
Cover Cropping Impacts Soil Microbial Communities and Functions in Mango Orchards

1
College of Resources and Environment, Huazhong Agricultural University, Wuhan 430070, China
2
Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, China
*
Author to whom correspondence should be addressed.
Agriculture 2021, 11(4), 343; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11040343
Submission received: 2 March 2021
/
Revised: 26 March 2021
/
Accepted: 27 March 2021
/
Published: 12 April 2021
(This article belongs to the Special Issue Soil Carbon and Microbial Processes in Agriculture Ecosystem)
Abstract
:Soil microbes play critical roles in nutrient cycling, net primary production, food safety, and climate change in terrestrial ecosystems, yet their responses to cover cropping in agroforestry ecosystems remain unknown. Here, we conducted a field experiment to assess how changes in cover cropping with sown grass strips affect the fruit yields and quality, community composition, and diversity of soil microbial taxa in a mango orchard. The results showed that two-year cover cropping increased mango fruit yields and the contents of soluble solids. Cover cropping enhanced soil fungal diversity rather than soil bacterial diversity. Although cover cropping had no significant effects on soil bacterial diversity, it significantly influenced soil bacterial community compositions. These variations in the structures of soil fungal and bacterial communities were largely driven by soil nitrogen, which positively or negatively affected the relative abundance of both bacterial and fungal taxa. Cover cropping also altered fungal guilds, which enhanced the proportion of pathotrophic fungi and decreased saprotrophic fungi. The increase in fungal diversity and alterations in fungal guilds might be the main factors to consider for increasing mango fruit yields and quality. Our results indicate that cover cropping affects mango fruit yields and quality via alterations in soil fungal diversity, which bridges a critical gap in our understanding of the linkages between soil biodiversity and fruit quality in response to cover cropping in orchard ecosystems.
1. Introduction
Cover cropping (i.e., sown grass strips) has been used as an important and effective method to improve soil fertilizer and soil carbon stacks [1]. A 12-year field experiment indicated that cover cropping increased soil organic carbon (SOC) stocks [1]. A meta-analysis showed that cover cropping contributed to the changes in global cropland soil carbon, with an overall mean change of 15.5% [2]. Compared with monospecies cover crops, cover crop mixtures sequestered more SOC [2]. Elevated SOC is also associated with improved soil health and fertility; therefore, increasing SOC may help to enhance agricultural productivity [3]. However, not all studies found that cover cropping results in SOC accumulation. Some studies demonstrated that the introduction of cover crops resulted in losses of SOC due to the faster growth of cover crops [4]. In addition to an increased carbon input, cover crops have been shown to increase biodiversity [5].
Soil harbors a rich diversity of invertebrate and microbial life, which drives biogeochemical processes at local and global scales. Soil microbes play critical roles in the nutrient cycling, climate regulation, decomposition and turnover of soil organic matter [6]. Cover cropping alters soil quality and thereby influences soil microbial communities in agro-ecosystems. The potential of cover crops to increase soil biodiversity and specific microbial patterns has been highlighted in very few studies, especially in fruit orchards. For example, sowing plant seed mixtures promoted the growth of the bacterial community and sarophytic fungi [7]. The long-term effects of green manure amendment are altered soil microbial properties, first found in a field experiment carried out in 1956 [8]. The soil microbiome plays important roles in fruit quality and production, as rhizosphere microbiome maintains plant health and primary productivity [9]. The plant-associated microbial community has been considered as the second genome of the plant, which is critical for plant health. Therefore, the cover cropping in orchards enhances the complexity and diversity of soil microbes in intensive agriculture soils, which in turn strongly influences plant health and net primary production.
The mango (Mangifera indica L.) is the most important fruit crop in the tropical zones, having socio-economic significance, originating from South East Asia and cultivated worldwide [10]. It is known as the king of fruits owing to its delicious taste and high vitamin C and mineral contents [11]. Mango fruit has become the second tropical crop in terms of production and cultivated acreage [12]. To steadily increase fruit yield, farmers have increased the use of chemical fertilizers, which has caused many environmental problems, such as soil degradation and water contamination [13]. Cover cropping began to be used in orchards to resolve degraded soils and maintain higher crop yields and quality [14]. This green manure method has been widely used in the fruit ecosystem worldwide. The positive effects of the long-term application of green manure have been reported by many studies [15,16]. For example, some studies found grass cover to affect SOC [17,18,19]. Soil carbon for plantings of switchgrass, no-till corn, and sweetgum with cover crops between the rows increased over the first 3 years [20]. However, the variations in the structures of the microbial community and diversity in soils caused by cover crops remain unknown, which have been considered as the main drivers of multiple soil functions and plant health. Therefore, the aims of this study are to identify (1) the effects of different sown grass types in a mango orchard on the diversity of soil bacteria and fungi; (2) the changes in the main taxa in response to sown grass trips; (3) the driving factors of the variations in soil microbial diversity and community structure; and (4) the link between soil microbial diversity, mango yield and fruit quality.
2. Materials and Methods
2.1. Site Description and Sampling
Mango is among the most important fruits in Hainan Province, Southern China. The planted area of this fruit is more than 25,000 ha, and the yields are more than 560,000 kg per year. The field experiment was conducted at the mango base in Tianya District, Sanya City, Hainan Province, China (109°24′ 52.70″ (E) and 18°19′54.10″ (N)). It belongs to the tropical maritime monsoon climate zone, with an annual average temperature of 25.7 °C. The highest temperature is in June, with an average of 28.7 °C. The lowest temperature is in January, with an average of 21.4 °C. The annual duration of sunshine is 2534 h. The annual average precipitation is 1347.5 mm. The terrain is a gentle area in the lower hills, and soil type is latosol [21].
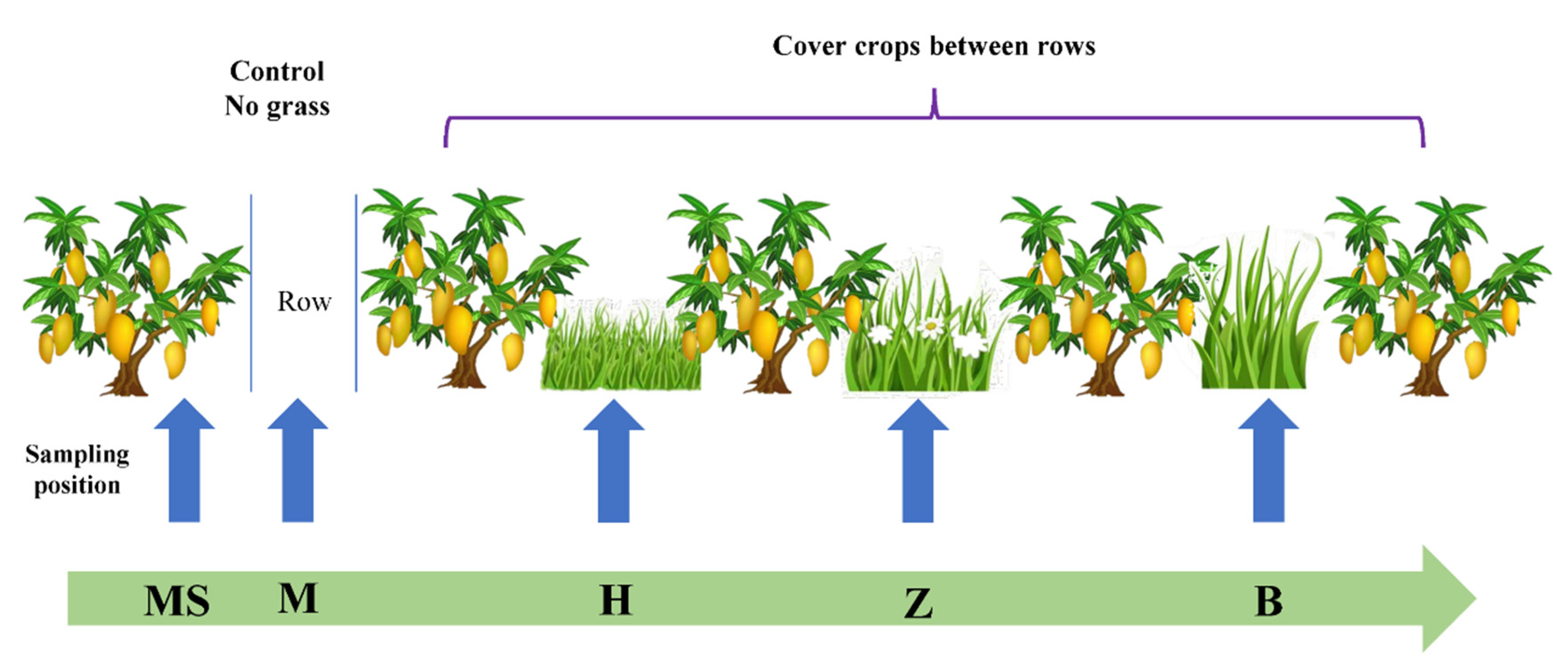
A total of four treatments with different types of cover cropping were implemented, including control (no-till + herbicide, M), planting Stylosanthes guianensis (Z), planting Brachiaria eruciformis (B) between rows, and planting Butterfly pea (H). Each treatment was repeated three times, with a plot area of 120 m2 (10 × 12 m) (Figure 1). All the plots were arranged in a randomized block design and separated by five 5 m buffer zones. The cover cropping was carried out in 2017. All treatments were transplanted with seedlings. Two rows of grass were planted between each mango row. The tested mango was Hongjinlong, with an age of 13 years. The row space between mango trees was 5 m. Fertilizers were applied in accordance with the conventional fertilization of fruit farmers, and no fertilizer was applied between rows.
Soils were collected in July (summer) 2019. In the control, soils were collected between the rows (M) and the drip line of mango trees (MS). Other soils (H, Z and B) with cover crops were collected between the rows. In each experimental plot, we collected 20 surface soil cores (0–20 cm and 20–40 cm) randomly between rows (Figure 1). All the soil samples in one experimental plot were mixed. In total, 18 samples per soil layer were collected. After removing the roots, stones and litters by hand, we sieved them through a 2 mm sieve and separated them into two portions. One portion (approximately 5 g) was preserved in a 10 mL centrifuge tube (sterilized) and stored at −80 °C. The second portion was air-dried and used to measure soil properties.
2.2. Mango Fruit Quality and Yield
When mango fruits were nearly matured (80%), all the mango fruits in each experimental plot were harvested, and the masses were measured as yields. We randomly chose 20 mango fruits in each experimental plot and placed them into plastic bags for analysis of the fruit quality. Total soluble solids content of mango fruit was measured by a digital refractometer and presented in percent Brix (%TSS). Organic acid concentration was measured by an automatic titrator [22]. Vitamin C (Vc) content was determined via the method of 2,6-dichlorophenol indophenol (Shao et al. 2013).
2.3. Analysis of Soil Properties
Soil properties were measured by the standardized methods as described previously. Soil bulk density (BD) was measured by the volume–mass relationship via a cutting ring [23]. Soil pH was determined by a glass pH meter in a 1:2.5 soil–water suspension. Soil organic matter (SOM) was digested with 5 mL concentrated H2SO4 and 5 mL 0.8 M K2Cr2O7, and then determined by 0.2 M Ferrous Ammonium Sulfate [24]. Soil total nitrogen (TN) was determined as described previously [25]. Soil available nitrogen (AVN) was extracted by 1 mol/L KCl and then measured by a Seal Auto Analyzer3 [26]. Soil total phosphorus (TP) and soil available P (AVP) were determined by molybdenum, antimony and scandium colorimetry. Total potassium (TK) and available potassium (AVK) in soil were determined by a flame photometer.
2.4. Molecular Analysis
We used the PowerSoil kit to extract DNA from 0.5 g soils. The protocol was listed in the manufacturer’s instructions. After extraction, the ratios of A260/A230 and A260/A280 were determined to assess the quality of extracted DNA.
The soil bacterial community was determined by sequencing hypervariable V3-V4 regions of 16S rRNA genes using primers 338F (ACTCCTACGGGAGGCAGCA) and 806R (GGACTACHVGGGTWTCTAAT) [27]. The PCR amplification conditions for 16S rRNA were 50 s at 94 °C, 30 s at 40 °C, 35 cycles of 60 s at 72 °C, followed by 5 min at 72 °C [28]. The ITS1 variable region was sequenced with primer sets ITS3 (5′-GCATCGATGAAGAACGCAGC-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) [29,30] to assess the soil fungal community. As for the ITS1 variable region, the PCR program was 5 min at 94 °C, 32 cycles of 30 s at 94 °C, 30 s at 54 °C, and 1 min at 72 °C [31]. Sequencing was conducted on an Illumina MiSeq PE300 platform.
After sequencing, bioinformatics processing was performed using QIIME, USEARCH and UNIOISE3 [32]. After removing the short sequences (<20 nucleotides) and chimeric sequences, the remaining sequences were clustered into operational taxonomic units (OTUs) using 97% similarity [33] (version 7.1 http://drive5.com/uparse/). Taxonomy was assigned against the Greengenes (16S gene, Release 13.5 http://greengenes.secondgenome.com/) and UNITE (ITS gene, Release 7.2 http://unite.ut.ee/index.php) databases. The Shannon diversity index and richness (the numbers of OTUs) were calculated to express the soil bacterial and fungal diversity.
2.5. Fungal Ecological Guilds Identification
We used FUNGuild to identify the fungal ecological guilds using fungal OTU dataset with taxon assignments [34]. This prediction method has been widely used in gaining insights into the distributions of soil fungal ecological groups [35,36]. In this study, we used the data with confidence levels of “highly probable” and “probable” to perform further analysis.
2.6. Statistical Analysis
We first used ANOVA to compare the differences of soil properties, mango fruit yield and quality, and bacterial and fungal diversity among different treatments on SPSS 20.0 (IBM Corporation, Armonk, NY, USA). We then conducted Pearson correlation analyses between the microbial diversity (Shannon diversity index) and soil properties. Prior to ANOVA and Pearson correlation analyses, the data were used to conduct a log transformation to meet the normality and homogeneity. Multiple regression models were constructed to compare the effects of soil properties on microbial diversity in R 3.5. Anosim analysis was performed to compare the effects of sown grass strips on the soil microbial community structure in R 3.5. The bacterial and fungal community structure was calculated based on the Bray–Curtis dissimilarity and visualized by a nonmetric multidimensional scaling (NMDS) plot in R 3.5 with the vegan package [37]. The effects of soil properties on the bacterial and fungal community structure were analyzed by the Mantel test in R 3.5. The associations between the soil microbial community structure and soil properties were determined by the Mantel test in R 3.5 [38].
3. Results
3.1. Soil Properties of the Tested Orchard
The impacts of cover cropping with sown grass strips on soil properties depended on the types of sown grass and soil layers. The upper soils (0–20 cm) were more sensitive to the application of sown grass strips. B20 had the highest soil AVN, which was significantly higher than other sown grass types. The application of sown grass strips had no significant effects on soil pH, SOM and AVK at both soil layers (Table 1).
3.2. Yield and Fruit Quality of Mango under Different Sown Grass Trips
The application of sown grass trips had significant effects on mango fruit yield, with the highest yield in B (Table 2). There were no significant differences observed between Z and M. The applications of B, H and Z significantly enhanced TSS content and decreased organic acid compared to M.
3.3. Variations in Soil Microbial Diversity under Different Sown Grass Strips
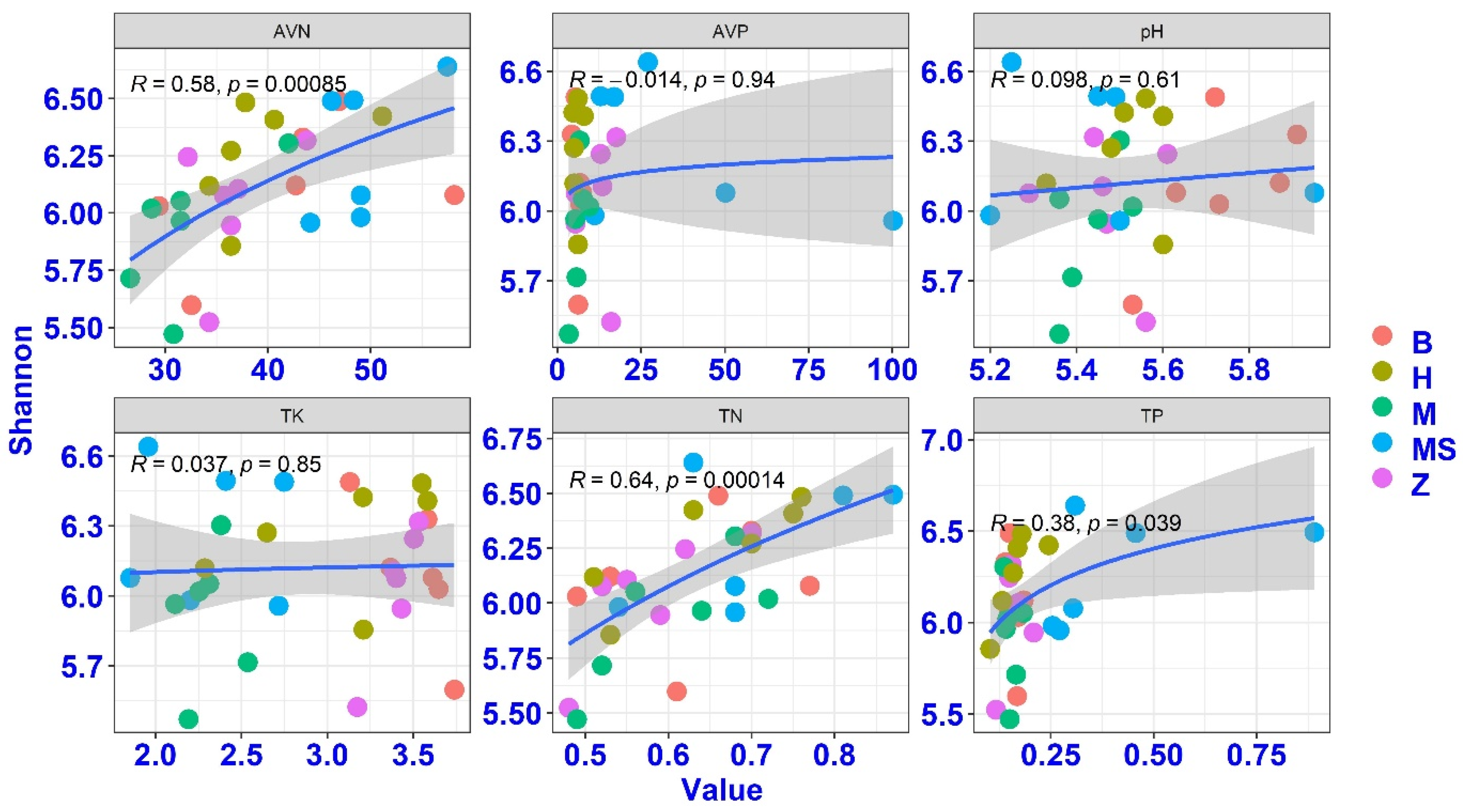
The observed bacterial Shannon diversity index ranged from 5.47 to 6.64, while phylotype richness (OTUs) varied from 1886 to 3509. Sown grass strips have no significant effects on soil bacterial α-diversity indices (Shannon diversity index and richness index). The soil fungal phylotype richness ranged from 419 to 1080, and the fungal Shannon diversity index ranged from 3.81 to 4.95. Among these soil properties, TN and AVN had significant associations with the bacterial Shannon diversity index (Figure 2). A multiple regression models indicated that soil TN and AVN were the best predictors of the soil bacterial Shannon diversity index (with a relative importance of 0.54 and 0.42), followed by AVP, with a relative importance of 0.04. Soil properties had no significant correlations with the fungal diversity index.
3.4. Variations in Soil Microbial Community under Different Sown Grass Strips
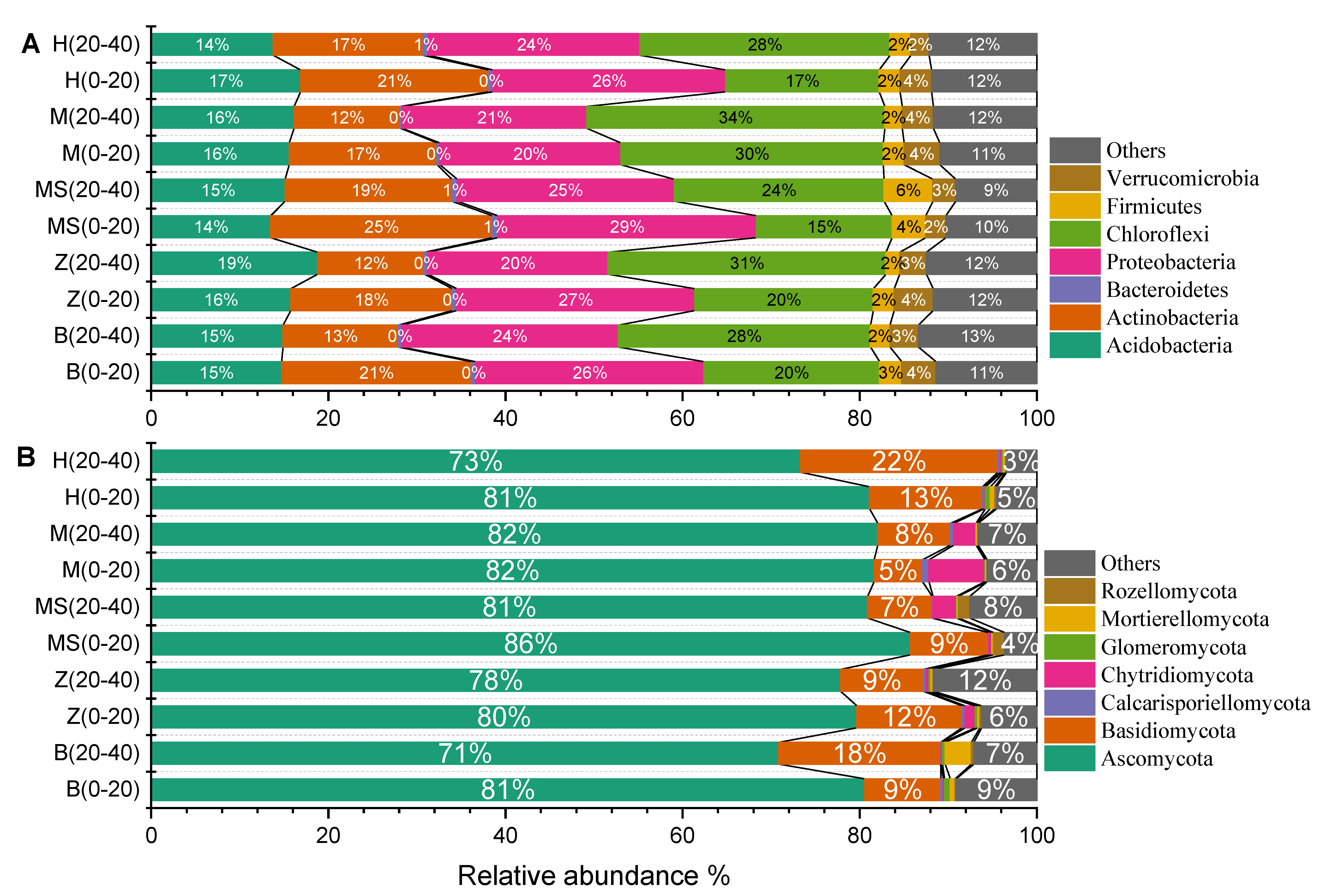
Across all soils, a total of 1,506,304 quality bacterial sequences, and an average of 50,210 sequences per sample, were obtained. Four of the 28 phyla detected were dominant, including Acidobacteria (15.51%), Actinobacteria (17.55%), Proteobacteria (24.26%) and Chloroflexi (24.77%) (with an average relative abundance of >5%, n = 30), accounting for more than 82% of the bacterial sequences (Figure 3A). The Proteobacteria taxa were dominated by Alphaproteobacteria (17.66%), followed by Deltaproteobacteria (4.12%) and Gammaproteobacteria (2.49%). The soil layer had no significant effects on the relative abundance of Acidobacteria and Proteobacteria. The upper soils (20.5%) had a higher relative abundance of Actinobacteria than that of the lower soils (14.6%).
Across all mango soils, a total of 1,777,037 quality fungal sequences, and an average of 59,234 sequences per sample, were obtained. The dominant fungal phyla in soils were Ascomycota and Basidiomycota, with average relative abundances of 79.38 and 11.28%, respectively (Figure 3B). Other minor phyla (Anthophyta, Cercozoa, Rozellomycota and Glomeromycota) were also found at a lower relative abundance (relative abundance < 1%). The soil layer and sown grass trip had no significant effects on the relative abundance of Ascomycota and Basidiomycota. Based on taxonomical classification at the class level, Sordariomycetes (36.69%), Eurotiomycetes (18.96%), Dothideomycetes (15.33%) and Agaricomycetes (9.50%) were more abundant than other groups (relative abundance > 1%), which accounted for 80.48% of the fungal sequences. Other fungal classes were less abundant in all the soils.
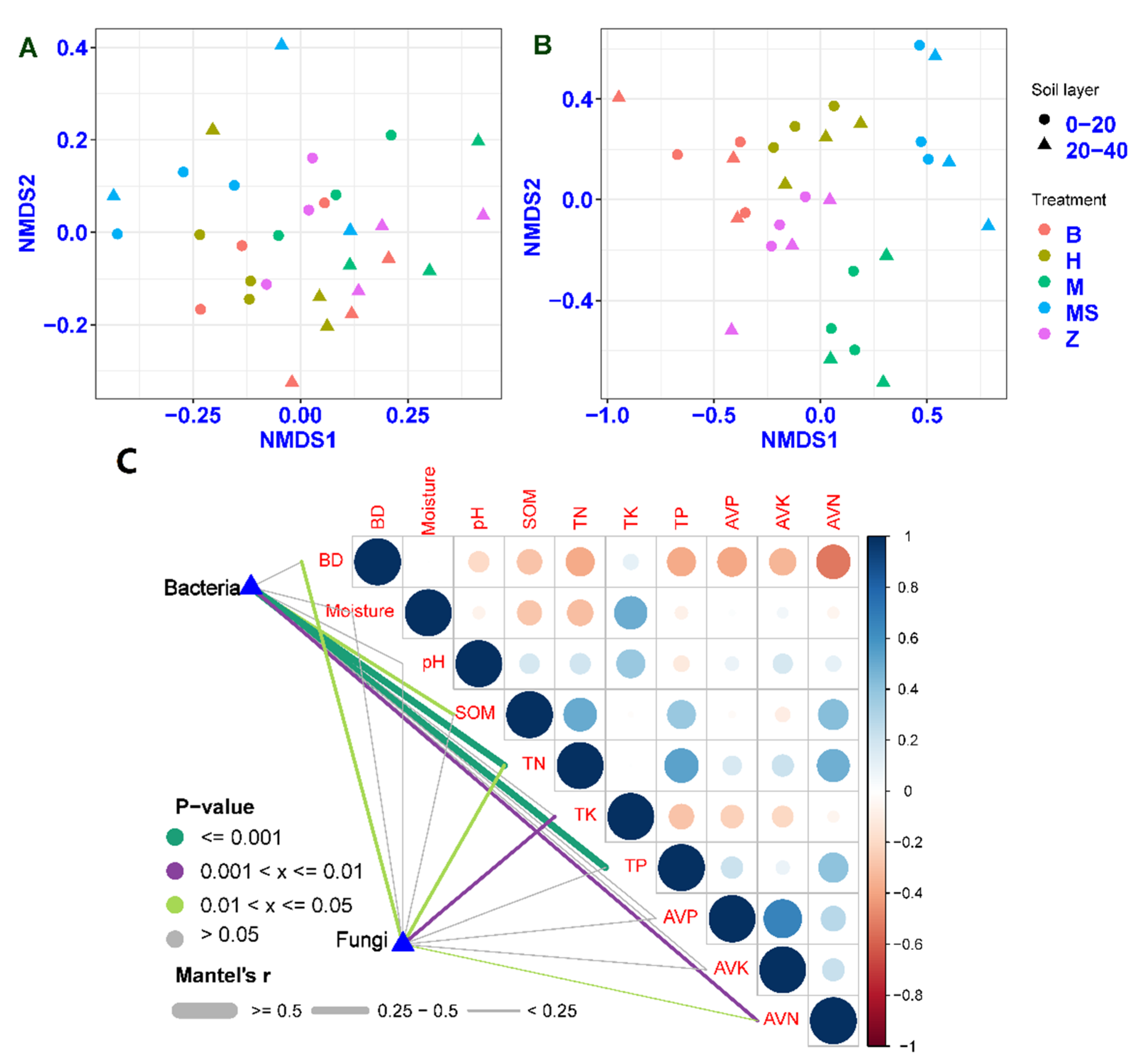
Anosim analysis indicated that the fungal community structure was strongly impacted by the application of sown grass strips (r2 = 0.825, p = 0.001 for 0–20 cm soil layer; r2 = 0.413, p = 0.001 for 20–40 cm soil layer). Soil bacterial community structure was not sensitive to sown grass strips (r2 = 0.179, p = 0.095 for 0–20 cm soil layer; r2 = 0.092, p = 0.203 for 20–40 cm soil layer). Different soil samples were clearly separated by the sown grass types in the NMDS plot (Figure 4A,B). The Mantel test showed that SOM (r = 0.16, p = 0.036), TN (r = 0.45, p = 0.001), TP (r = 0.42, p = 0.001) and AVN (r = 0.23, p = 0.004) had significant effects on the bacterial community structure (Figure 4C). For the fungal community structure, soil BD (r = 0.17, p = 0.033), TN (r = 0.17, p = 0.016), TK (r = 0.21, p = 0.002) and AVN (r = 0.15, p = 0.032) were the main factors.
3.5. Variations in Soil Fungal Ecological Guilds under Different Sown Grass Strips
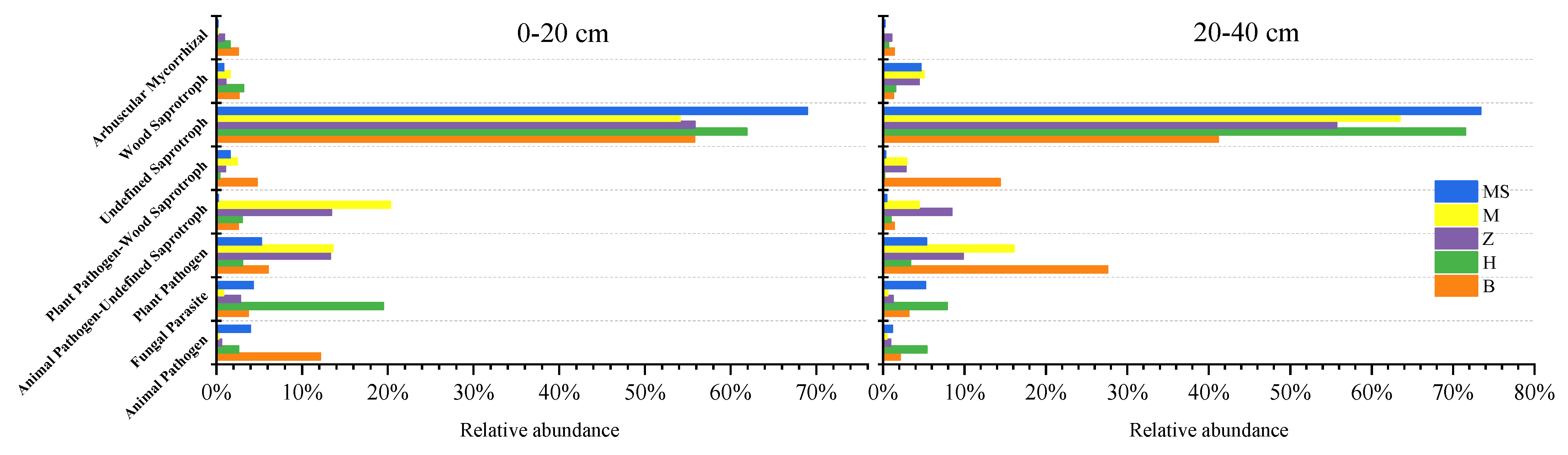
Soil functional fungal groups significantly altered under different cover cropping with different sown grass strips. Cover cropping enhanced the proportion of pathotroph fungi and decreased saprotroph fungi. Cover cropping increased the proportion of Arbuscular Mycorrhizal fungi and wood saprotroph fungi, while cover cropping had no significant effects on the plant pathogen or animal pathogen (Figure 5).
3.6. The Associations between Soil Microbial Community and Diversity and Mango Fruit Yields and Quality
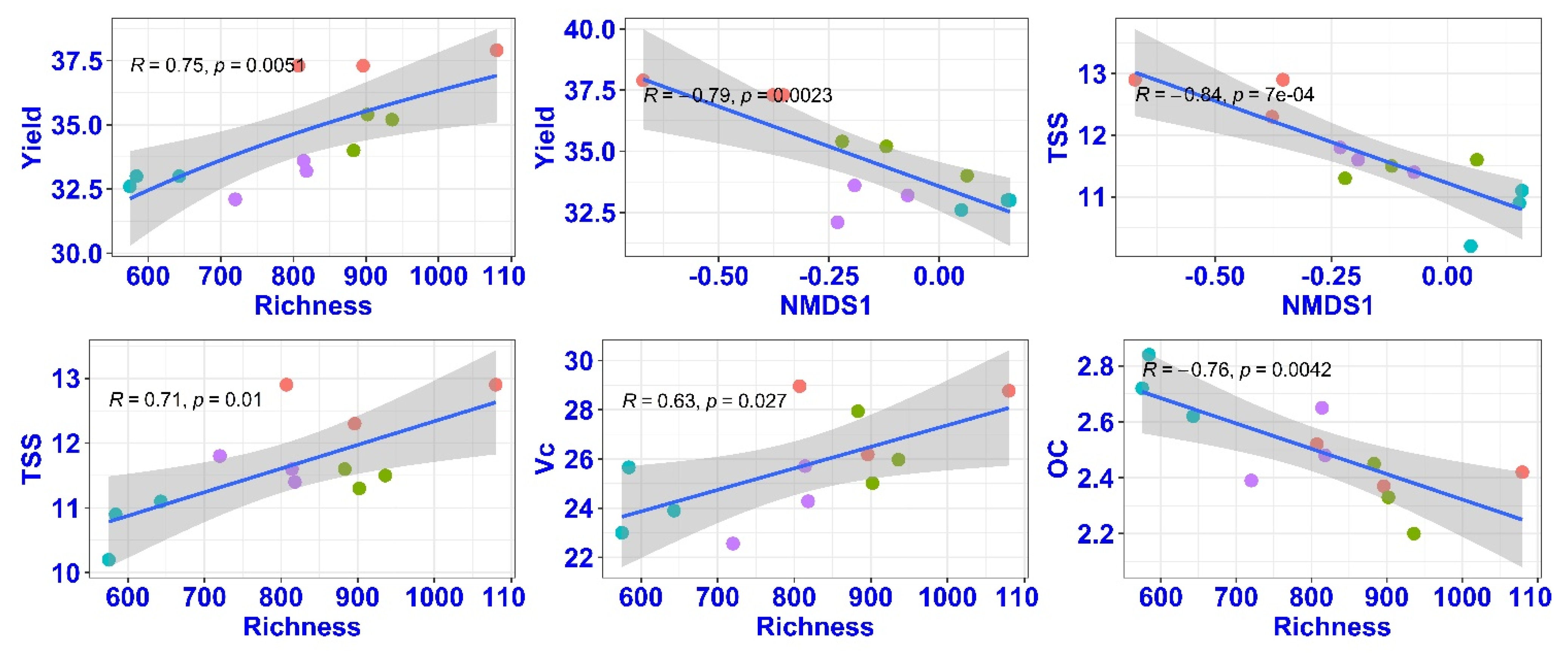
The correlation between fungal diversity (richness) and mango fruit yields was significant (r = 0.75, p < 0.01) (Figure 6). The soil fungal community structure (repressed by NMDS1) was significantly correlated with mango fruit yields (r = −0.79, p < 0.01). These also showed significant correlations between fungal diversity and mango fruit TSS (r = 0.71, p = 0.01) and organic acid (r = −0.76, p < 0.01). However, the soil bacterial diversity had no significant associations with mango fruit yields and TSS (data not shown).
4. Discussion
Cover cropping has been widely used in agriculture systems to improve crop yield and quality as well as soil quality. In our study, the mango yield was significantly enhanced by 3–14% after the application of sown grass strips between rows. Similarly, winter cover cropping improved corn yields [39], and soybean yield significantly increased after the 3-year application with a multispecies mixture of legumes, grasses and Brassica spp [40]. These positive effects of cover cropping in crop yields suggested that cover cropping is an effective method to increase yields. In this study, we also found a significant increase in mango yields with B and H. However, Z had a slight increase in mango yields, suggesting that Z is not suitable to use in mango orchards. The positive effects of B and H on mango yields might be explained by the variations in nutrients and microbial community diversity in soils caused by cover cropping.
A meta-analysis found that global cropland soil carbon changes due to cover cropping, which increases SOC in near-surface soils by an average of 15.5% [2]. However, in our study, we found the sown grass did not enhance SOC in both soil layers. This difference might be explained by the duration of application of the sown grass. Sown grass strips caused the changes in available nutrients, such as AVN and AVP. The growth of sown grass could produce root exudates and litters, which were the main resources of soil nutrients, especially available nutrients. These variations in soil nutrients directly and indirectly influenced soil microbial diversity and community compositions, respectively.
In this study, we found that soil fungal diversity was sensitive to the application of sown grass strips. The introduction of sown grass between the rows enhanced the fungal diversity compared to the control (no sown grass). This might be explained by the higher decomposition ability of litters and roots than bacteria. Fungi are considered the primary decomposers of dead plant biomass in terrestrial ecosystems. The results from the litter-bag decomposition experiments in the field and the laboratory indicated the overwhelming advantage of fungi during the litter decomposition process [41,42,43,44]. Pascoal and Cássio (2004) showed that the contribution of fungi to litter decomposition greatly exceeded that of bacteria [45]. Despite the critic roles of fungi, the roles of bacteria could not be neglected [46], especially during the litter decomposition in which they mainly worked at the later decomposition stage. Therefore, the long-term introduction of sown grass strips might cause variations in soil bacterial diversity and community.
Different types of sown grasses also had different effects on soil fungal diversity. B and H soils had higher soil fungal diversity than Z. These variations might be explained by the quality of plant litter, roots and root exudates, which were the main factors impacting soil fungal diversity and community. For example, high quality litter decomposed faster than the low-quality litter [47,48]. The fast decomposition released a much higher amount of nutrients to the soil and resulted in the fast succession of soil fungi.
Nitrogen was considered as the main resource of soil microbes and strongly affected soil biodiversity and community structure [26,49]. Wang et al. (2018) found that tropical forest soil microbial community composition was shaped by N addition, with the increase in the proportion of arbuscular mycorrhizal fungi [50]. In the present study, we found that soil TN and AVN had significant effects on soil bacterial diversity, suggesting that soil N was the best predictor of the regulation of soil bacterial diversity in the mango orchards.
Soil microorganisms are considered as the main regulator of soil nutrient cycles and net primary production. In this study, we found that soil microbes enhanced mango fruit yields and quality via increasing fungal diversity and alterations in fungal community structure. Soil microbial diversity may improve crop yields through these mechanisms. First, soil biodiversity mediated nutrients available in the soil, which were the main resources of crop plant growth. Many previous studies have suggested that soil biodiversity enhances plant growth and drives crop yields [51,52]. Second, soil microorganisms help to maintain soil health and prevent the invasion of pathogens. Higher soil biodiversity provides higher ecosystem functions, such as net primary production and nutrient cycling. Therefore, soil microbial diversity, especially for fungal diversity, is vital in order to improve mango fruit yields.
5. Conclusions
This field experiment showed that cover cropping with sown grass had significant effects on the soil microbial community in a mango orchard. Fungal diversity was more sensitive than bacterial diversity in response to sown grass. The application of Brachiaria eruciformis (B) had the strongest effects on soil fungal diversity. The increase in fungal diversity suggested that sown grass had positive influences on soil biodiversity. Across soil properties, AVN and TN were the most important predictors affecting soil bacterial communities, while soil nutrients (TN and AVN) were the most important factors in mediating soil fungal communities. These results show that the sown grass strips in a mango orchard regulated the soil microbial community and diversity via soil organic matter and nitrogen, which might be of great significance in mango production and quality and the suitability of mango orchards. Mixtures of sown grasses might be more effective for soil biodiversity and soil function, and operations in mango orchards and future research should be focused on this aspect.
Author Contributions
Conceptualization, W.T. and Z.W.; methodology, Z.W.; data analysis, Q.Z. and Z.W.; investigation, Z.W.; writing—original draft preparation, Q.Z. and Z.W.; writing—review and editing, W.T.; visualization, W.T.; supervision, Z.W.; project administration, Z.W.; funding acquisition, W.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (Nos. 41977023 and 32061123007).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Olson, K.R.; Ebelhar, S.A.; Lang, J.M. Cover crop effects on crop yields and soil organic carbon content. Soil Sci. 2010, 175, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Jian, J.; Du, X.; Reiter, M.S.; Stewart, R.D. A meta-analysis of global cropland soil carbon changes due to cover cropping. Soil Biol. Biochem. 2020, 143, 107735. [Google Scholar] [CrossRef]
- Van Eerd, L.L.; Congreves, K.A.; Hayes, A.; Verhallen, A.; Hooker, D.C. Long-term tillage and crop rotation effects on soil quality, organic carbon, and total nitrogen. Can. J. Soil Sci. 2014, 94, 303–315. [Google Scholar] [CrossRef]
- Bandick, A.K.; Dick, R.P. Field management effects on soil enzyme activities. Soil Biol. Biochem. 1999, 31, 1471–1479. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Xue, K.; Xie, J.; Deng, Y.; Wu, L.; Cheng, X.; Fei, S.; Deng, S.; He, Z.; Van Nostrand, J.D. Microbial mediation of carbon-cycle feedbacks to climate warming. Nat. Clim. Chang. 2012, 2, 106–110. [Google Scholar] [CrossRef]
- Bloor, J.M.G.; Niboyet, A.; Leadley, P.W.; Barthes, L. CO2 and inorganic N supply modify competition for N between co-occurring grass plants, tree seedlings and soil microorganisms. Soil Biol. Biochem. 2009, 41, 544–552. [Google Scholar] [CrossRef]
- Elfstrand, S.; Hedlund, K.; Mårtensson, A. Soil enzyme activities, microbial community composition and function after 47 years of continuous green manuring. Appl. Soil Ecol. 2007, 35, 610–621. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Vasugi, C.; Dinesh, M.; Sekar, K.; Shivashankara, K.; Padmakar, B.; Ravishankar, K. Genetic diversity in unique indigenous mango accessions (Appemidi) of the Western Ghats for certain fruit characteristics. Curr. Sci. 2012, 103, 199–207. [Google Scholar]
- Tharanathan, R.; Yashoda, H.; Prabha, T. Mango (Mangifera indica L.), “The king of fruits”—An overview. Food Rev. Int. 2006, 22, 95–123. [Google Scholar] [CrossRef]
- Neguse, T.B.; Wanzala, F.K.; Ali, W.M.; Owino, W.O.; Mwangi, G.S. Mango (Mangifera indica L.) production practices and constraints in major production regions of Ethiopia. Afr. J. Agric. Res. 2019, 14, 185–196. [Google Scholar]
- Massah, J.; Azadegan, B. Effect of chemical fertilizers on soil compaction and degradation. Agric. Mech. Asia Africa Lat. Am. 2016, 47, 44–50. [Google Scholar]
- Fageria, N. Green manuring in crop production. J. Plant Nutr. 2007, 30, 691–719. [Google Scholar] [CrossRef]
- Baggs, E.; Watson, C.; Rees, R. The fate of nitrogen from incorporated cover crop and green manure residues. Nutr. Cycl. Agroecosyst 2000, 56, 153–163. [Google Scholar] [CrossRef]
- Garcia-Franco, N.; Albaladejo, J.; Almagro, M.; Martínez-Mena, M. Beneficial effects of reduced tillage and green manure on soil aggregation and stabilization of organic carbon in a Mediterranean agroecosystem. Soil Tillage Res. 2015, 153, 66–75. [Google Scholar] [CrossRef]
- Mazzoncini, M.; Sapkota, T.B.; Barberi, P.; Antichi, D.; Risaliti, R. Long-term effect of tillage, nitrogen fertilization and cover crops on soil organic carbon and total nitrogen content. Soil Tillage Res. 2011, 114, 165–174. [Google Scholar] [CrossRef]
- Sainju, U.M.; Singh, B.P.; Whitehead, W.F. Long-term effects of tillage, cover crops, and nitrogen fertilization on organic carbon and nitrogen concentrations in sandy loam soils in Georgia, USA. Soil Tillage Res. 2002, 63, 167–179. [Google Scholar] [CrossRef]
- Duval, M.E.; Galantini, J.A.; Capurro, J.E.; Martinez, J.M. Winter cover crops in soybean monoculture: Effects on soil organic carbon and its fractions. Soil Tillage Res. 2016, 161, 95–105. [Google Scholar] [CrossRef]
- Tolbert, V.R.; Todd, D.E.; Mann, L.K.; Jawdy, C.M.; Mays, D.A.; Malik, R.; Bandaranayake, W.; Houston, A.; Tyler, D.; Pettry, D.E. Changes in soil quality and below-ground carbon storage with conversion of traditional agricultural crop lands to bioenergy crop production. Environ. Pollut. 2002, 116, S97–S106. [Google Scholar] [CrossRef]
- Wang, A.-T.; Wang, Q.; Li, J.; Yuan, G.-L.; Albanese, S.; Petrik, A. Geo-statistical and multivariate analyses of potentially toxic elements’ distribution in the soil of Hainan Island (China): A comparison between the topsoil and subsoil at a regional scale. J. Geochem. Explor. 2019, 197, 48–59. [Google Scholar] [CrossRef]
- Patil, A.S.; Maurer, D.; Feygenberg, O.; Alkan, N. Exploring cold quarantine to mango fruit against fruit fly using artificial ripening. Sci. Rep. 2019, 9, 1948. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Si, B.; Li, H.; Biswas, A. Elucidating controls of the variability of deep soil bulk density. Geoderma 2019, 348, 146–157. [Google Scholar] [CrossRef]
- Ren, H.; Xu, Z.; Huang, J.; Lü, X.; Zeng, D.-H.; Yuan, Z.; Han, X.; Fang, Y. Increased precipitation induces a positive plant-soil feedback in a semi-arid grassland. Plant Soil 2015, 389, 211–223. [Google Scholar] [CrossRef]
- Thomas, R.; Sheard, R.; Moyer, J. Comparison of conventional and automated procedures for nitrogen, phosphorus, and potassium analysis of plant material using a single digestion 1. Agron. J. 1967, 59, 240–243. [Google Scholar] [CrossRef]
- Zeng, Q.C.; An, S.S.; Liu, Y. Soil bacterial community response to vegetation succession after fencing in the grassland of China. Sci. Total Environ. 2017, 609, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berglyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.; An, S. Identifying the Biogeographic Patterns of Rare and Abundant Bacterial Communities Using Different Primer Sets on the Loess Plateau. Microorganisms 2021, 9, 139. [Google Scholar] [CrossRef] [PubMed]
- Fujita, S.-I.; Senda, Y.; Nakaguchi, S.; Hashimoto, T. Multiplex PCR Using Internal Transcribed Spacer 1 and 2 Regions for Rapid Detection and Identification of Yeast Strains. J. Clin. Microbiol. 2001, 39, 3617–3622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Q.; Jia, P.; Wang, Y.; Wang, H.; Li, C.; An, S. The local environment regulates biogeographic patterns of soil fungal communities on the Loess Plateau. Catena 2019, 183, 104220. [Google Scholar] [CrossRef]
- Ni, Y.; Yang, T.; Zhang, K.; Shen, C.; Chu, H. Fungal Communities Along a Small-Scale Elevational Gradient in an Alpine Tundra Are Determined by Soil Carbon Nitrogen Ratios. Front. Microbiol. 2018, 9, 1815. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J.; Abades, S.; Alfaro, F.D.; Bastida, F.; Berhe, A.A.; Cutler, N.A.; Gallardo, A. Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Větrovský, T.; Kohout, P.; Kopecký, M.; Machac, A.; Man, M.; Bahnmann, B.D.; Brabcová, V.; Choi, J.; Meszárošová, L.; Human, Z.R.; et al. A meta-analysis of global fungal distribution reveals climate-driven patterns. Nat. Commun. 2019, 10, 5142. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Guerra, C.A.; Cano-Díaz, C.; Egidi, E.; Wang, J.-T.; Eisenhauer, N.; Singh, B.K.; Maestre, F.T. The proportion of soil-borne pathogens increases with warming at the global scale. Nat. Clim. Chang. 2020, 10, 550–554. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- Goslee, S.C.; Urban, D.L. The ecodist package for dissimilarity-based analysis of ecological data. J. Stat. Softw. 2007, 22, 1–19. [Google Scholar] [CrossRef]
- Kuo, S.; Sainju, U.; Jellum, E. Winter cover cropping influence on nitrogen mineralization, presidedress soil nitrate test, and corn yields. Biol. Fertil. Soils 1996, 22, 310–317. [Google Scholar] [CrossRef]
- Chu, M.; Jagadamma, S.; Walker, F.R.; Eash, N.S.; Buschermohle, M.J.; Duncan, L.A. Effect of multispecies cover crop mixture on soil properties and crop yield. Agric. Environ. Lett. 2017, 2, 170030. [Google Scholar] [CrossRef] [Green Version]
- Stursova, M.; Zifcakova, L.; Leigh, M.B.; Burgess, R.; Baldrian, P. Cellulose utilization in forest litter and soil: Identification of bacterial and fungal decomposers. FEMS Microbiol. Ecol. 2012, 80, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship. Oikos 1997, 79, 439. [Google Scholar] [CrossRef]
- Voříšková, J.; Baldrian, P. Fungal community on decomposing leaf litter undergoes rapid successional changes. ISME J. 2012, 7, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Chauvet, E. Synergistic effects of water temperature and dissolved nutrients on litter decomposition and associated fungi. Glob. Chang. Biol. 2011, 17, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Pascoal, C.; Cássio, F. Contribution of fungi and bacteria to leaf litter decomposition in a polluted river. Appl. Environ. Microbiol. 2004, 70, 5266–5273. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.; Liu, Y.; Zhang, H.; An, S. Fast bacterial succession associated with the decomposition of Quercus wutaishanica litter on the Loess Plateau. Biogeochemistry 2019, 144, 119–131. [Google Scholar] [CrossRef]
- Zhang, P.; Tian, X.; He, X.; Song, F.; Ren, L.; Jiang, P. Effect of litter quality on its decomposition in broadleaf and coniferous forest. Eur. J. Soil Biol. 2008, 44, 392–399. [Google Scholar] [CrossRef]
- Cuchietti, A.; Marcotti, E.; Gurvich, D.E.; Cingolani, A.M.; Harguindeguy, N.P. Leaf litter mixtures and neighbour effects: Low-nitrogen and high-lignin species increase decomposition rate of high-nitrogen and low-lignin neighbours. Appl. Soil Ecol. 2014, 82, 44–51. [Google Scholar] [CrossRef]
- Deng, Q.; Cheng, X.; Hui, D.; Zhang, Q.; Li, M.; Zhang, Q. Soil microbial community and its interaction with soil carbon and nitrogen dynamics following afforestation in central China. Sci. Total. Environ. 2016, 541, 230–237. [Google Scholar] [CrossRef]
- Wang, C.; Lu, X.; Mori, T.; Mao, Q.; Zhou, K.; Zhou, G.; Nie, Y.; Mo, J. Responses of soil microbial community to continuous experimental nitrogen additions for 13 years in a nitrogen-rich tropical forest. Soil Biol. Biochem. 2018, 121, 103–112. [Google Scholar] [CrossRef]
- Saleem, M.; Hu, J.; Jousset, A. More than the sum of its parts: Microbiome biodiversity as a driver of plant growth and soil health. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 145–168. [Google Scholar] [CrossRef]
- Rillig, M.C.; Lehmann, A.; Lehmann, J.; Camenzind, T.; Rauh, C. Soil Biodiversity Effects from Field to Fork. Trends Plant Sci. 2018, 23, 17–24. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Experimental plot and soil sampling protocol: B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M, no-till + herbicide. In the control, soils were collected between the rows (M) and the drip line of mango trees (MS). Other soils (H, Z and B) with cover crops were collected between the rows.
Figure 1.
Experimental plot and soil sampling protocol: B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M, no-till + herbicide. In the control, soils were collected between the rows (M) and the drip line of mango trees (MS). Other soils (H, Z and B) with cover crops were collected between the rows.

Figure 2.
The associations between soil bacterial diversity and soil properties. TK, total potassium; TN, total nitrogen; TP, total phosphorus; AVP, available phosphorus; AVK, available potassium; AVN, available nitrogen. B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M and MS, no-till + herbicide.
Figure 2.
The associations between soil bacterial diversity and soil properties. TK, total potassium; TN, total nitrogen; TP, total phosphorus; AVP, available phosphorus; AVK, available potassium; AVN, available nitrogen. B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M and MS, no-till + herbicide.

Figure 3.
The community compositions of soil bacteria (A) and fungi (B) at phylum level. B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M and MS, no-till + herbicide.
Figure 3.
The community compositions of soil bacteria (A) and fungi (B) at phylum level. B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M and MS, no-till + herbicide.

Figure 4.
The community structure of soil bacteria (A) and fungi (B), and the associations between community structure and soil properties (C). B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M and MS, no-till + herbicide. In the control (M and MS), soils were collected between the rows (M) and the drip line of mango trees (MS).
Figure 4.
The community structure of soil bacteria (A) and fungi (B), and the associations between community structure and soil properties (C). B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M and MS, no-till + herbicide. In the control (M and MS), soils were collected between the rows (M) and the drip line of mango trees (MS).

Figure 5.
The functional groups of soil fungi under different cover cropping treatments. B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M, no-till + herbicide. In the control, soils were collected between the rows (M) and the drip line of mango trees (MS).
Figure 5.
The functional groups of soil fungi under different cover cropping treatments. B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M, no-till + herbicide. In the control, soils were collected between the rows (M) and the drip line of mango trees (MS).

Figure 6.
The associations between fungal community characteristics and mango fruit yield and quality under different cover cropping treatments. TSS, total soluble solids content; Vc, vitamin C; OC, organic acid. NMDS1 represented the community structure of soil fungi, and Richness indicated fungal diversity.
Figure 6.
The associations between fungal community characteristics and mango fruit yield and quality under different cover cropping treatments. TSS, total soluble solids content; Vc, vitamin C; OC, organic acid. NMDS1 represented the community structure of soil fungi, and Richness indicated fungal diversity.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The characteristics of soil properties of mango soils under different sown grass strips.
| Treatment | Soil layers | BD g/cm3 | Moisture % | TK% | TN% | TP% | pH | SOM % | AVP mg/kg | AVK mg/kg | AVN mg/kg |
|---|---|---|---|---|---|---|---|---|---|---|---|
| B20 | 0–20 cm | 1.48 ± 0.06 | 6.49 ± 1.19 | 3.44 ± 0.27 | 0.07 ± 0.01 | 0.01 ± 0 | 5.76 ± 0.14 | 1.35 ± 0.2 | 5.7 ± 1.59 | 97.43 ± 16.37 | 49.47 ± 7.68 |
| H20 | 1.48 ± 0.01 | 6.02 ± 0.14 | 3.26 ± 0.53 | 0.07 ± 0 | 0.02 ± 0 | 5.55 ± 0.06 | 1.12 ± 0.01 | 6.32 ± 1.5 | 90.33 ± 11.77 | 38.27 ± 2.14 | |
| Z20 | 1.45 ± 0.06 | 5.86 ± 0.66 | 3.49 ± 0.05 | 0.06 ± 0.01 | 0.02 ± 0 | 5.51 ± 0.09 | 1.36 ± 0.06 | 11.95 ± 6.12 | 81.07 ± 22.86 | 37.45 ± 5.85 | |
| M20 | 1.51 ± 0.04 | 3.75 ± 0.8 | 2.25 ± 0.13 | 0.07 ± 0 | 0.01 ± 0 | 5.49 ± 0.04 | 1.28 ± 0.18 | 7.15 ± 2.15 | 101.33 ± 28.45 | 34.07 ± 7.01 | |
| MS20 | 1.38 ± 0.06 | 5.01 ± 1.33 | 2.07 ± 0.3 | 0.07 ± 0.01 | 0.05 ± 0.03 | 5.55 ± 0.36 | 1.44 ± 0.11 | 30.03 ± 18.81 | 104.88 ± 11.55 | 51.57 ± 5.06 | |
| B40 | 20–40 cm | 1.46 ± 0.05 | 8.31 ± 1.69 | 3.58 ± 0.19 | 0.05 ± 0.01 | 0.02 ± 0 | 5.71 ± 0.17 | 1.16 ± 0.16 | 6.56 ± 0.3 | 93.18 ± 21.85 | 34.88 ± 6.95 |
| H40 | 1.53 ± 0.13 | 7.55 ± 1.58 | 2.9 ± 0.53 | 0.06 ± 0.01 | 0.02 ± 0.01 | 5.48 ± 0.13 | 0.96 ± 0.04 | 5.3 ± 0.73 | 91 ± 6.58 | 40.6 ± 9.15 | |
| Z40 | 1.55 ± 0.01 | 7.99 ± 0.8 | 3.32 ± 0.13 | 0.05 ± 0 | 0.02 ± 0 | 5.44 ± 0.14 | 1.19 ± 0.31 | 11.66 ± 5.5 | 86.07 ± 37.41 | 35.7 ± 1.4 | |
| M40 | 1.5 ± 0.02 | 7.56 ± 1.37 | 2.35 ± 0.17 | 0.05 ± 0 | 0.02 ± 0 | 5.37 ± 0.02 | 1.06 ± 0.27 | 5.57 ± 2.1 | 76.23 ± 8.73 | 29.63 ± 2.65 | |
| MS40 | 1.4 ± 0.04 | 7.08 ± 1.08 | 2.56 ± 0.31 | 0.07 ± 0.02 | 0.03 ± 0.01 | 5.4 ± 0.17 | 1.19 ± 0.22 | 42.71 ± 49.83 | 122.1 ± 45.21 | 46.43 ± 2.46 |
BD, soil bulk density; TK, total potassium; TN, total nitrogen; TP, total phosphorus; SOM, soil organic matter; AVP, available phosphorus; AVK, available potassium; AVN, available nitrogen.
Table 2.
The yield and fruit quality of mango under different sown grass trips.
| Treatment | Yield (kg/ha) | TSS (%) | Vc (mg/100 g) | Organic Acid (g/kg) |
|---|---|---|---|---|
| B | 18,736 ± 202 a | 12.7 ± 0.16 a | 27.97 ± 1.22 a | 2.44 ± 0.13 b |
| H | 17,424 ± 362 b | 11.47 ± 0.16 b | 26.31 ± 0.62 ab | 2.33 ± 0.07 b |
| Z | 16,488 ± 298 c | 11.59 ± 0.3 b | 24.18 ± 1.93 b | 2.51 ± 0.11 ab |
| M | 16,436 ± 189 c | 10.75 ± 0.21 c | 24.19 ± 0.49 b | 2.93 ± 0.06 a |
TSS, total soluble solids content; Vc, vitamin C. Different letters indicate significant differences under different sown grass trips. B, Brachiaria eruciformis; Z, Stylosanthes guianensis; H, Butterfly pea; M, no-till + herbicide. Different letter indicates significant differences between different sown grass trips.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wei, Z.; Zeng, Q.; Tan, W. Cover Cropping Impacts Soil Microbial Communities and Functions in Mango Orchards. Agriculture 2021, 11, 343. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11040343
AMA Style
Wei Z, Zeng Q, Tan W. Cover Cropping Impacts Soil Microbial Communities and Functions in Mango Orchards. Agriculture. 2021; 11(4):343. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11040343
Chicago/Turabian StyleWei, Zhiyuan, Quanchao Zeng, and Wenfeng Tan. 2021. "Cover Cropping Impacts Soil Microbial Communities and Functions in Mango Orchards" Agriculture 11, no. 4: 343. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11040343
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.