
Nitrogen and Phosphorus Addition to Soil Improves Seed Yield, Foliar Stomatal Conductance, and the Photosynthetic Response of Rapeseed (Brassica napus L.)

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.2. Physiological and Biochemical Parameters
2.3. Seed Yield Measurements
2.4. Statistical Analysis
3. Results
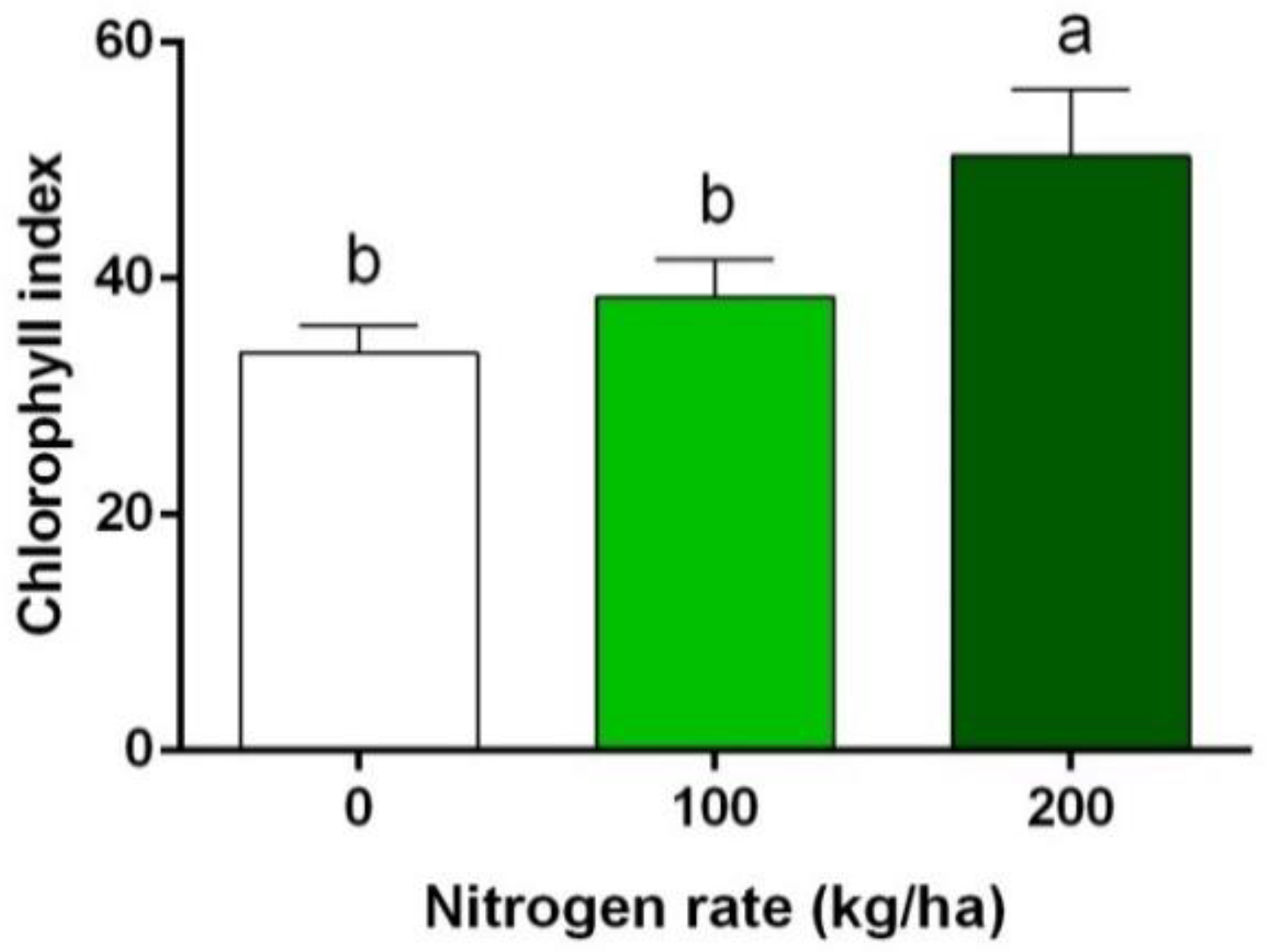
3.1. Chlorophyll
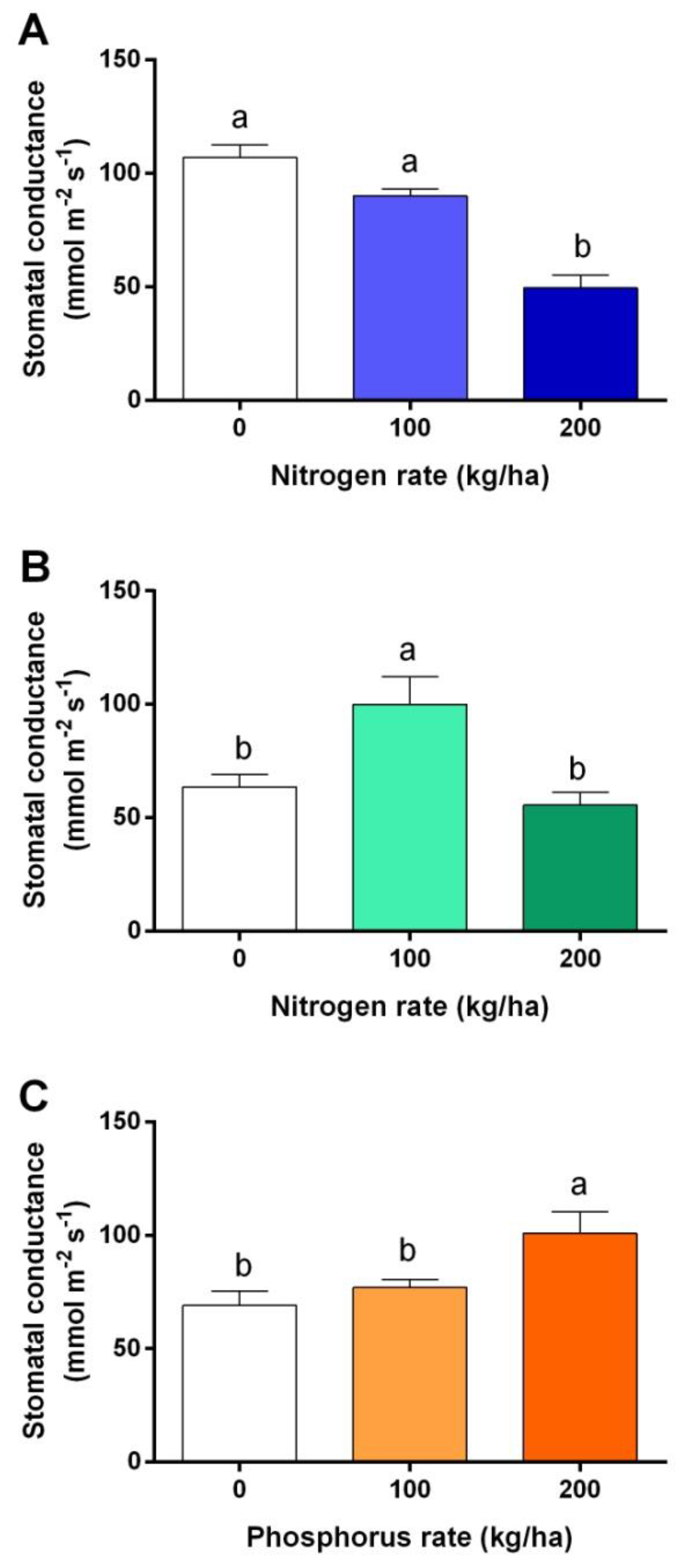
3.2. Stomatal Conductance
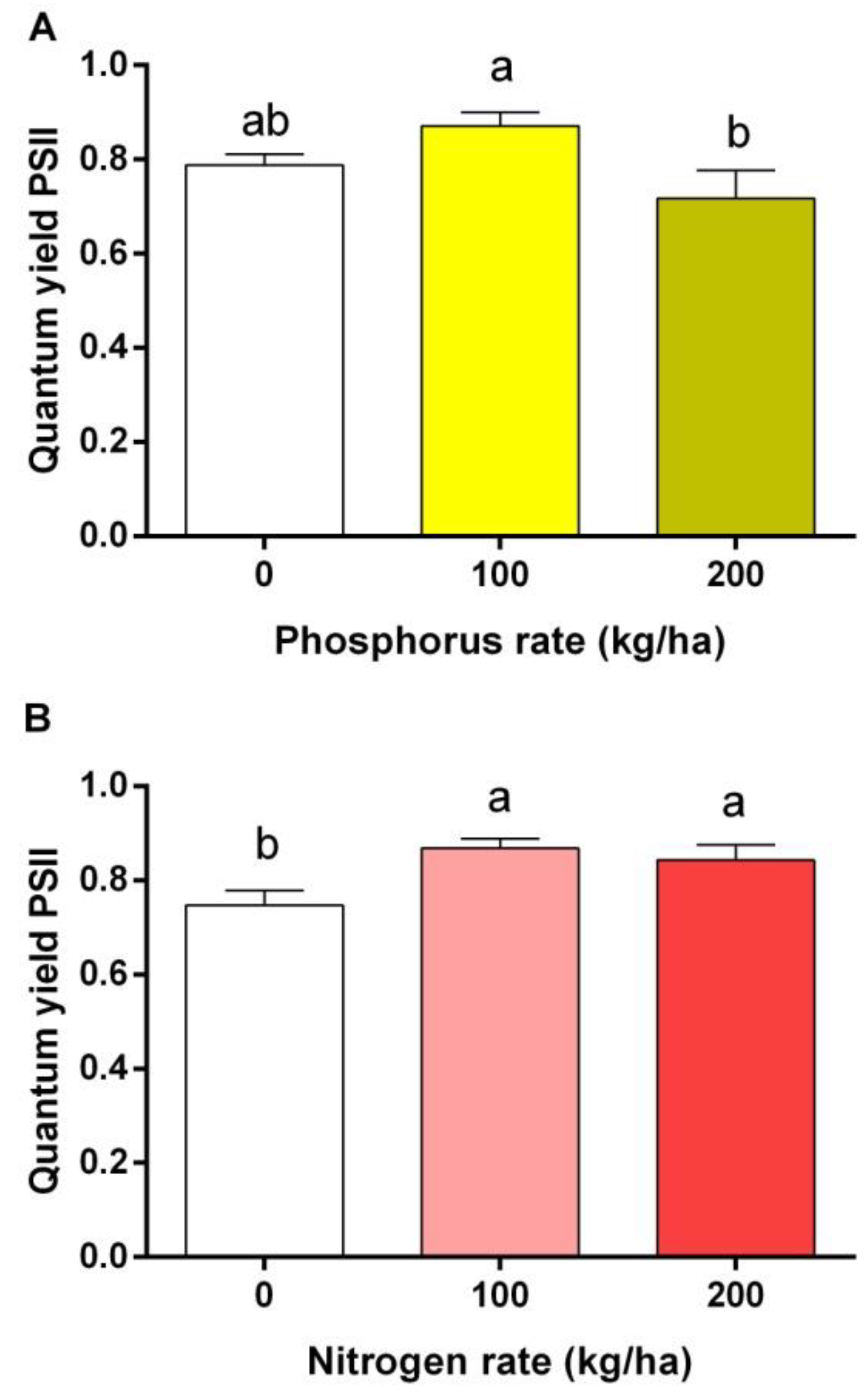
3.3. Quantum Yield of Photosystem II
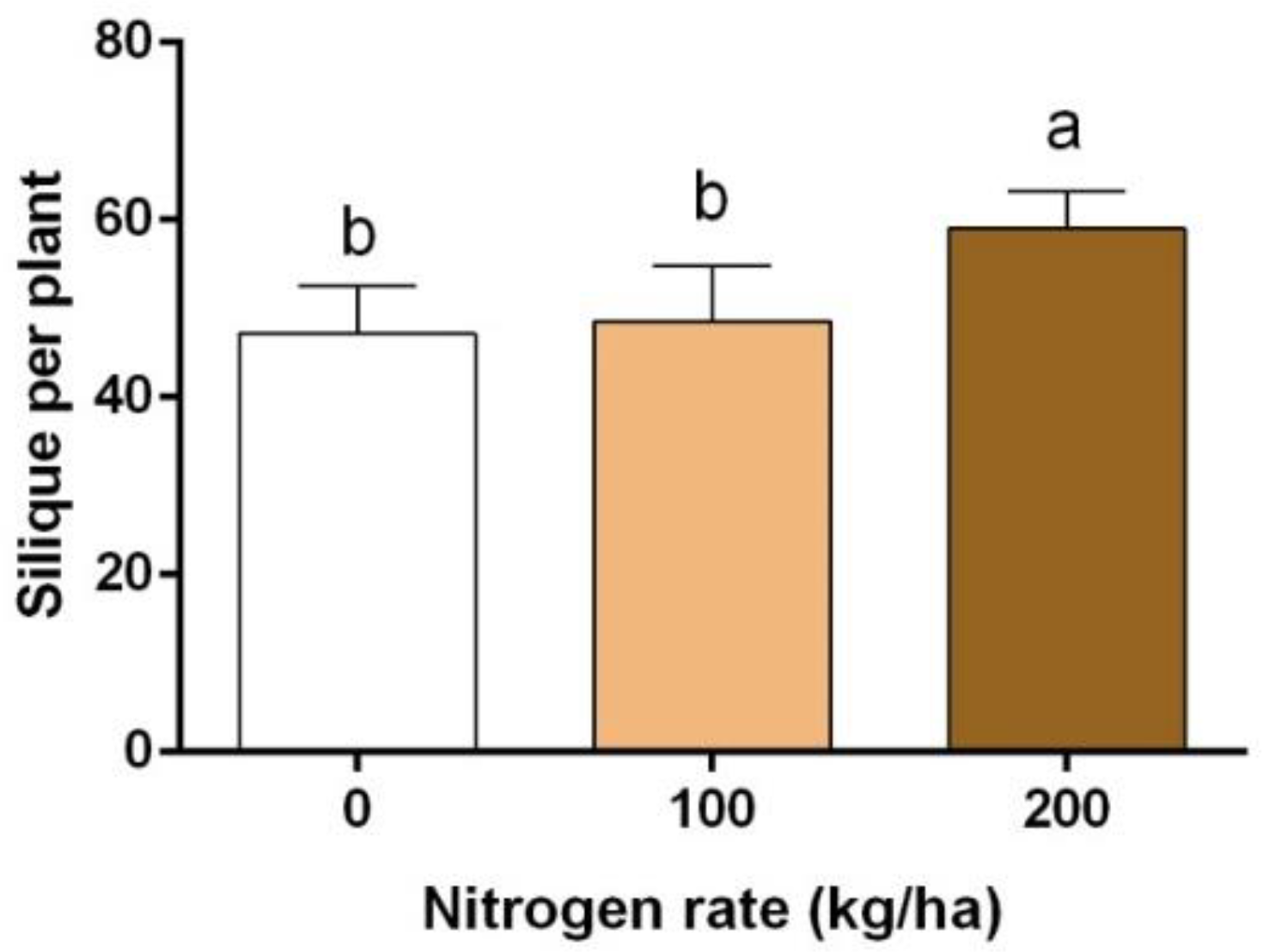
3.4. Yield and Seed Yield Components
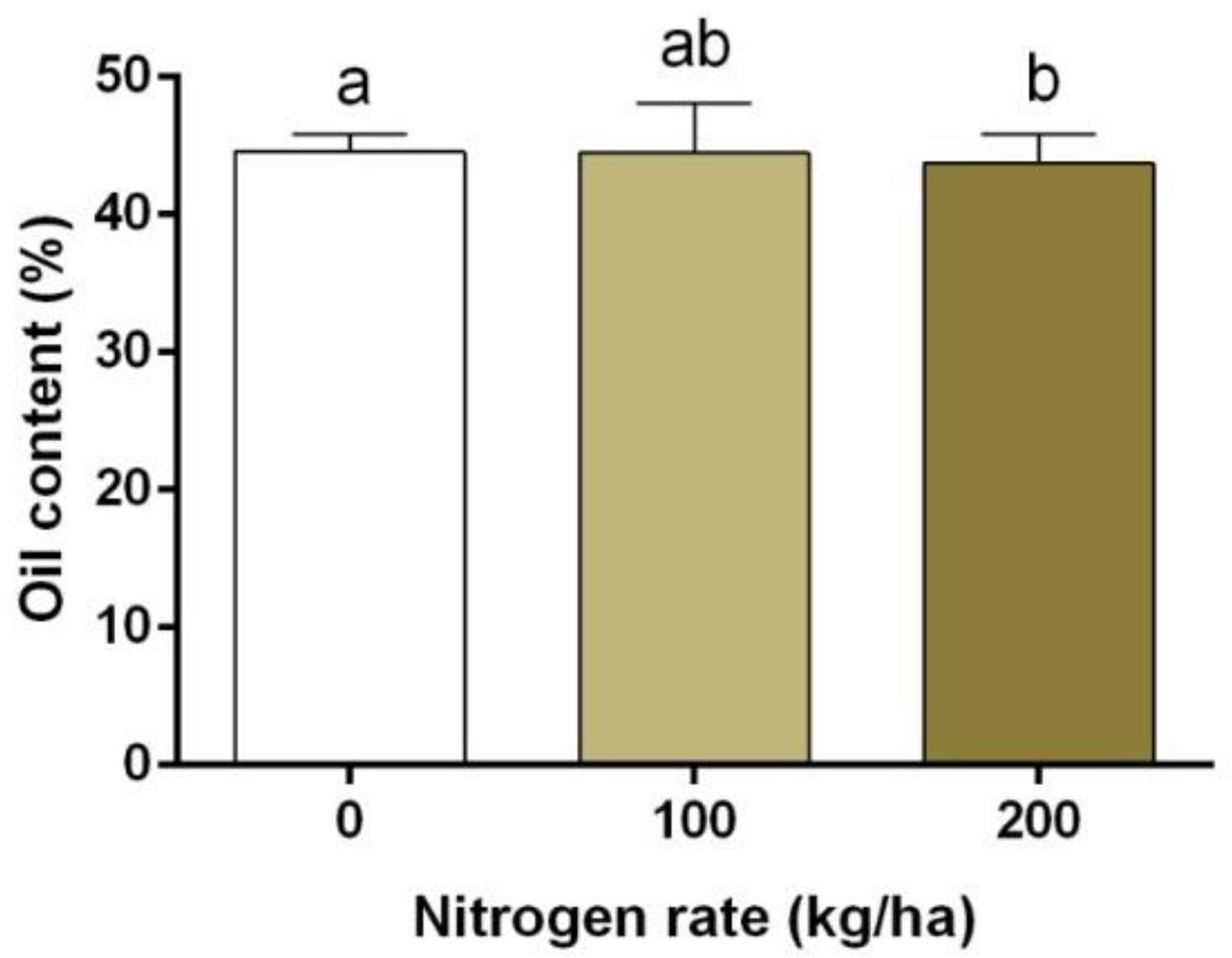
3.5. Oil Content and Oil Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aghajanlou, F.; Mirdavoudi, H.; Shojaee, M.; Mac Sweeney, E.; Mastinu, A.; Moradi, P. Rangeland Management and Ecological Adaptation Analysis Model for Astragalus curvirostris Boiss. Horticulturae 2021, 7, 67. [Google Scholar] [CrossRef]
- Zuo, Q.S.; Zhou, G.S.; Yang, S.F.; Yang, Y.; Wu, L.R.; Leng, S.H.; Yang, G.; Wu, J.S. Effects of nitrogen rate and genotype on seed protein and amino acid content in canola. J. Agric. Sci. 2015, 154, 438–455. [Google Scholar] [CrossRef]
- Kumar, A.; Memo, M.; Mastinu, A. Plant behaviour: An evolutionary response to the environment? Plant Biol. 2020, 22, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Premoli, M.; Aria, F.; Bonini, S.A.; Maccarinelli, G.; Gianoncelli, A.; Memo, M.; Mastinu, A. Cannabimimetic plants: Are they new cannabinoidergic modulators? Planta 2019, 249, 1681–1694. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, A.; Moradi, P.; Mastinu, A. Variation in Terpene Profiles of Thymus vulgaris in Water Deficit Stress Response. Molecules 2020, 25, 1091. [Google Scholar] [CrossRef] [Green Version]
- Plaxton, W.; Lambers, H. Phosphorus Metabolism in Plants; pp. 1 Online Resource (476 pages); Available online: https://0-onlinelibrary-wiley-com.brum.beds.ac.uk/doi/book/10.1002/9781118958841 (accessed on 14 April 2015).
- Rad, S.V.; Valadabadi, S.A.R.; Pouryousef, M.; Saifzadeh, S.; Zakrin, H.R.; Mastinu, A. Quantitative and Qualitative Evaluation of Sorghum bicolor L. under Intercropping with Legumes and Different Weed Control Methods. Horticulturae 2020, 6, 78. [Google Scholar] [CrossRef]
- Reza Yousefi, A.; Rashidi, S.; Moradi, P.; Mastinu, A. Germination and Seedling Growth Responses of Zygophyllum fabago, Salsola kali L. and Atriplex canescens to PEG-Induced Drought Stress. Environments 2020, 7, 107. [Google Scholar] [CrossRef]
- Gupta, A.K.; Rather, M.A.; Kumar Jha, A.; Shashank, A.; Singhal, S.; Sharma, M.; Pathak, U.; Sharma, D.; Mastinu, A. Artocarpus lakoocha Roxb. and Artocarpus heterophyllus Lam. Flowers: New Sources of Bioactive Compounds. Plants 2020, 9, 1329. [Google Scholar] [CrossRef]
- Lazzari, P.; Pau, A.; Tambaro, S.; Asproni, B.; Ruiu, S.; Pinna, G.; Mastinu, A.; Curzu, M.M.; Reali, R.; Bottazzi, M.E.; et al. Synthesis and pharmacological evaluation of novel 4-alkyl-5-thien-2’-yl pyrazole carboxamides. Cent. Nerv. Syst. Agents Med. Chem. 2012, 12, 254–276. [Google Scholar] [CrossRef]
- Mastinu, A.; Bonini, S.A.; Premoli, M.; Maccarinelli, G.; Mac Sweeney, E.; Zhang, L.; Lucini, L.; Memo, M. Protective Effects of Gynostemma pentaphyllum (var. Ginpent) against Lipopolysaccharide-Induced Inflammation and Motor Alteration in Mice. Molecules 2021, 26, 570. [Google Scholar] [CrossRef]
- Ma, B.L.; Zheng, Z.M.; Navabi, A. Relationship between plant nitrogen and phosphorus accumulations in a canola crop as affected by nitrogen management under ample phosphorus supply conditions. Can. J. Plant Sci. 2016, 96, 853–866. [Google Scholar] [CrossRef]
- Tabak, M.; Lepiarczyk, A.; Filipek-Mazur, B.; Lisowska, A. Efficiency of Nitrogen Fertilization of Winter Wheat Depending on Sulfur Fertilization. Agronomy 2020, 10, 1304. [Google Scholar] [CrossRef]
- Grant, C.A.; Bailey, L.D. Fertility Management in Canola Production. Can. J. Plant Sci. 1993, 73, 651–670. [Google Scholar] [CrossRef]
- Brennan, R.F.; Bolland, M.D.A. Comparing the nitrogen and phosphorus requirements of canola and wheat for grain yield and quality. Crop Pasture Sci. 2009, 60, 566. [Google Scholar] [CrossRef]
- Jin, Z.; Chen, C.; Chen, X.; Hopkins, I.; Zhang, X.; Han, Z.; Jiang, F.; Billy, G. The crucial factors of soil fertility and rapeseed yield—A five year field trial with biochar addition in upland red soil, China. Sci. Total Environ. 2019, 649, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Sieling, K.; Böttcher, U.; Kage, H. Effect of Sowing Method and N Application on Seed Yield and N Use Efficiency of Winter Oilseed Rape. Agronomy 2017, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Brennan, R.F.; Bolland, M.D.A. Effect of fertiliser phosphorus and nitrogen on the concentrations of oil and protein in grain and the grain yield of canola (Brassica napus L.) grown in south-western Australia. Aust. J. Exp. Agric. 2007, 47, 984. [Google Scholar] [CrossRef]
- Ding, L.; Wang, K.J.; Jiang, G.M.; Biswas, D.K.; Xu, H.; Li, L.F.; Li, Y.H. Effects of nitrogen deficiency on photosynthetic traits of maize hybrids released in different years. Ann. Bot. 2005, 96, 925–930. [Google Scholar] [CrossRef] [Green Version]
- Naservafaei, S.; Sohrabi, Y.; Moradi, P.; Mac Sweeney, E.; Mastinu, A. Biological Response of Lallemantia iberica to Brassinolide Treatment under Different Watering Conditions. Plants 2021, 10, 496. [Google Scholar] [CrossRef]
- Hossain, M.D.; Hanafi Musa, M.; Talib, J.; Jol, H. Effects of Nitrogen, Phosphorus and Potassium Levels on Kenaf (Hibiscus cannabinus L.) Growth and Photosynthesis under Nutrient Solution. J. Agric. Sci. 2010, 2. [Google Scholar] [CrossRef] [Green Version]
- Brennan, R.F.; Mason, M.G.; Walton, G.H. Effect of nitrogen fertilizer on the concentrations of oil and protein in Canola (Brassica napus) seed. J. Plant Nutr. 2000, 23, 339–348. [Google Scholar] [CrossRef]
- Jackson, G.D. Effects of Nitrogen and Sulfur on Canola Yield and Nutrient Uptake. Agron. J. 2000, 92, 644–649. [Google Scholar] [CrossRef]
- Cheema, M.A.; Malik, M.A.; Hussain, A.; Shah, S.H.; Basra, S.M.A. Effects of Time and Rate of Nitrogen and Phosphorus Application on the Growth and the Seed and Oil Yields of Canola (Brassica napus L.). J. Agron. Crop Sci. 2001, 186, 103–110. [Google Scholar] [CrossRef]
- Hocking, P.J.; Stapper, M. Effects of sowing time and nitrogen fertiliser on canola and wheat, and nitrogen fertiliser on Indian mustard. I. Dry matter production, grain yield, and yield components. Aust. J. Agric. Res. 2001, 52, 623. [Google Scholar] [CrossRef]
- Yuan, P.; Ding, G.D.; Cai, H.M.; Jin, K.M.; Broadley, M.R.; Xu, F.S.; Shi, L. A novel Brassica-rhizotron system to unravel the dynamic changes in root system architecture of oilseed rape under phosphorus deficiency. Ann. Bot. 2016, 118, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wan, J.; Liu, L.; Ye, M.; Jiang, X. Metagenomics reveals functional profiling of microbial communities in OCP contaminated sites with rapeseed oil and tartaric acid biostimulation. J. Environ. Manag. 2021, 289, 112515. [Google Scholar] [CrossRef]
- Tian, C.; Zhou, X.; Liu, Q.; Peng, J.W.; Wang, W.M.; Zhang, Z.H.; Yang, Y.; Song, H.X.; Guan, C.Y. Effects of a controlled-release fertilizer on yield, nutrient uptake, and fertilizer usage efficiency in early ripening rapeseed (Brassica napus L.). J. Zhejiang Univ. Sci. B 2016, 17, 775–786. [Google Scholar] [CrossRef] [Green Version]
- Hammac, W.A.; Maaz, T.M.; Koenig, R.T.; Burke, I.C.; Pan, W.L. Water and Temperature Stresses Impact Canola (Brassica napus L.) Fatty Acid, Protein, and Yield over Nitrogen and Sulfur. J. Agric. Food Chem. 2017, 65, 10429–10438. [Google Scholar] [CrossRef]
- Pan, W.L.; Young, F.L.; Maaz, T.M.; Huggins, D.R. Canola integration into semi-arid wheat cropping systems of the inland Pacific Northwestern USA. Crop Pasture Sci. 2016, 67, 253. [Google Scholar] [CrossRef]
- Hak, R.; Rinderlezimmer, U.; Lichtenthaler, H.K.; Natr, L. Chlorophyll-a Fluorescence Signatures of Nitrogen Deficient Barley Leaves. Photosynthetica 1993, 28, 151–159. [Google Scholar]
- Tóth, V.R.; Mészáros, I.; Veres, S.; Nagy, J. Effects of the available nitrogen on the photosynthetic activity and xanthophyll cycle pool of maize in field. J. Plant Physiol. 2002, 159, 627–634. [Google Scholar] [CrossRef]
- Huang, Z.A.; Jiang, D.A.; Yang, Y.; Sun, J.W.; Jin, S.H. Effects of Nitrogen Deficiency on Gas Exchange, Chlorophyll Fluorescence, and Antioxidant Enzymes in Leaves of Rice Plants. Photosynthetica 2004, 42, 357–364. [Google Scholar] [CrossRef]
- Lima, J.D.; Mosquim, P.R.; Matta, F.M. Leaf Gas Exchange and Chlorophyll Fluorescence Parameters in Phaseolus Vulgaris as Affected by Nitrogen and Phosphorus Deficiency. Photosynthetica 1999, 37, 113–121. [Google Scholar] [CrossRef]
- Verhoeven, A.S.; Demmig-Adams, B.; Adams, I.W. Enhanced Employment of the Xanthophyll Cycle and Thermal Energy Dissipation in Spinach Exposed to High Light and N Stress. Plant Physiol. 1997, 113, 817–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khamis, S.; Lamaze, T.; Lemoine, Y.; Foyer, C. Adaptation of the Photosynthetic Apparatus in Maize Leaves as a Result of Nitrogen Limitation: Relationships between Electron Transport and Carbon Assimilation. Plant Physiol. 1990, 94, 1436–1443. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Garvin, D.F.; Kochian, L.V. Nitrate-induced genes in tomato roots. Array analysis reveals novel genes that may play a role in nitrogen nutrition. Plant Physiol. 2001, 127, 345–359. [Google Scholar] [CrossRef] [Green Version]
- Broadley, M.R.; Escobar-Gutiérrez, A.J.; Burns, A.; Burns, I.G. Nitrogen-limited growth of lettuce is associated with lower stomatal conductance. New Phytol. 2001, 152, 97–106. [Google Scholar] [CrossRef]
- Trapani, N.; Hall, A.J.; Weber, M. Effects of constant and variable nitrogen supply on sunflower (Helianthus annus L.) leaf cell number and size. Ann. Bot. 1999, 84, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.R.; Matcha, S.K. Quantifying nitrogen effects on castor bean (Ricinus communis L.) development, growth, and photosynthesis. Ind. Crop Prod. 2010, 31, 185–191. [Google Scholar] [CrossRef]
- Xu, A.; Li, L.; Xie, J.; Wang, X.; Coulter, J.A.; Liu, C.; Wang, L. Effect of Long-Term Nitrogen Addition on Wheat Yield, Nitrogen Use Efficiency, and Residual Soil Nitrate in a Semiarid Area of the Loess Plateau of China. Sustainability 2020, 12, 1735. [Google Scholar] [CrossRef] [Green Version]
- Farooq, S.; Khalofah, A.; Khan, M.I.; Arif, M.; Hussain, A.; Ullah, R.; Irfan, M.; Mahpara, S.; Shah, R.U.; Ansari, M.J.; et al. Deep placement of nitrogen fertilizer improves yield, nitrogen use efficiency and economic returns of transplanted fine rice. PLoS ONE 2021, 16, e0247529. [Google Scholar] [CrossRef]
- Pasley, H.R.; Cairns, J.E.; Camberato, J.J.; Vyn, T.J. Nitrogen fertilizer rate increases plant uptake and soil availability of essential nutrients in continuous maize production in Kenya and Zimbabwe. Nutr. Cycl. Agroecosystems 2019, 115, 373–389. [Google Scholar] [CrossRef] [Green Version]
- Mahama, G.Y.; Prasad, P.V.V.; Mengel, D.B.; Tesso, T.T. Influence of Nitrogen Fertilizer on Growth and Yield of Grain Sorghum Hybrids and Inbred Lines. Agron. J. 2014, 106, 1623–1630. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Jia, Y.; Chen, H.; Zhang, L.; Yang, J.; Zhang, J.; Hu, X.; Ye, X.; Li, Y.; Zhou, Y. Growth, photosynthesis, and nutrient uptake in wheat are affected by differences in nitrogen levels and forms and potassium supply. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, R.F.; Bolland, M.D.A. Influence of potassium and nitrogen fertiliser on yield, oil and protein concentration of canola (Brassica napus L.) grain harvested in south- western Australia. Aust. J. Exp. Agric. 2007, 47, 976–983. [Google Scholar] [CrossRef]
- Ahmad, G.; Ali Nasrollahzadeh, A. The effect of application using nitragin and nitroxin biofertilizers on reduce the use of nitrogen chemical fertilizer in sunflower cultivation (Helianthus annuus L.). Environ. Conserv. J. 2018, 19, 39–46. [Google Scholar] [CrossRef]
- Ordonez, C.; Tejada, M.; Benitez, C.; Gonzalez, J.L. Characterization of a phosphorus-potassium solution obtained during a protein concentrate process from sunflower flour. Application on rye-grass. Bioresour. Technol. 2006, 97, 522–528. [Google Scholar] [CrossRef]
- Vera, C.L.; Woods, S.M.; Raney, J.R. Seeding rate and row spacing effect on weed competition, yield and quality of hemp in the Parkland region of Saskatchewan. Can. J. Plant Sci. 2006, 86, 911–915. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sand | Silt | Clay | pH | EC | O.C | K | P | N |
|---|---|---|---|---|---|---|---|---|
| (%) | (%) | (%) | ds/m | (%) | ppm | ppm | (%) | |
| 14 | 51 | 35 | 7.8 | 0.59 | 0.84 | 152 | 8.5 | 0.12 |
| Mean Square | |||||||
|---|---|---|---|---|---|---|---|
| Variables | df | Chl. Content (First Stage) mg g−1 | Chl. Content (Second Stage) mg g−1 | Stomatal Conductance (First Stage) mmol−2 s−1 | Stomatal Conductance (Second Stage) mmol−2 s−1 | Quantum Yield PSII (First Stage) Fv/Fm | Quantum Yield PSII (Second Stage) Fv/Fm |
| Repeat | 2 | 94.18 | 0.048 | 9326.8 | 1286.8 | 0.044 | 0.022 |
| Nitrogen (N) | 2 | 664.6 ** | 36.11 ns | 7851.9 ** | 5037.9 * | 0.032 ns | 0.037 * |
| Phosphorus (P) | 2 | 15.81 ns | 94.75 ns | 2457.8 ** | 267.8 ns | 0.059 * | 0.010 ns |
| N*P | 4 | 46.2 ns | 198.03 ** | 1270.6 ns | 1327.6 ns | 0.005 ns | 0.010 ns |
| Subplot error | 16 | 52.3 | 32.42 | 477.5 | 1245.5 | 0.014 | 0.008 |
| CV | 17.2 | 15.3 | 26.5 | 18.9 | 15.2 | 10.6 | |
| Mean Square | ||||||||
|---|---|---|---|---|---|---|---|---|
| Variables | df | Seed Yield (kg/ha) | Oil Yield (kg/ha) | Oil Content (%) | Seed per Silique | Siliques per Plant | 1000 Seed Weight (g) | Harvest Index (%) |
| Repeat | 2 | 2,107,874 | 421,173 | 0.712 | 5.53 | 607.7 | 0.147 | 86.5 |
| Nitrogen (N) | 2 | 3,939,121 ** | 732,087 ** | 2.89 * | 2.61 ns | 377.4 * | 0.218 ns | 45.9 ns |
| Phosphorus (P) | 2 | 1,106,963 ns | 210,125 ns | 0.161 ns | 13.46 ns | 295.5 ns | 0.713 ns | 16.3 ns |
| N*P | 4 | 1,903,944 * | 405,293 * | 1.21 ns | 11.08 ns | 109.8 ns | 1.055 * | 74.4 * |
| Subplot error | 16 | 570,051 | 102,704 | 0.65 | 10.38 | 103 | 0.339 | 18.3 |
| CV | 28.6 | 27.4 | 2.3 | 12.4 | 19.6 | 15.4 | 18.3 | |
| Nitrogen (kg/ha) | Phosphorus (kg/ha) | Seed Yield (kg/ha) | Oil Yield (kg/ha) | 1000 Seed Weight (g) | Harvest Index (%) | Chl. Content (Second Stage) |
|---|---|---|---|---|---|---|
| 0 | 1606.7 c | 722.6 d | 3.4 abc | 19.4 d | 35.5 bc | |
| 0 | 75 | 2427.1 bc | 1077.5 bcd | 4.27 ab | 29.7 ab | 37.2 bc |
| 150 | 1781.8 c | 795.2 cd | 3.53 abc | 19.9 cd | 32.4 c | |
| 0 | 1974.1 c | 860.6 cd | 3.3 bc | 25.5 abcd | 36.4 bc | |
| 100 | 75 | 3284.3 ab | 1466.0 ab | 4.3 a | 25.9 abcd | 34.6 c |
| 150 | 2888.5 bc | 1304.1 bc | 4.1 ab | 27.2 abc | 45.5 ab | |
| 0 | 4234.8 a | 1880.7 a | 4.3 ab | 32.6 a | 50.8 a | |
| 200 | 75 | 3292.8 ab | 1440.9 ab | 3.6 abc | 24.5 bcd | 31.8 c |
| 150 | 2235.9 bc | 971.3 bcd | 3 c | 25.0 bcd | 31.3 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zangani, E.; Afsahi, K.; Shekari, F.; Mac Sweeney, E.; Mastinu, A. Nitrogen and Phosphorus Addition to Soil Improves Seed Yield, Foliar Stomatal Conductance, and the Photosynthetic Response of Rapeseed (Brassica napus L.). Agriculture 2021, 11, 483. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11060483
Zangani E, Afsahi K, Shekari F, Mac Sweeney E, Mastinu A. Nitrogen and Phosphorus Addition to Soil Improves Seed Yield, Foliar Stomatal Conductance, and the Photosynthetic Response of Rapeseed (Brassica napus L.). Agriculture. 2021; 11(6):483. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11060483
Chicago/Turabian StyleZangani, Esmaeil, Kamran Afsahi, Farid Shekari, Eileen Mac Sweeney, and Andrea Mastinu. 2021. "Nitrogen and Phosphorus Addition to Soil Improves Seed Yield, Foliar Stomatal Conductance, and the Photosynthetic Response of Rapeseed (Brassica napus L.)" Agriculture 11, no. 6: 483. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11060483