
Responses of Soil Abiotic Properties and Microbial Community Structure to 25-Year Cucumber Monoculture in Commercial Greenhouses

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling
2.2. Soil Abiotic Properties
2.3. Soil DNA Extraction and Quantitative PCR
2.4. MiSeq Sequencing
2.5. Statistical Analyses
3. Results
3.1. Soil Properties
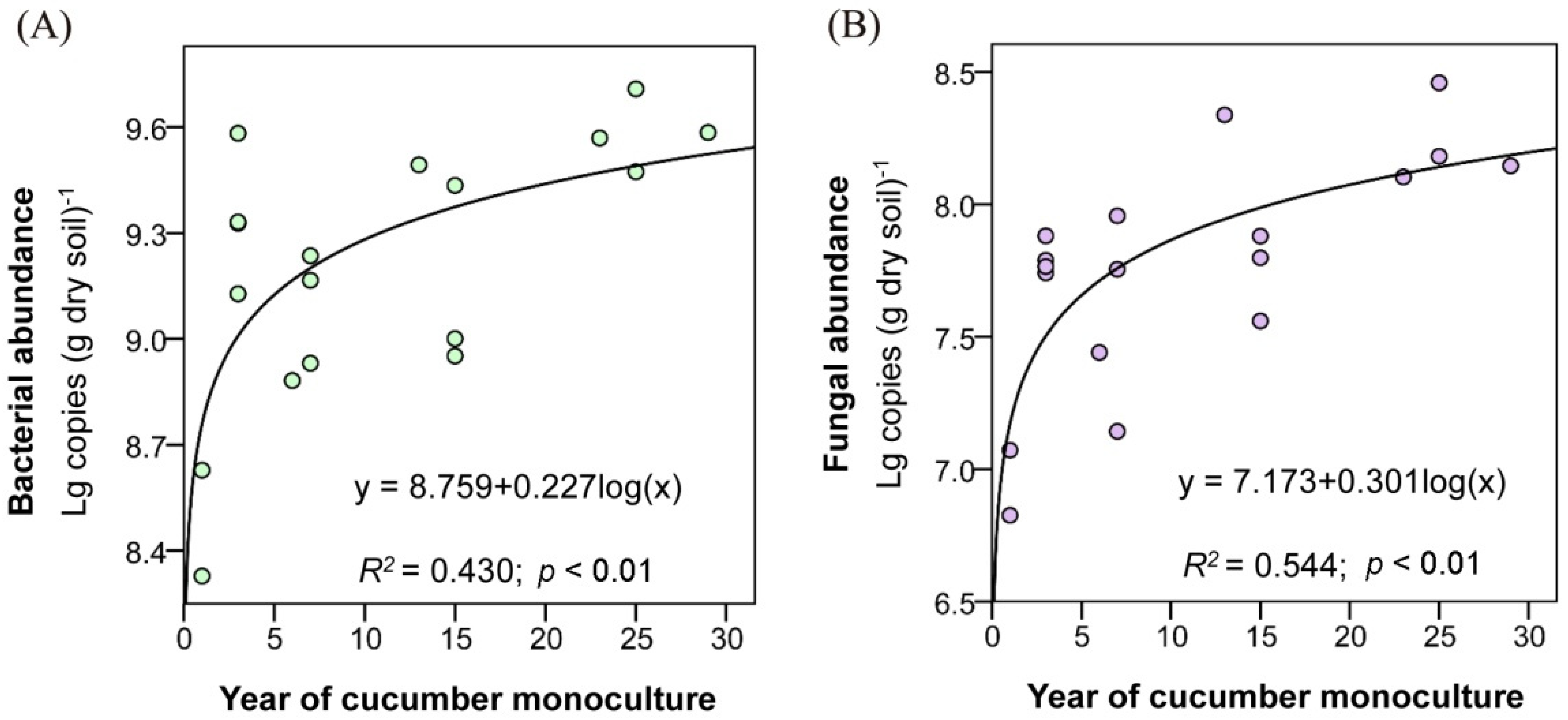
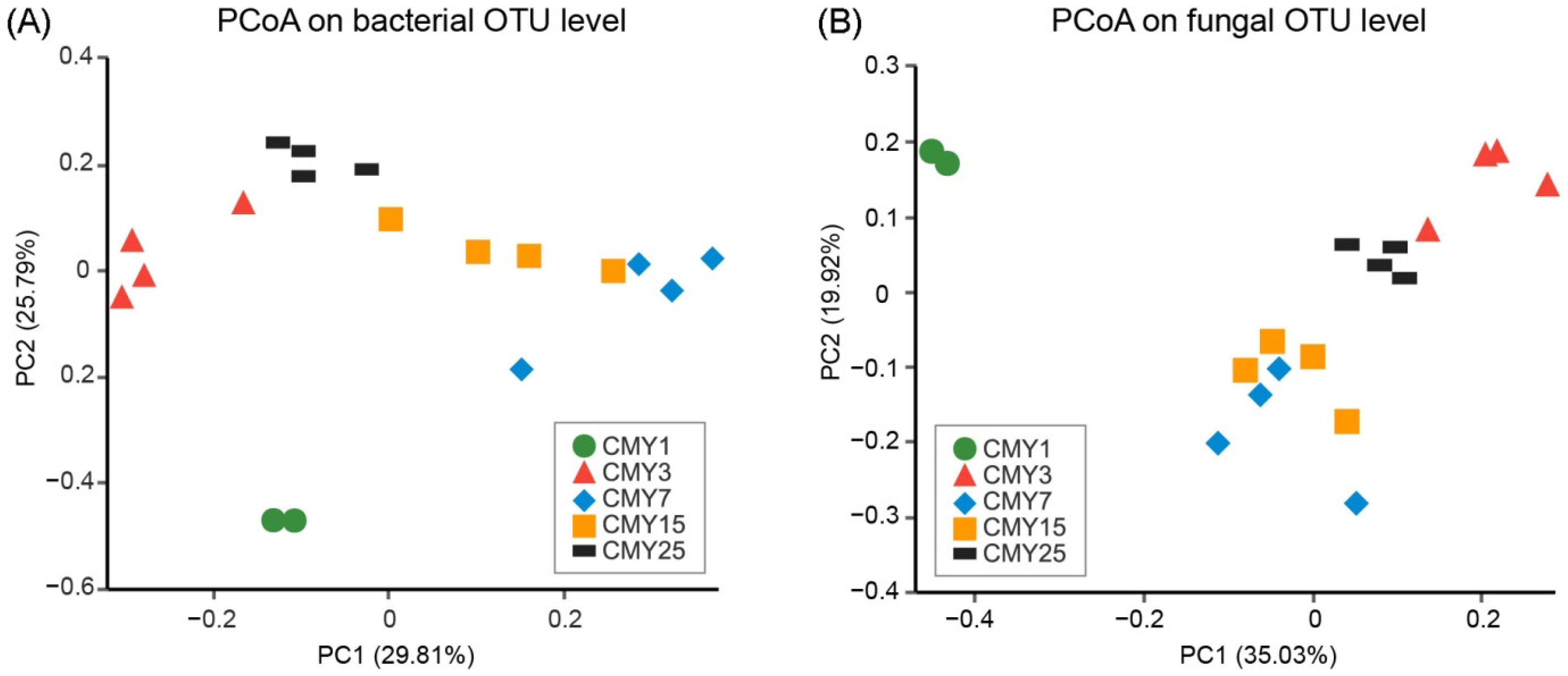
3.2. Soil Microbial Abundance and Diversity
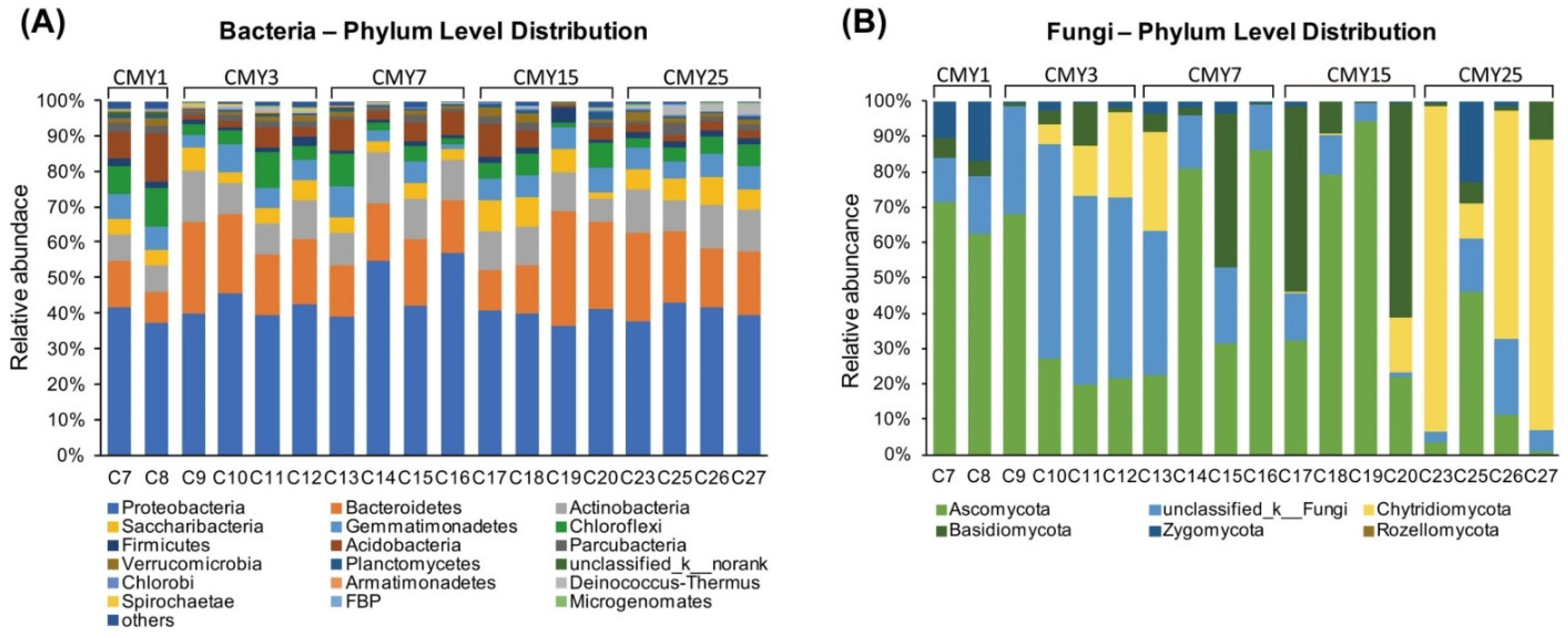
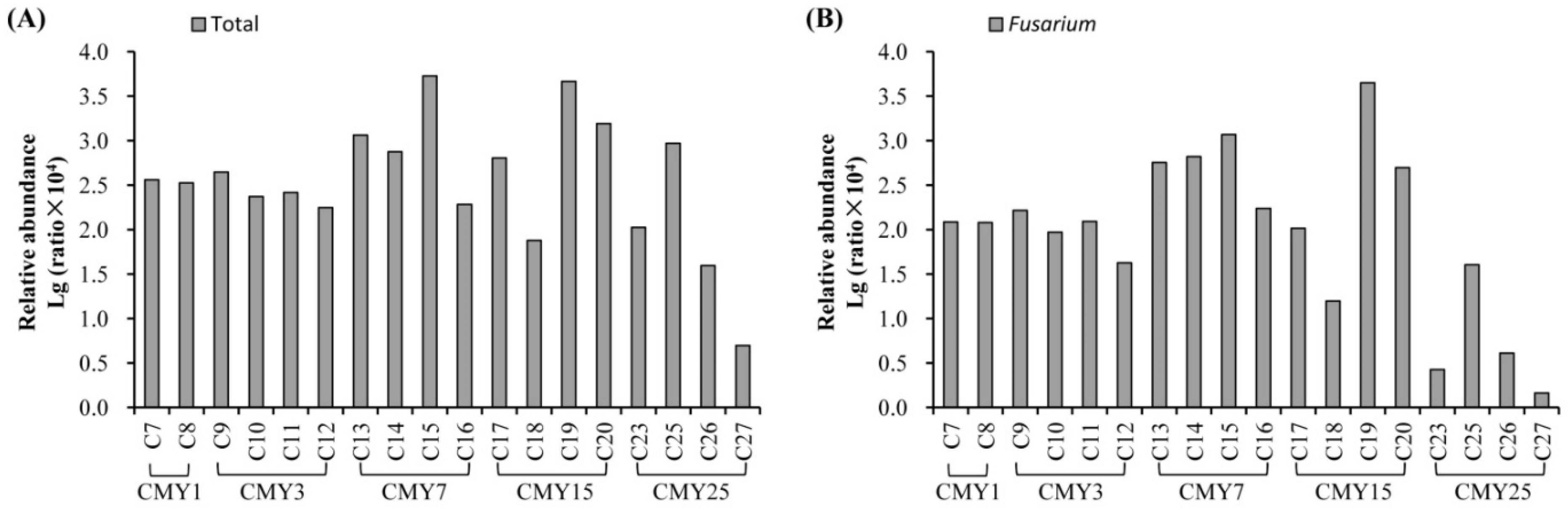
3.3. Soil Microbial Taxonomic Composition
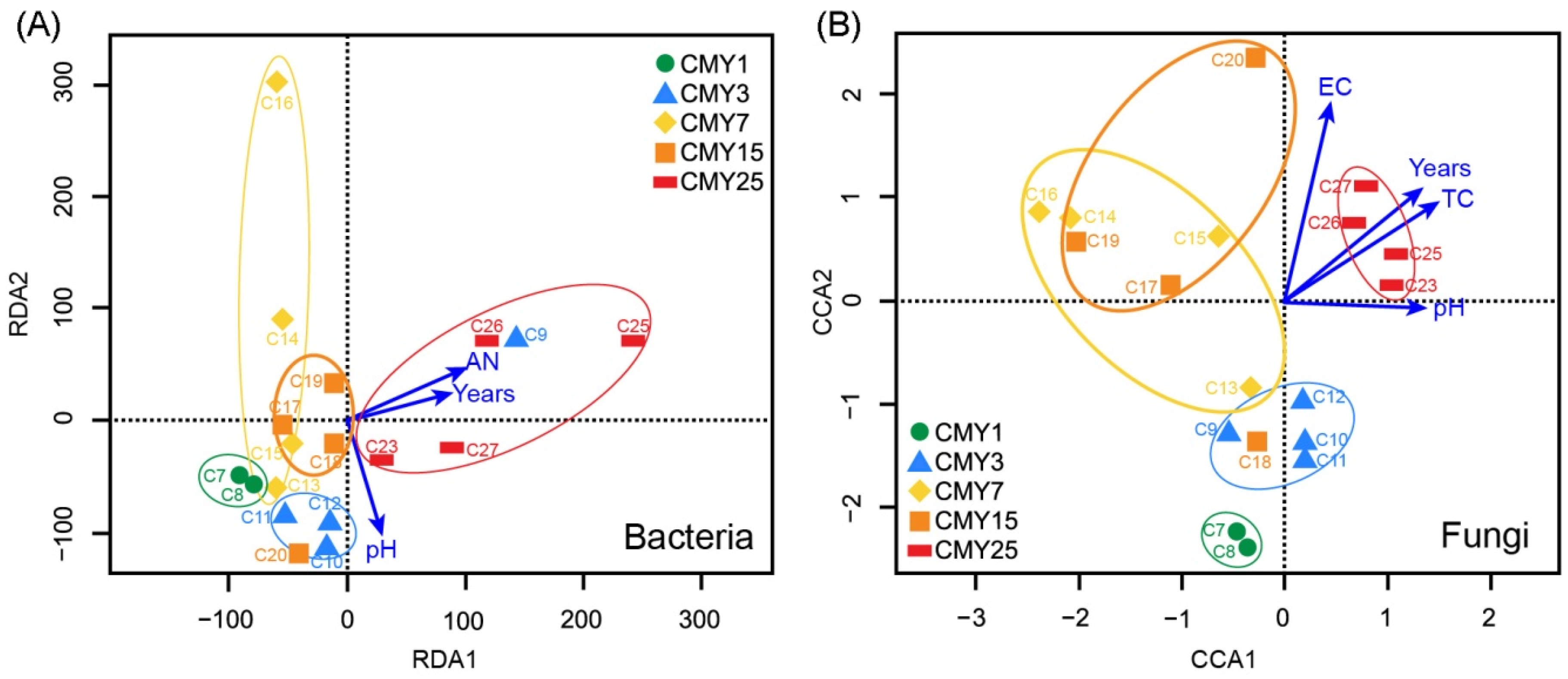
3.4. Correlations among Soil Abiotic Properties, Microbial Community Structure, and Monoculture Years
4. Discussion
4.1. Impacts of Long-Term Cucumber Monoculture on Soil Abiotic Properties
4.2. Impacts of Long-Term Monoculture on Soil Microbial Abundance and Structure
4.3. Correlation between Soil Abiotic Properties and Soil Microbial Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, X.; Wu, F. Dynamics of the diversity of fungal and Fusarium communities during continuous cropping of cucumber in the greenhouse. FEMS Microbiol. Ecol. 2012, 80, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Yu, H.; Yu, G.; Pan, K.; Bao, J. Improved bacterial community diversity and cucumber yields in a rotation with kidney bean-celery-cucumber. Acta Agric. Scand. Sect. B Soil Plant Sci. 2011, 61, 122–128. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, F. p-Coumaric acid influenced cucumber rhizosphere soil microbial communities and the growth of Fusarium oxysporum f.sp. cucumerinum Owen. PLoS ONE 2012, 7, e48288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Liu, J.; Wu, F. Soil microbial communities in cucumber monoculture and rotation systems and their feedback effects on cucumber seedling growth. Plant Soil 2017, 415, 507–520. [Google Scholar] [CrossRef]
- Li, Z.; Zu, C.; Wang, C.; Yang, J.; Yu, H.; Wu, H. Different responses of rhizosphere and non-rhizosphere soil microbial communities to consecutive Piper nigrum L. monoculture. Sci. Rep. 2016, 6, 35825. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yu, G.; Wu, F. Soil phenolics in a continuously mono-cropped cucumber (Cucumis sativus L.) system and their effects on cucumber seedling growth and soil microbial communities. Eur. J. Soil Sci. 2012, 63, 332–340. [Google Scholar] [CrossRef]
- Guo, L.; Han, L.; Yang, L.; Zeng, H.; Fan, D.; Zhu, Y.; Feng, Y.; Wang, G.; Peng, C.; Jiang, X.; et al. Genome and transcriptome analysis of the fungal pathogen Fusarium oxysporum f. sp. cubense causing banana vascular wilt disease. PLoS ONE 2014, 9, e95543. [Google Scholar]
- Ye, S.F.; Yu, J.Q.; Peng, Y.H.; Zheng, J.H.; Zou, L.Y. Incidence of Fusarium wilt in Cucumis sativus L. is promoted by cinnamic acid, an autotoxin in root exudates. Plant Soil 2004, 263, 143–150. [Google Scholar] [CrossRef]
- Zhou, X.; Yu, G.; Wu, F. Effects of intercropping cucumber with onion or garlic on soil enzyme activities, microbial communities and cucumber yield. Eur. J. Soil Biol. 2011, 47, 279–287. [Google Scholar] [CrossRef]
- Pervaiz, Z.H.; Iqbal, J.; Zhang, Q.; Chen, D.; Wei, H.; Saleem, M. Continuous cropping alters multiple biotic and abiotic indicators of soil health. Soil Syst. 2020, 4, 59. [Google Scholar] [CrossRef]
- Wang, L.; Yang, F.; Yaoyao, E.; Yuan, J.; Raza, W.; Huang, Q.; Shen, Q. Long-term application of bioorganic fertilizers improved soil biochemical properties and microbial communities of an apple orchard soil. Front. Microbiol. 2016, 7, 1893. [Google Scholar] [CrossRef]
- Han, J.; Luo, Y.; Yang, L.; Liu, X.; Wu, L.; Xu, J. Acidification and salinization of soils with different initial pH under greenhouse vegetable cultivation. J. Soils Sed. 2014, 14, 1683–1692. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Yang, F.; Yaoyao, E.; Raza, W.; Huang, Q.; Shen, Q. Application of bioorganic fertilizer significantly increased apple yields and shaped bacterial community structure in orchard soil. Microb. Ecol. 2017, 73, 404–416. [Google Scholar] [CrossRef]
- Yu, H.Y.; Li, T.X.; Zhang, X.Z. Nutrient budget and soil nutrient status in greenhouse system. Agric. Sci. China 2010, 9, 871–879. [Google Scholar] [CrossRef]
- Liu, J.; Sui, Y.; Yu, Z.; Shi, Y.; Chu, H.; Jin, J.; Liu, X.; Wang, G. High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China. Soil Biol. Biochem. 2014, 70, 113–122. [Google Scholar] [CrossRef]
- Agtmaal, M.V.; Straathof, A.; Termorshuizen, A.; Teurlincx, S.; Hundscheid, M.; Ruyters, S.; Busschaert, P.; Lievens, B.; Boer, W.D. Exploring the reservoir of potential fungal plant pathogens in agricultural soil. Appl. Soil Ecol. 2017, 121, 152–160. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, J.J.; Banerjee, S.; Zhou, N.; Zhao, Z.Y.; Zhang, K.; Tian, C.Y. Soil pH is equally important as salinity in shaping bacterial communities in saline soils under halophytic vegetation. Sci. Rep. 2018, 8, 4550. [Google Scholar] [CrossRef]
- Lu, R. Methods of Soil and Agricultural Chemistry Analysis; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Mills, D.A. Improved selection of internal transcribed spacer-specific primers enables quantitative, ultra-high-throughput profiling of fungal communities. Appl. Environ. Microbiol. 2013, 79, 2519–2526. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Gevers, D.; Westcott, S.L. Reducing the effects of PCR amplification and sequencing Artifacts on 16s rRNA-based studies. PLoS ONE 2011, 6, e27310. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Lou, J.; Yang, L.; Wang, H.; Wu, L.; Xu, J. Assessing soil bacterial community and dynamics by integrated high-throughput absolute abundance quantification. PeerJ 2018, 2018, e4514. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Zhang, M.M.; Wang, N.; Hu, Y.B.; Sun, G.Y. Changes in soil physicochemical properties and soil bacterial community in mulberry (Morus alba L.)/alfalfa (Medicago sativa L.) intercropping system. Microbiologyopen 2018, 7, e555. [Google Scholar] [CrossRef]
- Giacometti, C.; Cavani, L.; Baldoni, G.; Ciavatta, C.; Marzadori, C.; Kandeler, E. Microplate-scale fluorometric soil enzyme assays as tools to assess soil quality in a long-term agricultural field experiment. Appl. Soil Ecol. 2014, 75, 80–85. [Google Scholar] [CrossRef]
- Wei, M.; Hu, G.; Wang, H.; Bai, E.; Lou, Y.; Zhang, A.; Zhuge, Y. 35 years of manure and chemical fertilizer application alters soil microbial community composition in a Fluvo-aquic soil in Northern China. Eur. J. Soil Biol. 2017, 82, 27–34. [Google Scholar] [CrossRef]
- Shi, W.M.; Yao, J.; Yan, F. Vegetable cultivation under greenhouse conditions leads to rapid accumulation of nutrients, acidification and salinity of soils and groundwater contamination in South-Eastern China. Nutr. Cycl. Agroecosyst. 2009, 83, 73–84. [Google Scholar] [CrossRef]
- Shen, W.; Ni, Y.; Gao, N.; Bian, B.; Zheng, S.; Lin, X.; Chu, H. Bacterial community composition is shaped by soil secondary salinization and acidification brought on by high nitrogen fertilization rates. Appl. Soil Ecol. 2016, 108, 76–83. [Google Scholar] [CrossRef]
- Kingery, W.L.; Wood, C.W.; Delaney, D.P.; Williams, J.C.; Mullins, G.L. Impact of long-term land application of broiler litter on environmentally related soil properties. J. Environ. Qual. 1994, 23, 139–147. [Google Scholar] [CrossRef]
- Ding, J.; Ma, M.; Jiang, X.; Liu, Y.; Zhang, J.; Suo, L.; Wang, L.; Wei, D.; Li, J. Effects of applying inorganic fertilizer and organic manure for 35 years on the structure and diversity of ammonia-oxidizing archaea communities in a Chinese Mollisols field. MicrobiologyOpen 2020, 9, e00942. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Zhang, J.; Shi, Y.; Wu, F.; Zhou, X. Green manures of Indian mustard and wild rocket enhance cucumber resistance to Fusarium wilt through modulating rhizosphere bacterial community composition. Plant Soil 2019, 441, 283–300. [Google Scholar] [CrossRef]
- Andronov, E.E.; Petrova, S.N.; Pinaev, A.G.; Pershina, E.V.; Rakhimgaliyeva, S.; Akhmedenov, K.M.; Gorobets, A.V.; Sergaliev, N.K. Analysis of the structure of microbial community in soils with different degrees of salinization using T-RFLP and real-time PCR techniques. Eurasian Soil Sci. 2012, 45, 147–156. [Google Scholar] [CrossRef]
- Meng, Q.; Yin, J.; Rosenzweig, N.; Douches, D.; Hao, J.J. Culture-based assessment of microbial communities in soil suppressive to potato common scab. Plant Dis. 2012, 96, 712–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Year of Monoculture | EC a (μs·cm−1) | pH | SOC b (g·kg−1) | TN c (g·kg−1) | C/N | AN d (mg·kg−1) | TP e (‰) | TK f % | 16S Gene (×108 Copies·g−1) | 18S Gene (×106 Copies·g−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CMY1 | 1 | 215.9 a | 6.23 b | 10.53 a | 1.39 a | 7.56 ab | 210.12 a | 0.47 a | 2.475 a | 2.13–4.25 | 6.7–11.8 |
| CMY3 | 3 | 547.8 a | 6.21 b | 24.39 b | 3.39 b | 7.20 a | 497.30 b | 1.06 abc | 2.378 a | 13.41–38.27 | 55.1–76.1 |
| CMY7 | 6~7 | 1002 b | 4.80 a | 34.87 bc | 3.71 b | 9.37 b | 567.32 bc | 0.99 ab | 2.478 a | 7.62–17.21 | 13.9–90.6 |
| CMY15 | 13~15 | 1165 b | 5.48 ab | 36.90 c | 4.43 b | 8.46 ab | 743.38 c | 1.18 bc | 2.430 a | 8.95–31.13 | 36.3–218.0 |
| CMY25 | 23~29 | 1385 b | 5.85 b | 58.94 d | 7.27 c | 8.13 ab | 1088.96 d | 1.70 c | 2.155 a | 29.74–51.09 | 127.0–288.0 |
| Diversity Indices | Sobs | Chao | Shannon | Heip | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Analytical Method | Pearson | Spearman | Pearson | Spearman | Pearson | Spearman | Pearson | Spearman | ||
| Bacteria | Years a | Cor | −0.392 | −0.523 * | −0.432 | −0.558 * | −0.226 | −0.490 * | −0.252 | −0.472 * |
| Sig | 0.108 | 0.026 | 0.073 | 0.016 | 0.367 | 0.039 | 0.313 | 0.048 | ||
| EC b | Cor | −0.635 ** | −0.664 ** | −0.614 ** | −0.682 ** | −0.526 * | −0.653 ** | −0.568 * | −0.598 ** | |
| Sig | 0.005 | 0.003 | 0.007 | 0.002 | 0.025 | 0.003 | 0.014 | 0.009 | ||
| pH | Cor | 0.412 | 0.502 * | 0.342 | 0.471 * | 0.495 * | 0.526 * | 0.439 | 0.527 * | |
| Sig | 0.089 | 0.034 | 0.165 | 0.048 | 0.037 | 0.025 | 0.068 | 0.025 | ||
| AN c | Cor | −0.512 * | −0.548 * | −0.540 * | −0.575 * | - | - | - | - | |
| Sig | 0.030 | 0.019 | 0.021 | 0.013 | - | - | - | - | ||
| Fungi | Years a | Cor | −0.630 ** | −0.709 ** | −0.649 ** | −0.735 ** | −0.542 * | −0.662 ** | −0.469 * | −0.487 * |
| Sig | 0.005 | 0.001 | 0.004 | 0.001 | 0.02 | 0.003 | 0.05 | 0.04 | ||
| EC b | Cor | −0.827 ** | −0.808 ** | −0.805 ** | −0.789 ** | −0.652 ** | −0.736 ** | −0.520 * | −0.538 * | |
| Sig | <0.000 | <0.000 | <0.000 | <0.000 | 0.003 | 0.001 | 0.027 | 0.021 | ||
| AN c | Cor | −0.67 0 ** | −0.633 ** | −0.688 ** | −0.668 ** | −0.485 * | −0.562 * | - | - | |
| Sig | 0.002 | 0.005 | 0.002 | 0.002 | 0.041 | 0.015 | - | - | ||
| SOC d | Cor | −0.480 * | −0.41 | −0.511 * | −0.443 | - | - | - | - | |
| Sig | 0.043 | 0.091 | 0.03 | 0.066 | - | - | - | - | ||
| TN e | Cor | −0.469 * | −0.325 | −0.508 * | −0.385 | - | - | - | - | |
| Sig | 0.050 | 0.188 | 0.031 | 0.115 | - | - | - | - | ||
| Soil Properties | Correlation | Bacteria | Fungi | ||
|---|---|---|---|---|---|
| Pearson | Spearman | Pearson | Spearman | ||
| SOC a | Cor | 0.633 ** | 0.488 * | 0.605 ** | 0.624 ** |
| Sig | 0.005 | 0.040 | 0.008 | 0.006 | |
| TN b | Cor | 0.699 ** | 0.550 * | 0.617 ** | 0.593 ** |
| Sig | 0.001 | 0.018 | 0.006 | 0.009 | |
| TP c | Cor | 0.536 * | 0.470 * | - | 0.480 * |
| Sig | 0.022 | 0.049 | - | 0.044 | |
| AN d | Cor | 0.711 ** | 0.610 ** | 0.557 * | 0.633 ** |
| Sig | 0.001 | 0.007 | 0.016 | 0.005 | |
| EC e | Cor | 0.527 * | 0.583 * | - | 0.564 * |
| Sig | 0.025 | 0.011 | - | 0.015 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.-H.; Lu, X.-H.; Guo, R.-J.; Hao, J.-J.; Miao, Z.-Q.; Yang, L.; Li, S.-D. Responses of Soil Abiotic Properties and Microbial Community Structure to 25-Year Cucumber Monoculture in Commercial Greenhouses. Agriculture 2021, 11, 341. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11040341
Gao Y-H, Lu X-H, Guo R-J, Hao J-J, Miao Z-Q, Yang L, Li S-D. Responses of Soil Abiotic Properties and Microbial Community Structure to 25-Year Cucumber Monoculture in Commercial Greenhouses. Agriculture. 2021; 11(4):341. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11040341
Chicago/Turabian StyleGao, Yu-Han, Xiao-Hong Lu, Rong-Jun Guo, Jian-Jun Hao, Zuo-Qing Miao, Li Yang, and Shi-Dong Li. 2021. "Responses of Soil Abiotic Properties and Microbial Community Structure to 25-Year Cucumber Monoculture in Commercial Greenhouses" Agriculture 11, no. 4: 341. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11040341