1. Introduction
Poultry production holds a respectful position to meet the ever-increasing demand for edible protein. Continuous improvement in the genetic strains in meat-type chicken has reduced the marketable age. The extensively competitive environment of the livestock industry puts pressure to minimize the cost of production. Feed formulation and additives have greatly improved the production performances and feed efficiency of modern broiler strains [
1]. Nutritional programming is among the most favorable methods to produce maximum weight with minimal expenditure. Feed cost is the major expenditure for rearing meat-type chicken [
2]. Precision in nutrition may help overcome this issue. Supplementation of economical products having sufficient availability and arising from unused stuff can be beneficial.
Industrial and agricultural byproducts accumulate as waste and result in a problem for handling. Sawdust and shavings are the most underutilized waste of the wood industry [
3]. They are mostly used as bedding in the poultry industry. There is a considerable possibility to utilize this waste as a fraction in animal feed. Multiple problems can be solved by utilizing these wastes, such as reducing the accumulation of waste and the cost of animal feed. In particular, waste arising from wood sources are rich in fibers. The soluble fibers are suitable for fermentation and short-chain fatty acid (SCFA) production in the gut. The primary concern in utilizing wood waste is its high insoluble fiber content, which negatively influences nutrient digestibility and energy intake by diluting the nutrients [
4]. Inconsistent results have been obtained by using different insoluble fiber sources in chicken feed [
5] that encourage the possibility to explore the enrichment of insoluble fibers in wood waste. Recent development in technology has shown a way to modulate the insoluble fiber contents [
6]. An additional benefit may be the utilization of mechanical techniques, such as heat treatment, for the pretreatment of wood powder to modulate the dietary fiber content. Steam explosion, for instance, could be used to depolymerize hemicellulose in an ecofriendly and economical way [
7]. Furthermore, it may help transform the lignocellulosic biomass structure to be efficiently used as feed additives [
6,
8,
9]. If used properly, dietary fibers can act as a prebiotic product that can modulate growth performance by favoring the growth of beneficial microbes in the intestine. The potential action of prebiotic products is as a substrate for facilitating the fermentation process, enhancing luminal osmosis and modulating immunity by macrophage stimulation and SCFAs production, which may behave as an antagonist to foodborne pathogens [
10].
The primary role of the gastrointestinal tract (GIT) is to digest and assimilate the nutrients of food. The non-digestible fibers present in undigested food are transported to the ceca after passing through the upper GIT track [
11]. The ceca in chickens are the site for the microorganisms containing both favorable and pathogenic microorganisms [
12]. Cecal microbiota has a direct connection with feed efficiency, which may further be correlated with growth performances in chickens [
13]. For instance,
Bacteroides and
Lactobacillus are positively correlated with feed efficiency and growth performances, respectively [
13,
14]. Evaluation of the cecum microbiota will provide a deep insight into how the microbial population is altered due to shredded steam-exploded pine particle (SPP) supplementation and how the growth performance in chickens is modulated.
It is essential to reduce both industrial wastes by their utilization and feed cost in livestock production due to its highly competitive nature. Considering the impact of non-digestible fibers on the chicken gut, the present experiment was conducted to evaluate the effects of increasing the dose of SPP as the source of fiber on broilers’ growth performances and gut microbiota. We hypothesized that SPP can be supplemented to the poultry feed by up to 2% without having any adverse effects.
3. Results
Table 2 presents the effects of different concentrations of dietary SPP supplementation on growth performance parameters from 2 to 4 weeks. ADG, ADFI, and FCR were similar (
p > 0.05) and not affected by dietary SPP supplementation in chickens. However, the final weight was 2.94% higher in the 2% SPP group, and the last week’s FCR was 1.19% lower compared to the control.
Table 3 presents the effect of SPP supplementation on the weight of the digestive organs in the chickens. The weight of the digestive organs, such as the duodenum, jejunum, ileum, liver, and spleen, were similar (
p > 0.05) and not affected by replacing corn with SPP as a prebiotic in the broiler diets.
No variations (
p > 0.05) were seen in the length of the intestinal organs, such as the duodenum, jejunum, and ileum, of chickens fed with an increasing concentration of SPPs in their diets (
Table 4).
No significant effects (
p > 0.05) were observed for the plasma biochemicals, such as glucose, total protein, triglyceride, and total cholesterol, among the treatment groups (
Table 5).
A total of 24 cecum samples from three treatment groups were used for metagenomic analysis. However, three samples (one from each treatment group) were removed at the time of initial screening. After screening for quality control through chimeric reads and 97% sequence similarity, 585,545 sequences were generated that ranged between 18,561 to 41,782. From each sample, 146 to 231 OTUs were generated via clustering analysis with an abundance higher than 0.005%, resulting in a total of 4185 OTUs.
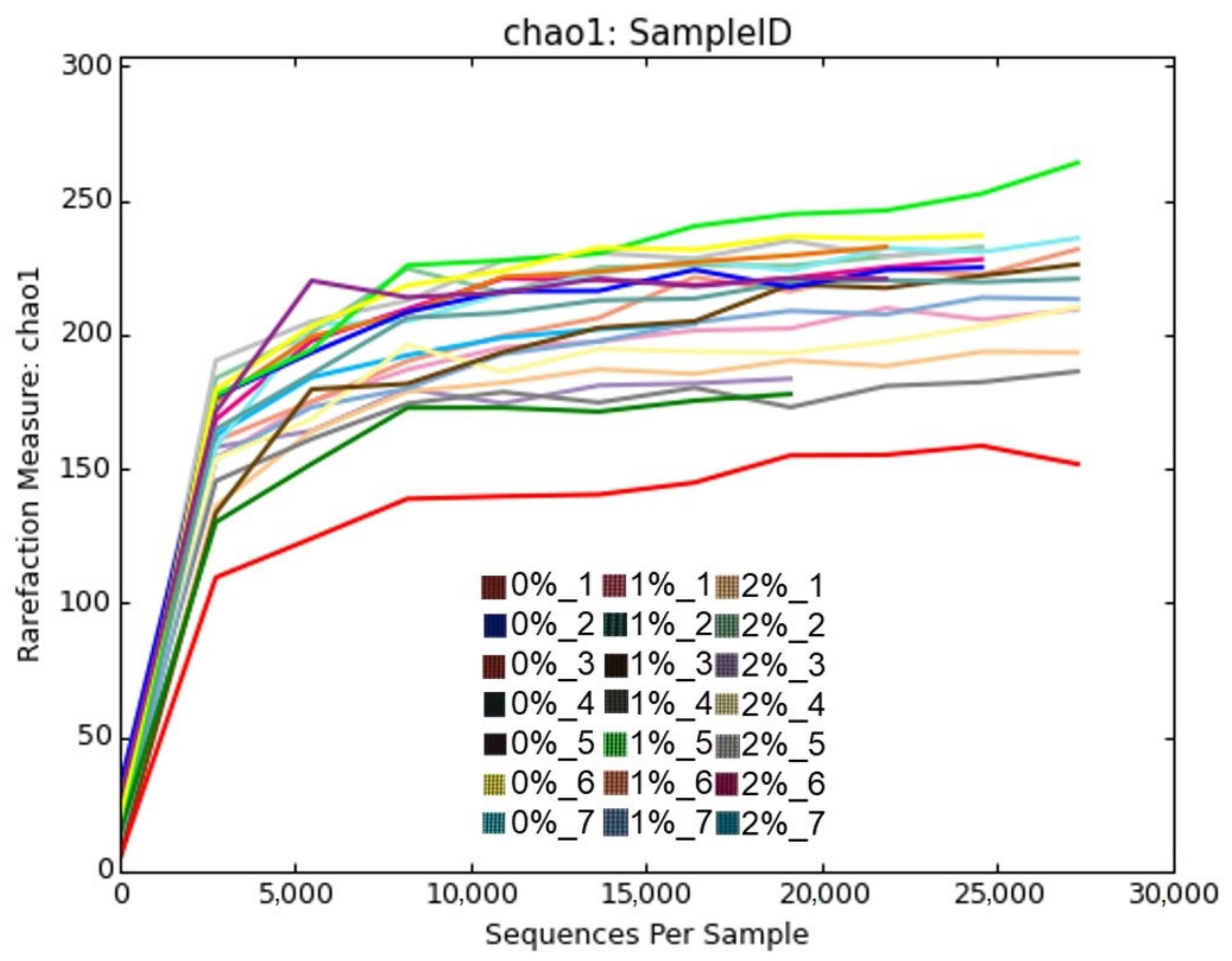
The occurrence of a sufficient number of bacterial communities in the chicken cecum samples was detected by plotting a rarefaction curve for chao1 (
Figure 1). Adequate sampling depth was observed from most of the samples and was represented by flattening of the rarefaction curves towards the right, ultimately reaching a plateau.
For alpha-diversity analysis, community richness and diversity were analyzed using OTUs, Chao1, Shannon, Inverse Simpson, and Goods coverage indices (
Table 6). No significant differences (
p > 0.05) were observed in the alpha-diversity indices among the treatment groups.
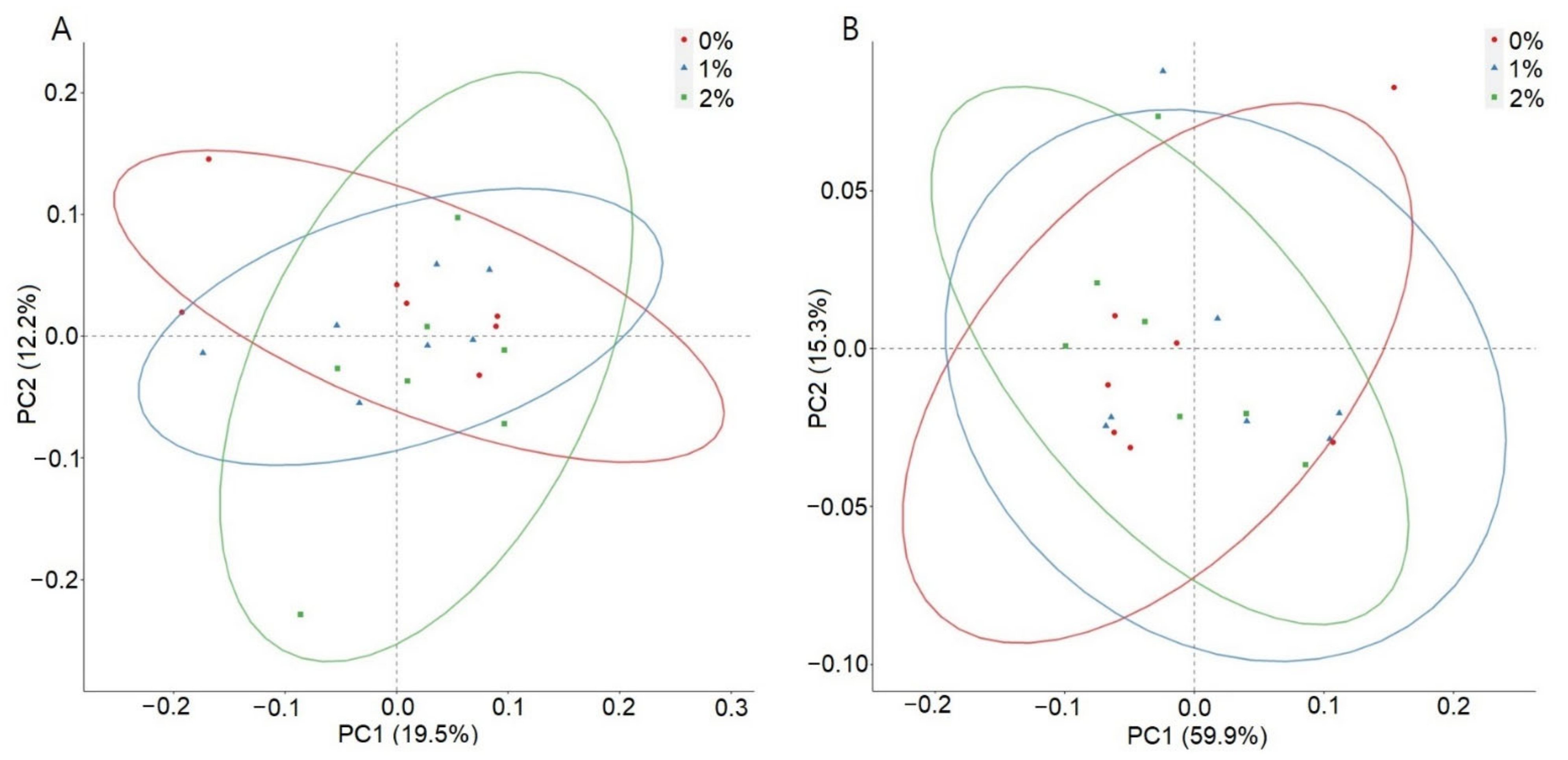
The PCoA based on unweighted unifrac distances were similar (
Figure 2A), whereas weighted unifrac distances showed around 75% cumulative variability based on the two dimensions of PCoA, contributing 59.9% and 15.3% of the inertia, respectively (
Figure 2B).
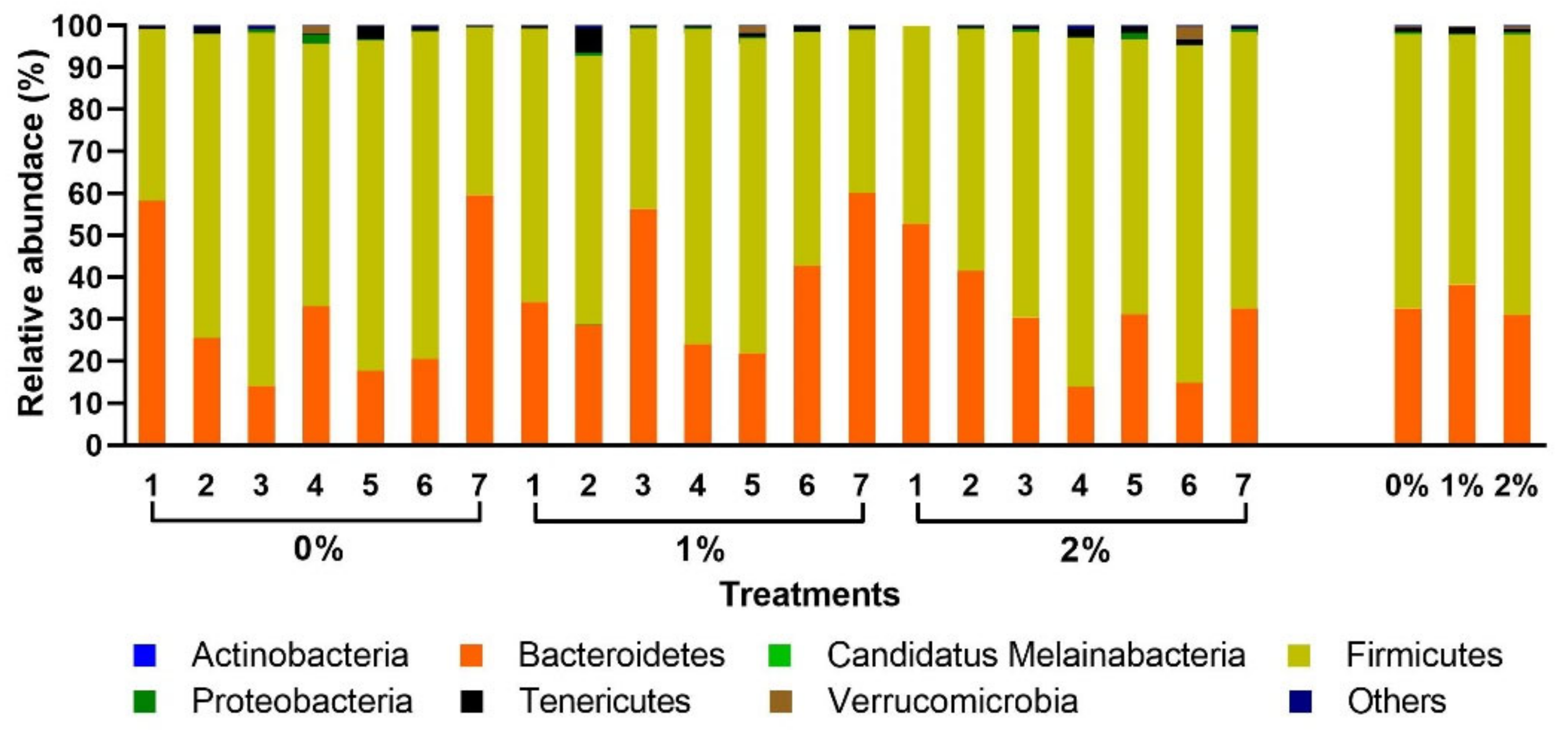
To evaluate the effects of SPP supplementation on the modulation of the cecum microbial community, phylum and genus were analyzed. The bacterial community in the phylum was similar and not significantly affected by the treatment groups.
Firmicutes and
Bacteroidetes were the two most abundant phyla dominating bacterial communities present in the cecum of 28-day-old broiler chickens (
Figure 3).Firmicutes and
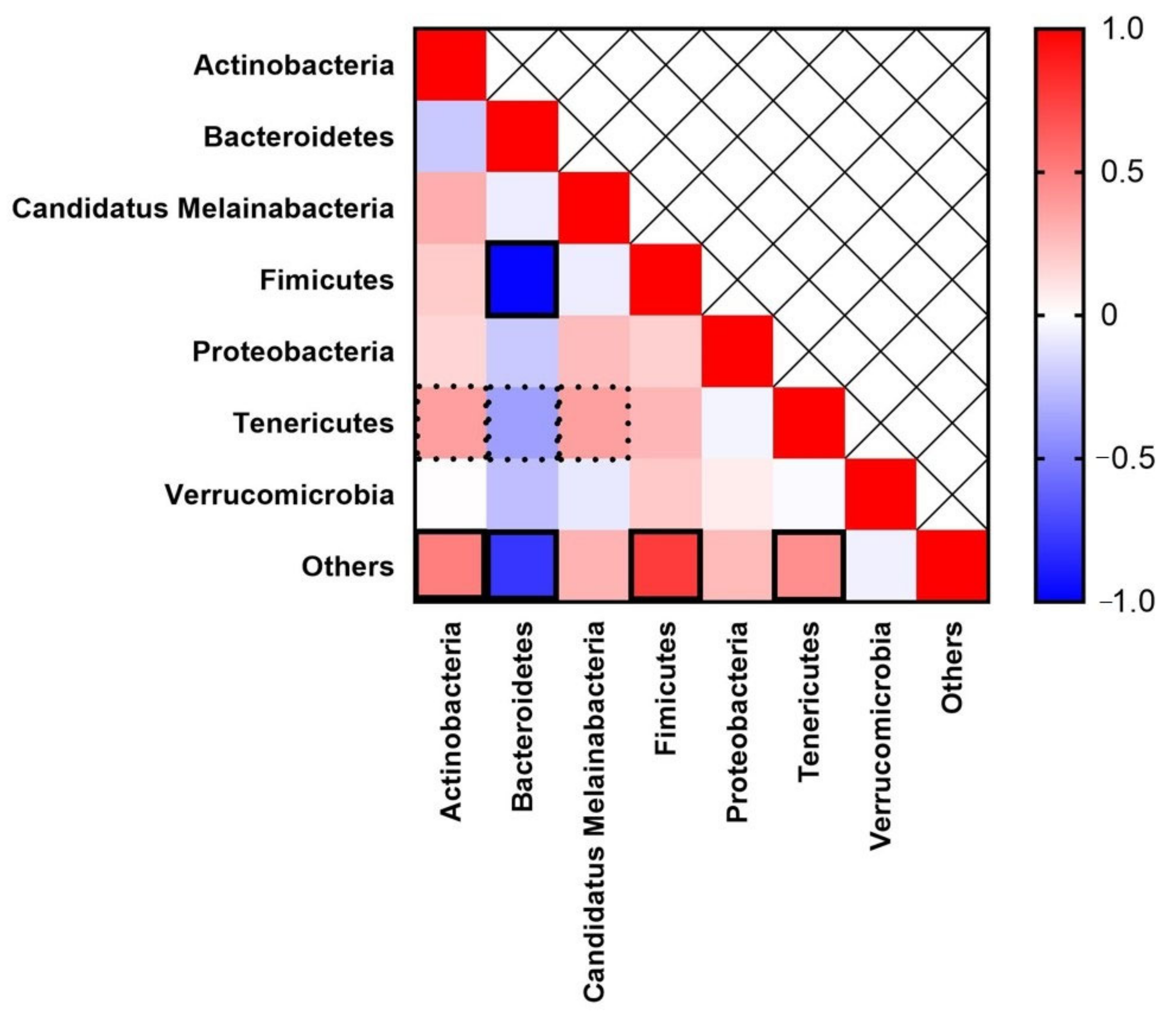
Bacteroidetes were found to be inversely correlated (Spearman 𝑅 = −0.981,
p = 0.001) with each other (
Figure 4).
Bacteroidetes were also found to be inversely correlated with the
Tenericutes (Spearman 𝑅 = −0.378,
p = 0.091) and other (Spearman 𝑅 = −0.791,
p = 0.001) bacteria present in the phylum. However,
Actinobacteria (Spearman 𝑅 = 0.500,
p = 0.021),
Firmicutes (Spearman 𝑅 = 0.764,
p = 0.001), and
Tenericutes (Spearman 𝑅 = 0.442,
p = 0.045) were positively correlated with the other bacteria in the phylum.
Tenericutes was positively correlated with
Actinobacteria (Spearman 𝑅 = 0.376,
p = 0.093) and
Candidatus Melainabacteria (Spearman 𝑅 = −0.369,
p = 0.099).
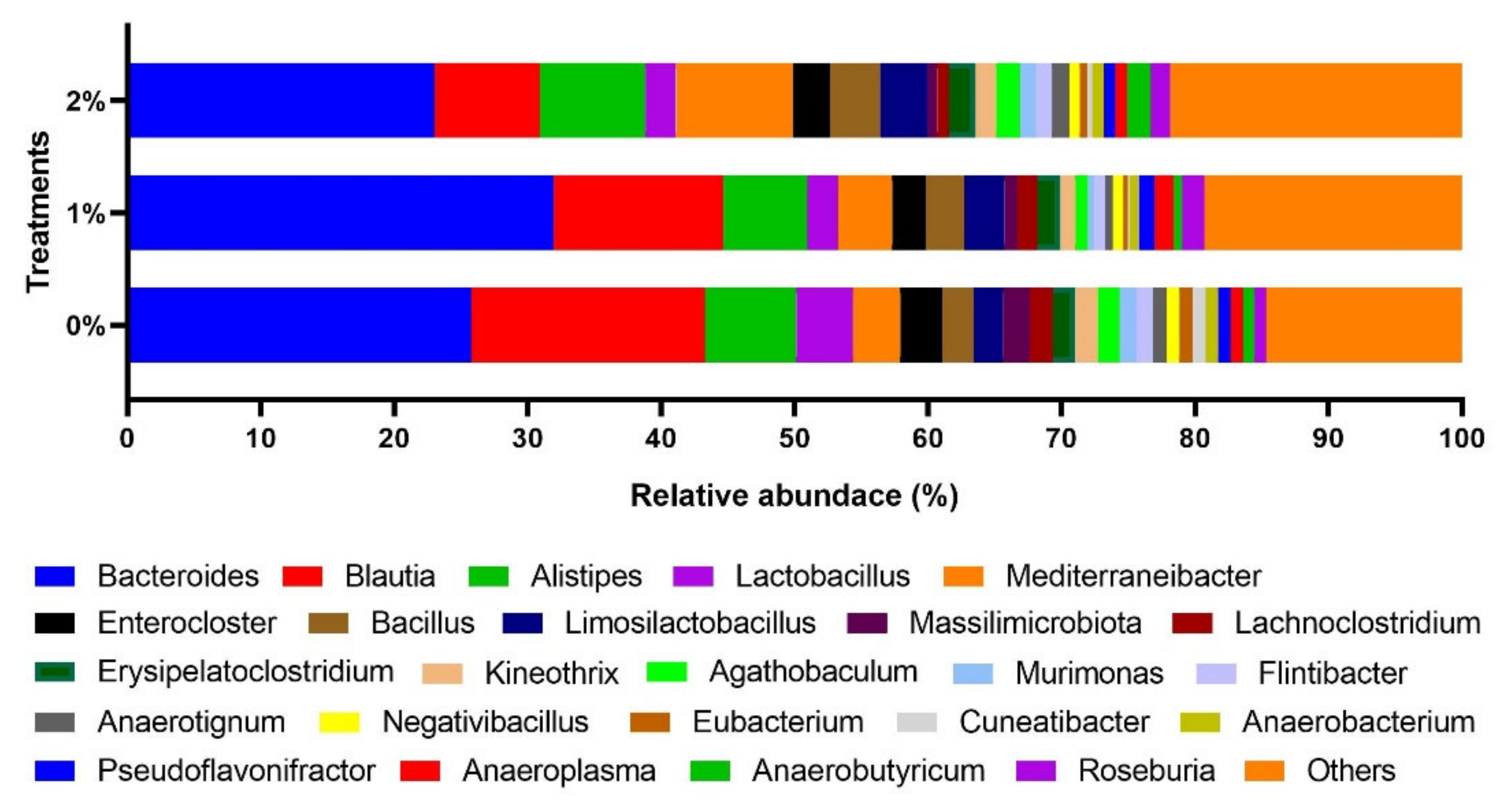
The effect of SPP supplementation on the cecum microbiota at the genus level is presented in
Figure 5. The top three dominant genus microbiotas were
Bacteroides (0% SPP: 25.8%, 1% SPP: 32.0%, and 2% SPP: 23.0%),
Blautia (0% SPP: 17.5%, 1% SPP: 12.7, and 2% SPP: 7.9), and
Alistipes (0% SPP: 6.8%, 1% SPP: 6.3, and 2% SPP: 7.9).
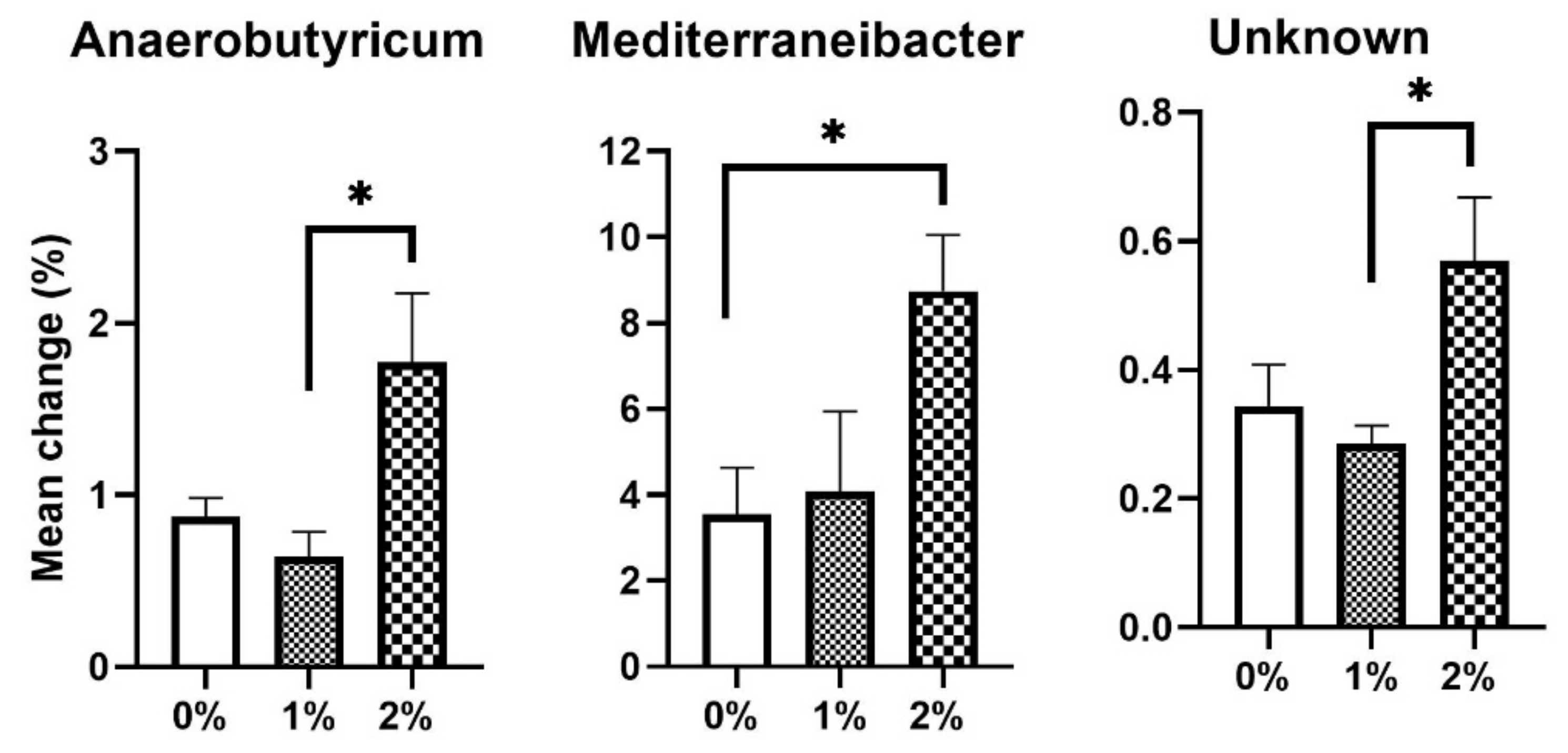
Anaerobutyricum and
Mediterraneibacter were the highly significant genera that were enhanced (
p < 0.05) in 2% SPP in comparison to 1% and 0% SPP-supplemented diets in chickens (
Figure 6). One more genus had a higher mean percent in 2% SPP- in comparison to 1% SPP-supplemented diets in chickens. However, the sequence of this genus is still unidentified.
4. Discussion
The SPP contains indigestible fibers and lignin that can act as a prebiotic in chicken diets. Various prebiotic products, such as mannanoligosaccharides (MOSs) and fructooligosaccharides (FOSs), have already been used in chicken diets [
15,
22]. In the present study, we used fibers from a pinewood source. Moreover, we used the steam explosion method to enhance the utilizable fiber. The benefit of using SPPs as a prebiotic is due to its cheap and sufficient availability.
In contrast, the availability of specialized prebiotic products, such as MOS, is inadequate and better techniques for the economical production of such products are yet to be explored to make it competitive enough to be used in livestock production [
23]. Furthermore, inconsistent results have been obtained by dietary supplementation of prebiotic products on growth performances. For instance, Attia and his group reported enhanced growth performance of chickens after supplementing prebiotic products, such as MOS, in chicken diets [
15]. Similarly, Mookiah and his group also reported an increase in chicken growth performances after dietary supplementation of a prebiotic product [
24]. Contrary to this, other researchers reported that dietary supplementation of prebiotics, such as FOS or MOS, did not affect the growth performance in chickens [
22,
23]. The discrepancy in the results could be attributed to the difference in the source of prebiotic products utilizing different raw materials and microorganism enzymatic activity, resulting in differential hydrolysis of polysaccharides [
23]. The motivation for using dietary fibers in the present study was different from those, such as FOS and MOS. The present study was conducted to utilize the byproducts from the wood industry while that of using specialized prebiotic products is to increase growth performances in chickens, making it incomparable for its effects on growth performance. However, an increase (42 g;
p > 0.05) in the final body weight of 2% SPP-supplemented chickens in a 21-day feeding trial in comparison to the control indicates the potential of SPP to positively influence the body weight in chickens. Further studies are warranted to evaluate the effect of dietary SPP after long-term supplementation.
A study conducted on steam-exploded
Quercus mongolica supplementation in chicken diets reported significantly lower ADG with increasing
Quercus mongolica concentration in comparison to the control [
6]. Contrary to this, no significant variations were seen in ADG in SPP-supplemented diets in the present study. These results could be due to the difference in the source of wood used in the studies. No negative effects on growth performance parameters indicate that supplementation of up to 2% SPP in chicken diets can consume industrial wood waste.
The gut is a vital organ responsible for the digestion and absorption of nutrients. Supplementation of dietary fibers has been known to possess modulatory roles in chickens by modifying gut health. We evaluated the relative organ weight and absolute length, which were similar and not affected by supplementation of increasing amounts of SPP in chicken diets. Our finding correlates with those of Wang et al. They reported a similar relative organ weight and absolute length at 14 and 27 days of age after dietary supplementation of an increasing concentration of prebiotics [
25]. The absorptive area is dependent on the length of the intestine and is directly correlated to nutrient digestion and body weights [
26]. We did not evaluate the digestion of nutrients. However, the similar ADG, ADFI, and FCR values in the current study correspond to no effects on intestinal length and weight in chickens. Furthermore, with increasing age, the absorptive capacity of the intestine depends mainly on the villus surface [
27]. This indicates that if the supplementation of dietary fibers, such as SPPs, been done for a more extended period, it may have improved the gut responses in chickens.
Steam explosion is directly correlated with the destruction of vegetative structure and its severity results in increasing cellulose and hemicellulose damage [
28,
29]. In the present study, plasma glucose levels showed a decreasing trend (
p > 0.05) with an increasing concentration of SPP-supplemented diets in comparison to the control. The reason behind this could be related to the availability of lower carbohydrates responsible for energy production due to the higher cellulose damage in SPP-supplemented diets. Similarly, numerically lower values (
p > 0.05) of plasma total protein were also observed in the SPP-supplemented diets in the present study. This can be attributed to the increase in inevitable crude protein and amino acid endogenous losses in broilers fed high fiber levels [
30]. Previous studies conducted on chickens fed modified dietary fiber from cassava pulp reported a decrease in fat, triglyceride, and cholesterol levels in chickens [
31]. Although not significant, numerically lower plasma triglyceride was seen in 2% SPP-supplemented diets in the present study.
Dietary fibers may act as prebiotic substances that can modulate the intestinal microbiota [
10]. It is expected that the non-digested fibers may act as a source for the proliferation of beneficial bacteria in the chicken gut. Recent studies suggested that dietary supplementation of alfalfa meal stimulated the proliferation of beneficial bacteria, such as
Lactobacillus and
Bacteroides, and inhibited potentially pathogenic bacteria including
Clostridium [
32]. Additionally, dietary supplementation of chicory forage has benefited intestinal health by increasing the microbial community diversity in the caecum of chickens [
33]. Similarly, it was reported that orange and grapefruit peels, which are rich in fiber content and used as feed additives in broiler diets, significantly modulated the gut microbiota profile [
34]. Contrary to our results, a study conducted by adding tannins and bacitracin to chickens’ diet reported similar OTUs to that of the control, while the cecal richness of bacitracin-treated animals remained significantly lower [
35]. In the present study, we used SPPs as a feed additive that was produced by pretreating pine particles (wood powder) with steam explosion to modify the fraction in terms of its fiber contents. However, this discrepancy can be attributed to the differences, such as the additives, broiler genetic line, and trial duration (i.e., 28 versus 42 days), thereby hindering the comparison between differently designed experiments. Furthermore, the cecum plays an important role in breaking up non-digestible fibers through the process of anaerobic fermentation [
11]. Thus, the evaluation of microbiota in the cecum through metagenome analysis may facilitate the determination of the beneficial effects of dietary SPPs in the chicken gut.
In general, OTUs are an indicator of relative richness, whereas Chao1 indicates the estimated richness of microflora. Similarly, Shannon, Inverse Simpson, and Goods coverage are indicators of microbial diversity. The lack of variation in the richness and diversity indicators suggests that SPP supplementation did not modify the microbial community. However, previous studies suggested that prebiotic substances as a source of fiber enhance the proliferation of favorable bacteria in the gut [
10]. A recent study conducted on chickens by supplementing 4% dietary fiber from 9 weeks to 20 weeks of age had higher alpha-diversity indices than 2% fibers in diets [
36]. These results confirm the role of dietary fiber in positively modifying the alpha-diversity in chickens. However, the lack of variation of the alpha-diversity indices in the present study could be due to the lower concentration (1% and 2% SPPs) of dietary SPPs and shorter period (3 weeks) of feed supplementation. Fluctuation of microbiota occurs in the early few weeks of age and can be noticed until 25 days of age [
37]. The ceca in the present experiment were collected on the 28th day of age, and thus an unstable microbial population may also lead to non-significant results.
The beta diversity was evaluated by using unweighted and weighted unifrac distances. Unweighted unifrac distances are used to identify the differences in existing or missing organisms. The similarity in the PCoA of unweighted unifrac distances indicates identical species in all the treatment groups. Weighted unifrac distances are used to differentiate the abundance of microbes. Dimension one and two of PCoA from weighted unifrac contributed 59.9% and 15.3% of the inertia, respectively, making the overall variability around 75%, indicating differences in the abundance of organisms among treatment groups. Differences in the abundance of the bacteria could be attributed to the availability of fibers from the dietary source supplemented with an increasing concentration of SPPs. The possibility of differences in the abundance of organisms could also exist due to the fluctuation in the microbiota during the first four weeks of age [
37], which requires further investigation with long-term experiments.
In the present study,
Firmicutes was the most abundant phyla followed by
Bacteroidetes and were found to be inversely correlated. These findings are in accordance with the previous studies showing that
Firmicutes and
Bacteroidetes were the dominant phyla throughout the ceca of chickens with the age of 7 days to 42 days [
38]. However, in another study,
Bacteroidetes was found to be the most abundant phylum in the chicken cecum [
39]. The difference in the results could be related to the different farm conditions, such as temperature and hygiene. The action of the bacterial phylum takes place through SCFAs. Acetate, butyrate, and propionate are the major SCFAs in chickens, which were found to be produced in the cecum of a high-fiber feed substrate when tested in vitro [
40].
Firmicutes is the main producer of butyrate, while
Bacteroidetes is mainly responsible for producing acetate and propionate in chickens [
41]. This indicates that dietary fiber in SPP supplementation may help maintain the SCFAs in chickens. Heat stress has been associated with modified phylum microbiota in chicken [
42]. Although we did not evaluate the effect of temperature or other aspects, modified fiber supplementation could effectively maintain the bacterial phylum in chickens and open the way for future research. In the present study,
Firmicutes and
Tenericutes were positively correlated.
Tenericutes was found to have an inverse relationship with
Bacteroidetes. Our findings agree with the previous finding that
Firmicutes and
Tenericutes also show a similar increasing trend in chickens [
42].
Bacteroides, Blautia, and
Alistipes were the three most abundant bacterial genera found in the cecum of the present study.
Blautia is known to metabolize glucose and dietary cellulose to produce SCFAs [
43,
44]. However, in the present study, the abundance of
Blautia showed a decreasing trend (
p > 0.05). The reason behind this could be related to the increase in the cellulosic damage in the SPP-supplemented diets due to the steam-explosion method applied for its production [
28,
29]. The direct association can also be seen with the decrease in the plasma glucose level in this study. In the present study,
Mediterraneibacter and
Anaerobutyricum are the only two known genera that also belong to the top 24 bacterial genera that were found to be significantly modified by the treatment. According to Togo et al. [
45] and Shetty et al. [
46],
Mediterraneibacter and
Anaerobutyricum are the reclassifications of
Ruminococcus and
Eubacterium, respectively. Due to the presence of high fiber in diets, we expect that fiber-degrading genera could have a higher abundance in the cecum of chickens fed SPP-supplemented diets. Previous studies suggested that
Ruminococcus and
Eubacterium are two known fibrolytic bacterial genera that can degrade fibers through enzymatic action [
47,
48]. They can degrade dietary fiber, especially from plant sources [
49]. The higher availability of fibers in the 2% SPP-supplemented diets may have increased the abundance of
Mediterraneibacter and
Anaerobutyricum in the cecum. One more unidentified genus, with a higher abundance in 2% SPP compared to 1% SPP, was observed and needs further investigation.



 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}