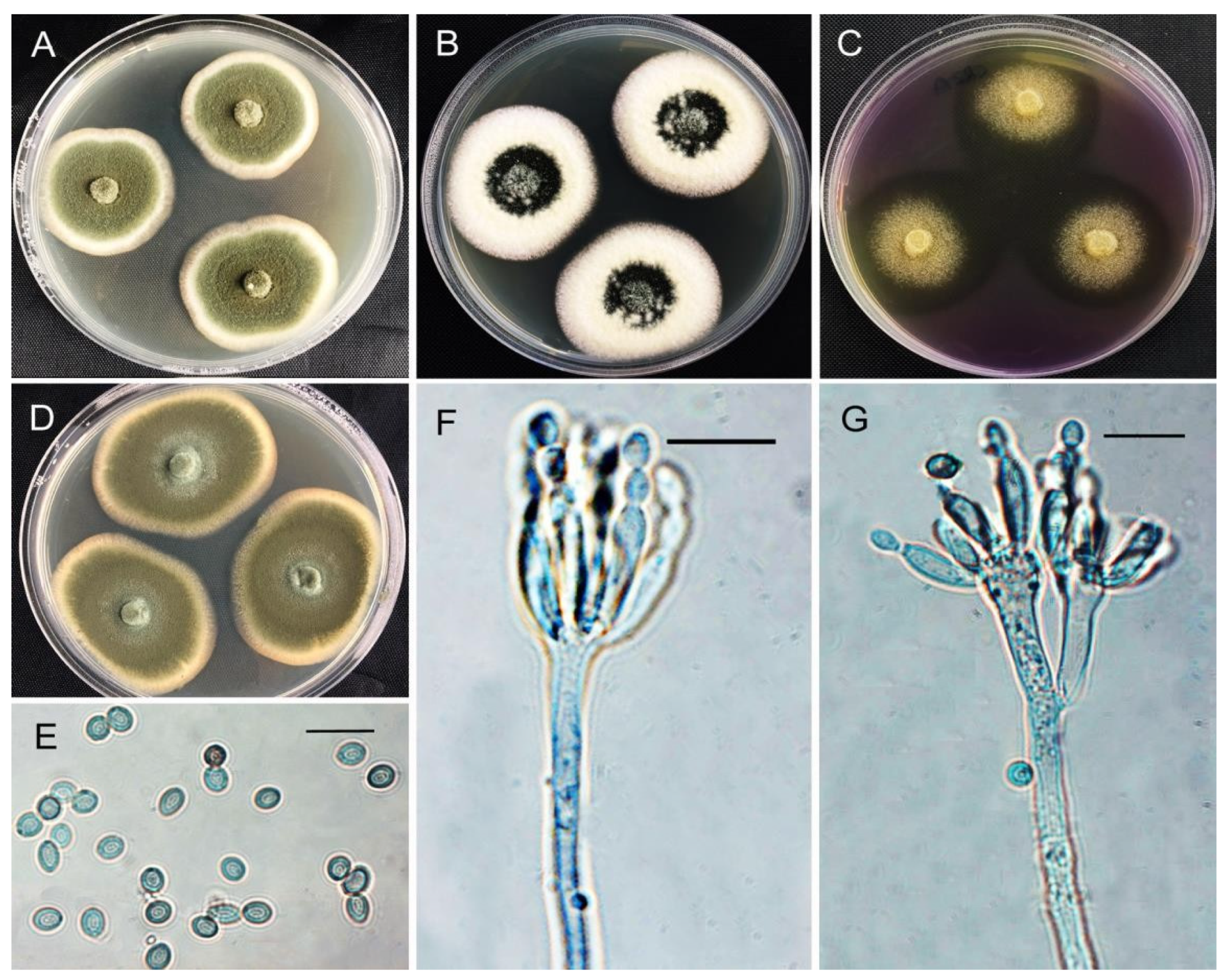
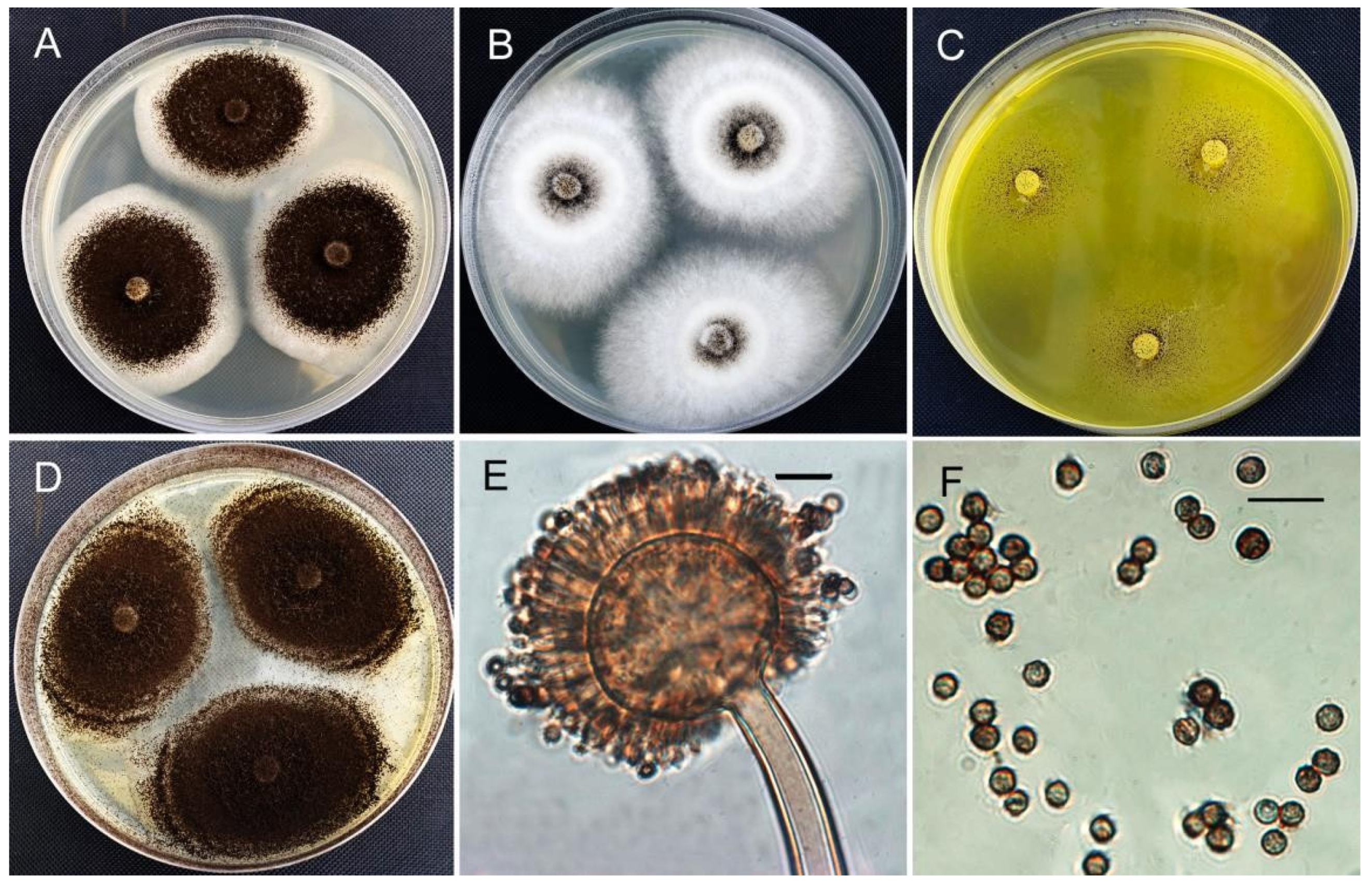
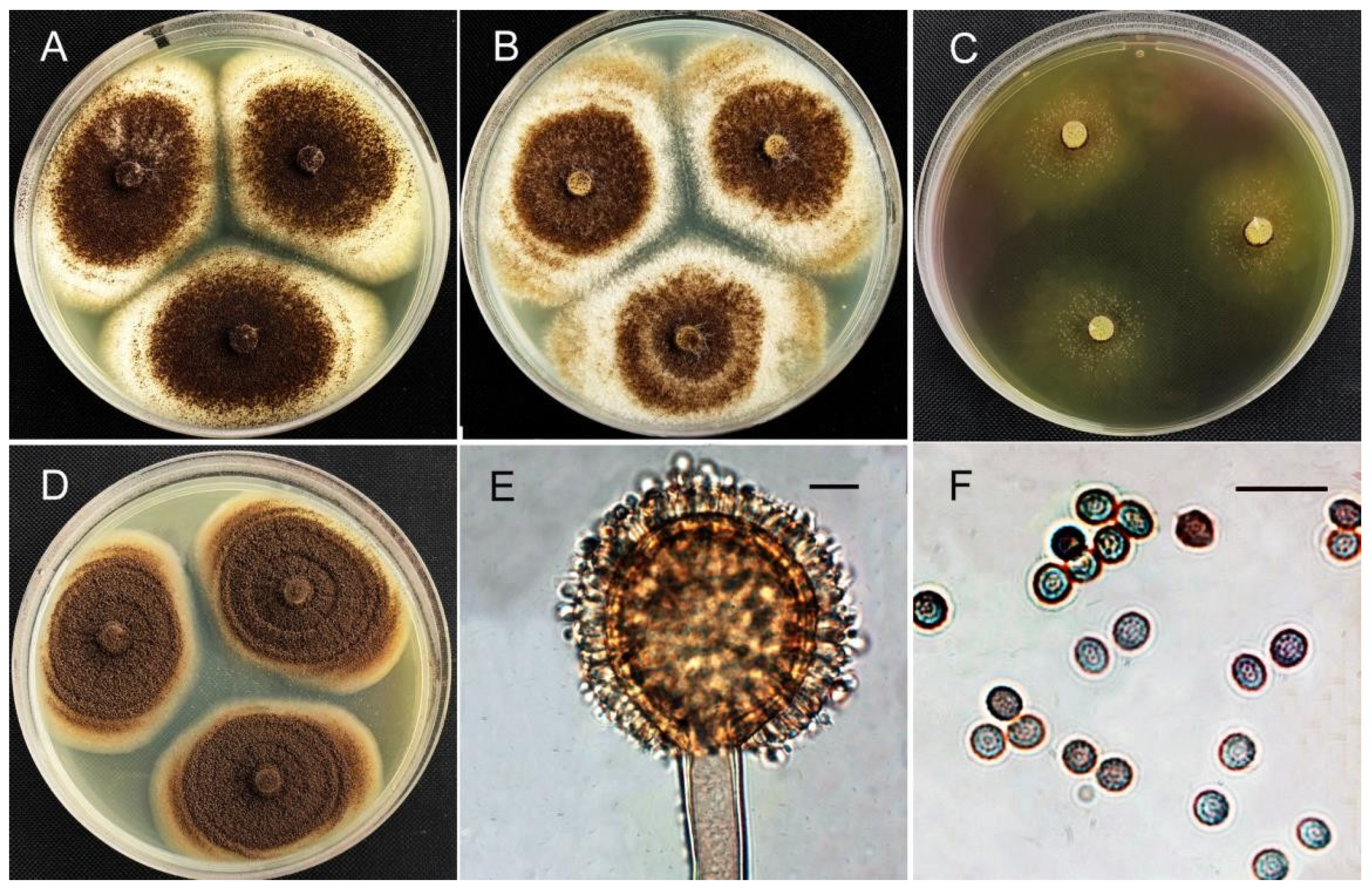
3.4. Morphological Identification
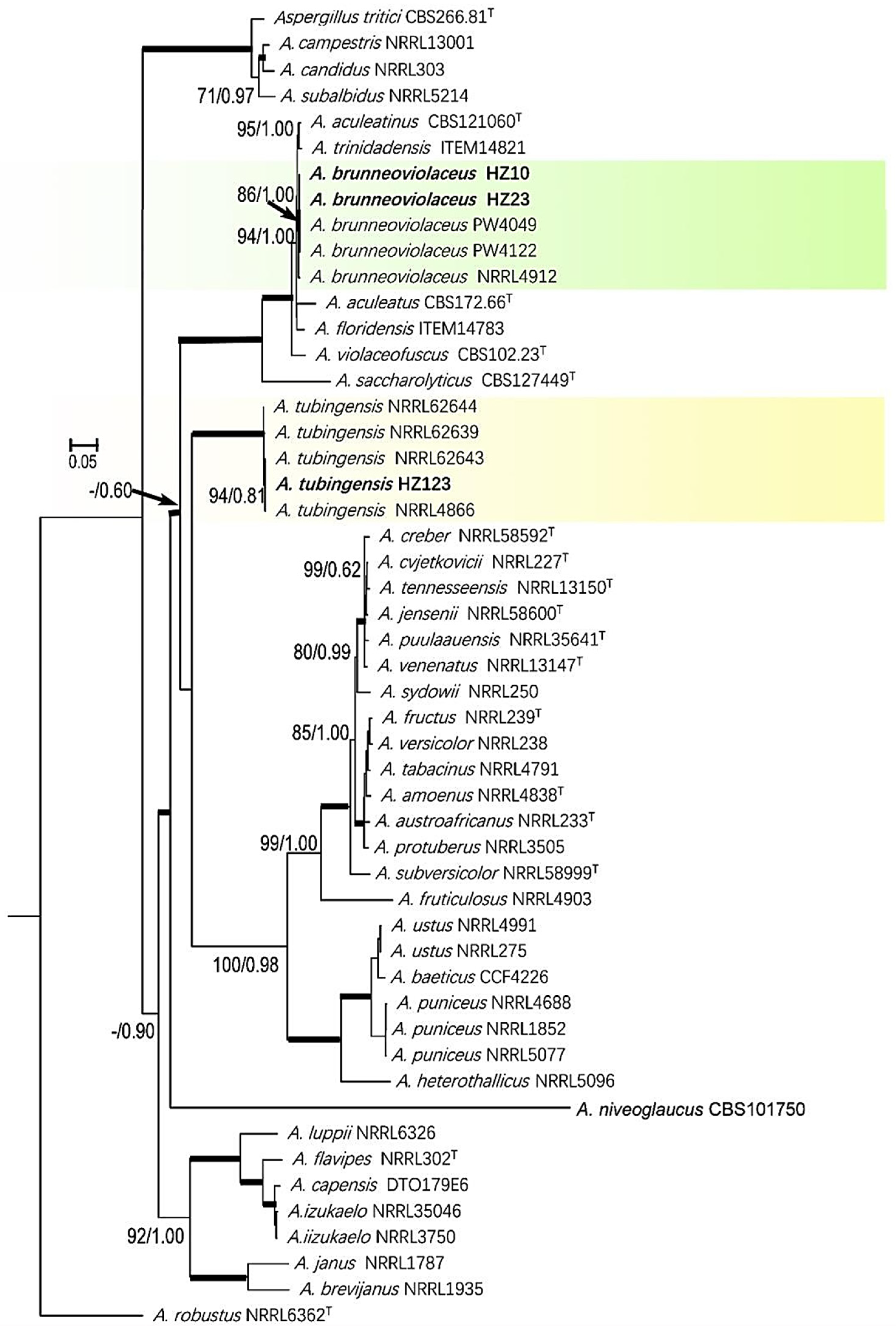
3.5. Phylogenic Analysis of Sequence Data
3.6. Effect of Fungal Isolates on Plant Growth Promotion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, X.Q.; Su, Y.; Ahmed, T.; Ren, H.Y.; Javed, M.R.; Yao, Y.L.; An, Q.L.; Yan, J.L.; Li, B. Effects of different organic fertilizers on improving soil from newly reclaimed land to crop soil. Agriculture 2021, 11, 560. [Google Scholar] [CrossRef]
- Larney, F.J.; Angers, D.A. The role of organic amendments in soil reclamation: A review. Can. J. Soil Sci. 2012, 92, 19–38. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- Souza, R.D.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef]
- Zhang, F.; Huo, Y.; Cobb, A.B.; Luo, G.; Zhou, J.; Yang, G.; Wilson, G.W.T.; Zhang, Y. Trichoderma biofertilizer links to altered soil chemistry, altered microbial communities, and improved grassland biomass. Front. Microbiol. 2018, 9, 848. [Google Scholar] [CrossRef]
- Li, Z.; Bai, T.; Dai, L.; Wang, F.; Tao, J.; Meng, S.; Hu, Y.; Wang, S.; Hu, S. A study of organic acid production in contrasts between two phosphate solubilizing fungi: Penicillium oxalicum and Aspergillus niger. Sci. Rep. 2016, 6, 25313. [Google Scholar] [CrossRef] [PubMed]
- Narsian, V.; Patel, H.H. Aspergillus aculeatus as a rock phosphate solubilizer. Soil Biol. Biochem. 2000, 32, 559–565. [Google Scholar] [CrossRef]
- Haas, H. Fungal siderophore metabolism with a focus on Aspergillus fumigates. Nat. Prod. Rep. 2014, 31, 1266–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, N.V.; Rajam, K.S.; Rani, M.E. Plant growth promotion efficacy of indole acetic acid (iaa) produced by a mangrove associated fungi-Trichoderma viride vkf3. Int. J. Curr. Microbiol. 2017, 6, 2692–2701. [Google Scholar] [CrossRef]
- Lee, S.; Yap, M.; Behringer, G.; Hung, R.; Bennett, J.W. Volatile organic compounds emitted by Trichoderma species mediate plant growth. Fungal Biol. Biotechnol. 2016, 3, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.M.; Sultana, F. Application and mechanisms of plant growth promoting fungi (PGPF) for phytostimulation. In Organic Agriculture; IntechOpen: London, UK, 2020; pp. 1–31. [Google Scholar]
- Mehta, S.; Nautiyal, C.S. An efficient method for qualitative screening of phosphate solubilizing bacteria. Curr. Microbiol. 2001, 43, 51–55. [Google Scholar] [CrossRef]
- Tang, A.; Haruna, A.O.; Majid, N.M.A.; Jalloh, M.B. Potential PGPR properties of cellulolytic, nitrogen-fixing, phosphate-solubilizing bacteria in rehabilitated tropical forest soil. Microorganisms 2020, 8, 442. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.X.; Zhang, M.C.; Loh, B.; Leptihn, S.; Ahmed, T.; Li, B. A novel NRPS cluster, acquired by horizontal gene transfer from algae, regulates siderophore iron metabolism in Burkholderia seminalis R456. Int. J. Biol. Macromol. 2021, 182, 838–848. [Google Scholar] [CrossRef] [PubMed]
- Murugappan, R.M.; Aravinth, A.; Karthikeyan, M. Chemical and structural characterization of hydroxamate siderophore produced by marine Vibrio harveyi. J. Ind. Microbiol. Biotechnol. 2011, 38, 265–273. [Google Scholar] [CrossRef]
- Khamna, S.; Yokota, A.; Lumyong, S. Actinomycetes isolated from medicinal plant rhizosphere soils: Diversity and screening of antifungal compounds, indole-3-acetic acid and siderophore production. World J. Microbiol. Biotechnol. 2009, 25, 649–655. [Google Scholar] [CrossRef]
- Raut, V.; Shaikh, I.; Naphade, B.; Prashar, K.; Adhapure, N. Plant growth promotion using microbial IAA producers in conjunction with azolla: A novel approach. Chem. Biol. Technol. 2017, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Z.; Li, M.J. A new species of Bipolaris from the halophyte Sesuvium portulacastrum in Guangdong Province, China. Mycotaxon 2009, 109, 289–300. [Google Scholar] [CrossRef]
- Tomaha, A.A.; Abd Alamera, I.S.; Li, B.; Zhang, J. A new species of Trichoderma and gliotoxin role: A new observation in enhancing biocontrol potential of T. virens against Phytophthora capsici on chili pepper. Biol. Control. 2020, 145, 104261. [Google Scholar] [CrossRef]
- Nasri, T.; Hedayati, M.T.; Abastabar, M.; Pasqualotto, A.C.; Armaki, M.T.; Hoseinnejad, A.; Nabili, M. PCR-RFLP on beta-tubulin gene for rapid identification of the most clinically important species of Aspergillus. J. Microbiol. Methods 2015, 117, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Haituk, S.; Suwannarach, N.; Hongsanan, S.; Senwanna, C.; Cheewangkoon, R. New genus of epiphytic sooty mold: Alloscorias syngonii (Readerielliopsidaceae) from Thailand. Phytotaxa 2021, 507, 271–282. [Google Scholar] [CrossRef]
- Hong, S.B.; Go, S.J.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Aspergillusfumigatus and related species. Mycologia 2005, 97, 1316–1329. [Google Scholar] [CrossRef]
- Tippmann, H.F. Analysis for free: Comparing programs for sequence analysis. Brief. Bioinform. 2004, 5, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Munkhbata, B.; Tounaia, K.; Manoa, S.; Andoa, H.; Oyungerelb, G.; Chaec, G.T.; Hand, H.; Jiae, G.J.; Tokunagaf, K.; et al. Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis. Gene 2005, 346, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Tsang, C.C.; Tang, J.Y.M.; Lau, S.K.P.; Woo, P.C.Y. Taxonomy and evolution of Aspergillus, Penicillium and Talaromyces in the omics era—Past, present and future. Comput. Struct. Biotechnol. 2018, 16, 197–210. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsube, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Kubátová, A.; Hujslová, M.; Frisvad, J.C.; Chudíčková, M.; Kolařík, M. Taxonomic revision of the biotechnologically important species Penicillium oxalicum with the description of two new species from acidic and saline soils. Mycol. Prog. 2019, 18, 215–228. [Google Scholar] [CrossRef]
- Horn, B.W.; Olarte, R.A.; Peterson, S.W.; Carbone, I. Sexual reproduction in Aspergillus tubingensis from section Nigri. Mycologia 2013, 105, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Jurjević, Z.; Peterson, S.W.; Stea, G.; Solfrizzo, M.; Varga, J.; Hubka, V.; Perrone, G. Two novel species of aspergillus section nigri from indoor air. IMA Fungus 2012, 3, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Ibrahim, M.; Cui, Z.Q.; Xie, G.L.; Jin, G.L.; Kube, M.; Li, B.; Zhou, X.P. Multi-omics analysis of niche specificity provides new insights into ecological adaptation in bacteria. ISME J. 2016, 10, 2072–2075. [Google Scholar] [CrossRef] [Green Version]
- Araujo, V.C.; Rossati, K.F.; Xavier, L.V.; de Oliveira, V.A.; Carmo, G.J.D.; de Assis, G.A.; Mendes, G.D. Enhanced growth in nursery of coffee seedlings inoculated with the rhizosphere fungus Aspergillus niger for field transplantation. Rhizosphere 2020, 15, 100236. [Google Scholar] [CrossRef]
- Javed, A.; Shah, A.H.; Hussain, A.; Shinwari, Z.K.; Khan, S.A.; Khan, W.; Jan, S.A. Potential of endophytic fungus Aspergillus terreus as potent plant growth promoter. Pak. J. Bot. 2020, 52, 1083–1086. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Gupta, V.V.S.R.; Harvey, P.R.; Ryder, M.H. The effect of Penicillium fungi on plant growth and phosphorus mobilization in neutral to alkaline soils from southern Australia. Can. J Microbiol. 2007, 53, 106–115. [Google Scholar] [CrossRef]
- Gomez-Munoz, B.; Jensen, L.S.; de Neergaard, A.; Richardson, A.E.; Magid, J. Effects of Penicillium bilaii on maize growth are mediated by available phosphorus. Plant Soil 2018, 431, 159–173. [Google Scholar] [CrossRef]
- Mendes, G.D.O.; Freitas, A.L.M.; Pereira, O.L.; Silva, I.R.; Vassilev, N.B.; Costa, M.D. Mechanisms of phosphate solubilization by fungal isolates when exposed to different P sources. Ann. Microbiol. 2014, 64, 239–249. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef] [Green Version]
- Ikram, M.; Ali, N.; Jan, G.; Jan, F.G.; Rahman, I.U.; Iqbal, A.; Hamayun, M. IAA producing fungal endophyte Penicillium roqueforti Thom., enhances stress tolerance and nutrients uptake in wheatplants grown on heavy metal contaminated soils. PLoS ONE. 2018, 13, e0208150. [Google Scholar] [CrossRef]
- Mehmood, A.; Hussain, A.; Irshad, M.; Hamayun, M.; Khan, N. In vitro production of IAA by endophytic fungus Aspergillus awamori and its growth promoting activities in zea mays. Symbiosis 2019, 77, 225–235. [Google Scholar] [CrossRef]
- Chowdappa, S.; Jagannath, S.; Konappa, N.; Udayashankar, A.C.; Jogaiah, S. Detection and characterization of antibacterial siderophores secreted by endophytic fungi from Cymbidium aloifolium. Biomolecules 2020, 10, 1412. [Google Scholar] [CrossRef]
- Osman, Y.; Gebreil, A.; Mowafy, A.M.; Anan, T.I.; Hamed, S.M. Characterization of Aspergillus niger siderophore that mediates bioleaching of rare earth elements from phosphorites. World J. Microbiol. Biotechnol. 2019, 35, 0959–3993. [Google Scholar] [CrossRef] [PubMed]
- Vyas, P.; Gulati, A. Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbol. 2009, 9, 174. [Google Scholar] [CrossRef] [Green Version]
- Rasheed, A.; Eugene, C.T.; Annick, B.; Damase, P.K. Relationship between plant growth and organic acid exudates from ectomycorrhizal and non-ectomycorrhizal Pinus patula. S. Afr. J. Plant Soil 2015, 32, 183–188. [Google Scholar]
- Krishna, P.; Reddy, M.S.; Patnaik, S.K. Aspergillus tubingensis reduces the pH of the bauxite residue (red mud) amended soils. Water Air Soil Pollut. 2005, 167, 201–209. [Google Scholar] [CrossRef]
- Samson, R.A.; Noonim, P.; Meijer, M.; Houbraken, J.C.; Frisvad, J.C.; Varga, J. Diagnostic tools to identify black aspergilla. Stud. Mycol. 2007, 59, 129–145. [Google Scholar] [CrossRef]
- Singh, S.K.; Reddy, K.R. Regulation of photosynthesis, fluorescence, stomatal conductance and water-use efficiency of cowpea (Vigna unguiculata [L.] Walp.) under drought. J. Photochem. Photobiol. B Biol. 2011, 105, 40–50. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Identity | Colony Diameter (mm) | Acid Production | |||
|---|---|---|---|---|---|---|
| CYA (25 °C) | MEA (25 °C) | CREA (25 °C) | CYA (37 °C) | |||
| HZ06 | P. oxalicum | 18–21 | 13–18 | 12–15 | 22–37 | + |
| HZ123 | A. tubingensis | 52–60 | 42–48 | 10–14 | 52–54 | +++ |
| HZ10, 23 | A. brunneoviolaceus | 60–62 | 46–50 | 32–33 | 22–29 | ++ |
| Strains | SL (cm) | GPE (%) | RL (cm) | GPE (%) | SFW (g) | GPE (%) | SDW (g) | GPE (%) | RFW (g) | GPE% | RDW (g) | GPE (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. oxalicum HZ06 | 14.07 ± 0.76 | 22.35 a | 27.83 ± 0.98 | 0.83 b | 9.29 ± 0.96 | 47.69 a | 1.88 ± 0.29 | 39.26 a | 3.81 ± 0.46 | 28.72 ab | 0.65 ± 0.08 | 41.30 a |
| A. brunneoviolaceus HZ23 | 12.95 ± 0.96 | 12.61 b | 28.15 ± 1.26 | 1.99 b | 8.40 ± 0.80 | 33.55 b | 1.73 ± 0.27 | 28.15 b | 3.78 ± 0.41 | 27.70 b | 0.59 ± 0.07 | 28.26 b |
| A. brunneoviolaceus HZ10 | 12.26 ± 0. 89 | 6.61 c | 29.84 ± 1.54 | 8.12 a | 7.58 ± 0.62 | 20.51 c | 1.67 ± 0.26 | 23.70 b | 4.02 ± 0.68 | 35.81 a | 0.66 ± 0.10 | 43.48 a |
| A. tubingensis HZ123 | 12.50 ± 0.89 | 8.70 bc | 30.10 ± 2.58 | 9.06 a | 6.24 ± 1.74 | −0.79 d | 1.32 ± 0.14 | −2.22 c | 3.79 ± 0.40 | 28.04 b | 0.51 ± 0.08 | 10.87 c |
| Control | 11.50 ± 0.53 | — | 27.59 ± 1.89 | — | 6.29 ± 0.84 | — | 1.35 ± 0.26 | — | 2.96 ± 0.37 | — | 0.46 ± 0.08 | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Li, D.; Yan, J.; Zhang, Y.; Wang, H.; Zhang, J.; Ahmed, T.; Li, B. Effect of Plant-Growth-Promoting Fungi on Eggplant (Solanum melongena L.) in New Reclamation Land. Agriculture 2021, 11, 1036. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11111036
Li X, Li D, Yan J, Zhang Y, Wang H, Zhang J, Ahmed T, Li B. Effect of Plant-Growth-Promoting Fungi on Eggplant (Solanum melongena L.) in New Reclamation Land. Agriculture. 2021; 11(11):1036. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11111036
Chicago/Turabian StyleLi, Xuqing, Dingyi Li, Jianli Yan, Ya Zhang, Hong Wang, Jingze Zhang, Temoor Ahmed, and Bin Li. 2021. "Effect of Plant-Growth-Promoting Fungi on Eggplant (Solanum melongena L.) in New Reclamation Land" Agriculture 11, no. 11: 1036. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11111036