
Carotenoid Content and Bioaccessibility in Commercial Maize Hybrids
by
 , ,
, ,
Dora Zurak
1, 
Darko Grbeša
1,
Marija Duvnjak
1,
Goran Kiš
1 ,
,
Tatjana Međimurec
2 and
Kristina Kljak
1,* 1
Faculty of Agriculture, University of Zagreb, Svetošimunska 25, 10000 Zagreb, Croatia
2
Ministry of Agriculture, Directorate for Professional Support to the Development of Agriculture and Fisheries, Bani 110, Buzin, 10010 Zagreb, Croatia
*
Author to whom correspondence should be addressed.
Agriculture 2021, 11(7), 586; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070586
Submission received: 24 May 2021
/
Revised: 17 June 2021
/
Accepted: 22 June 2021
/
Published: 24 June 2021
Abstract
:The objective of the present study was to determine the content and bioaccessibility of carotenoids in 104 maize hybrids available at the market. Lutein, zeaxanthin, α- and β-cryptoxanthin, and β-carotene were determined in whole grains and micelles produced during standardized INFOGEST digestibility analysis, and their bioaccessibility was calculated as the ratio of micellar and grain carotenoids. Tested hybrids varied in total carotenoid content, with 34% having total carotenoid content in the range of 15–20 µg/g dry matter (DM) and 41% in the range of 20–25 µg/g DM. The amount of bioaccessible carotenoids increased linearly (p < 0.05) with increasing content in the grain, and decreased among determined carotenoidsd in the order: lutein (52%) > zeaxanthin (43%) > β-carotene (43%) > α-cryptoxanthin (27%) > β-cryptoxanthin (26%). Bioaccessibility of lutein, zeaxanthin, and β-carotene decreased with increasing content in the grain (p < 0.05). On average, only 43% of the total carotenoids were bioaccessible in commercial maize hybrids tested, which should be considered when formulating an animal diet.
1. Introduction
Carotenoids in foods are recognized as compounds with a variety of important biological roles in all living systems. Since humans and animals cannot synthesize them de novo, they are considered essential components of their diet. Due to their pigmenting and antioxidant properties and their function as provitamin A precursors, carotenoids have wide application as food additives and colorants. Despite their wide occurrence in nature, most industrially used carotenoids are synthesized chemically, and only a small portion is obtained by extraction from plants or algae [1]. Nowadays, great attention is attributed to food origin, and synthetic compounds tend to be replaced with natural alternatives. In recent years, efforts have been made to improve the carotenoid content in staple foods to overcome vitamin A deficiency in areas with limited access to animal products, fruits and vegetables [2]. Cereal grains provide more than 50% of the daily requirement of calories, but they often lack in essential minerals and vitamins [3]. However, yellow maize is considered the only one with an appreciable carotenoid content (11.14 µg/g dry matter (DM); [4]).
Maize (Zea mays L.) is a major staple food in Latin America, Asia, and Africa, whereby in other regions of the world, it is mainly used as animal feed [5,6]. Due to their recognized value, maize hybrids are biofortified in some countries—such as Brasil, Nigeria, or India—to achieve higher carotenoid content [7]. Maize grain contains provitamin A carotenoids, mainly β-carotene and β-cryptoxanthin, although the non-provitamin A carotenoids lutein and zeaxanthin dominate. However, maize genotypes differ substantially in carotenoid profile; 64 different genotypes, including varieties of different geographical origins, commercial hybrids, lines in the selection, cornflakes, popcorn, and sweet corn, had total carotenoid content ranging from 0.50 to 68.80 µg/g DM, with ranges 0–29.70 µg/g DM for lutein, 0.30–38.20 µg/g DM for zeaxanthin, 0–5.90 µg/g DM β-cryptoxanthin, 0–3.90 µg/g DM for α-carotene and 0–2.80 µg/g DM for β-carotene [8]. These reported variable contents, as well as low concentrations of provitamin A carotenoids in grains of different genotypes are one of the main reasons why yellow maize is often unjustifiably neglected as a source of carotenoids.
The carotenoid profile of maize genotypes represents only a potential to provide humans and animals with the carotenoids contained in the grain. To exert their biological activities, carotenoids must be released from the grain matrix, solubilized in lipid emulsion droplets, and transferred into mixed micelles, and thus become bioaccessible [9]. Bioaccessibility can be defined as the food fraction released from the matrix into the gastrointestinal tract and available for intestinal absorption/assimilation [10]. The amount of the released carotenoids depends on their physicochemical properties (such as cis vs. trans isomers), deposition form within the food matrix (localization, particle size), degree of food processing (raw vs. processed foods), interaction with other food compounds (lipids, fibers), gut health, nutritional status and genotype [11,12].
Growing concerns about food safety and the negative aspects of pigment production have increased the search for natural alternatives and thus interest in the genetic manipulation of carotenoid content in cereal grains [1,13,14]. However, numerous commercial maize hybrids are available on the market with little information on their carotenoid profiles. Many commercial hybrids contain high concentrations of carotenoids, but the lack of data on their content, release from the grain matrix, and incorporation into mixed micelles does not provide good insight into their provitamin A and pigmentation potential. The current published literature contains data on the bioaccessibility and bioavailability of carotenoids primarily from maize products and includes some, but not all maize grain carotenoids [15,16,17]. Although domestic animals eat minimally processed (post-harvest dried and milled or pelleted) maize grain, information on carotenoid bioaccessibility would provide more appropriate insight into pigmentation potential than carotenoid content. Therefore, the objective of this study was to determine the bioaccessibility of individual and total carotenoids from commercial maize hybrids using an in vitro method and to investigate the relationship between carotenoid content and their bioaccessibility.
2. Materials and Methods
2.1. Sample Preparation
This study was conducted on 104 commercial maize hybrids (Table 1). The maize hybrids were grown in a test field in central Croatia in 2019, with each hybrid planted on a plot 6 m wide and 50 m long. A representative sample of each hybrid was taken at harvest with a total weight of 2 kg; five subsamples were taken immediately after harvest with the maize harvester and combined into one sample. Subsequently, the samples were dried at 40 °C to a moisture content below 12%. To prevent their spoilage and loss of carotenoids, samples were packed in vacuum-sealed bags and stored at −4 °C. Immediately before analysis, the samples were brought to room temperature. Then, a portion of each sample was ground in a laboratory mill with a 1 mm sieve (Cyclotec 1093, Foss Tecator, Sweden) for in vitro digestibility, and the other portion was ground in a ball grinder (MM200, Retsch, Germany) for carotenoid analysis. The moisture content of all samples was determined by drying at 103 ± 2 °C for 4 h.
2.2. In Vitro Digestion Procedure
The INFOGEST in vitro procedure was used to determine the bioaccessibility of maize carotenoids [18]. Although this procedure was developed to mimic digestion in the human tract, it is the most commonly used method to determine carotenoid bioaccessibility [19,20,21,22] and was selected in the present study due to the lack of a standardized digestibility procedure for animals. The potential applicability of the method to animal studies has been tested in some aspects, e.g., Egger et al. [23] compared the INFOGEST procedure with digestion in pigs and showed that protein hydrolysis determined using the method is similar to in vivo protein digestion at the gastric and intestinal endpoints.
The method was adapted to a starch-rich matrix by adding amyloglucosidase and invertase in the small intestine phase, according to Englyst et al. [24]. Additional adaptation was made to stimulate digestive behavior in the digestive tract of poultry; maize samples were ground to pass a 1 mm screen to stimulate gridding action in the gizzard [25]. All enzymes used in this study were purchased from Sigma-Aldrich (St. Louis, MO, USA, SAD). The used enzymes were from porcine origin: α-amylase (A3716, labeled activity 10 U/mg; experimentally determined 10 U/mg), pepsin (P7000, labeled activity 599 U/mg, experimentally determined: 574 U/mg), pancreatin (P7545, labeled activity 8 × USP, experimentally determined trypsin activity 9 U/mg), invertase (I4504, labeled activity ≥ 300 U/mg), amyloglucosidase (A7095, labeled activity ≥ 2 60 U/mg) and bile salts (B8631). The amount of bile salts added to the reaction mixture of enzymes and sample was calculated on the basis that porcine bile extract contains 50% bile salts with an average molecular mass of 442 g/mol [26].
Oral (SSF), gastric (SGF) and intestinal (SIF) fluids used in the in vitro digestion procedure were prepared as described by Brodkorb et al. [18]. Briefly, 1.25 g of the maize sample was mixed with 1.25 mL of ultrapure water, and 2 mL of SSF (pH 7), 0.25 mL of α-amylase solution (1500 U/mL in ultrapure water), 12.5 μL of 0.3 M CaCl2, and ultrapure water to reach 5 mL were added to stimulate oral phase. After incubation for 2 min at 37 °C with horizontal shaking, 4 mL of SGF (pH 3), 3 μL of 0.3 M CaCl2 and 0.5 mL of pepsin solution (40,000 U/mL in ultrapure water) were added to the mixture to simulate the gastric phase. The pH was adjusted to 3 with 6 M HCl, and the volume of 10 mL was adjusted with ultrapure water. The mixture was incubated for 2 h at 37 °C with horizontal shaking. To stimulate the intestinal phase, 4.25 mL of SIF (pH 7), 20 μL of 0.3 M CaCl2, and 2.5 mL of an enzyme mixture containing pancreatin (800 U/mL), amyloglucosidase (13 U/mL) and invertase (0.6 U/mL) were added to the mixture. The pH was adjusted to 7 with 1 M NaOH, and the volume of 20 mL was adjusted with ultrapure water. The mixture was incubated for 3 h at 37 °C with horizontal shaking. At the end of intestinal incubation, the tubes were placed on ice to stop intestinal digestion.
Bioaccessibility was defined as the proportion of carotenoids (individual and total) recovered in the micellar fraction after in vitro digestion relative to the amount of carotenoids in maize samples. The micellar fraction was defined as the fraction obtained after centrifugation to remove microcrystalline carotenoid aggregates and microbial contamination [27].
2.3. Extraction of Carotenoids from Whole Maize Grain
Carotenoids from whole maize grain were extracted according to the procedure described by Kurilich and Juvik [28], using β-apo-carotenal as internal standard. Briefly, after homogenization with ethanol, the samples were saponified with 80% KOH and incubated for 10 min at 85 °C in a water bath. The test tubes were then cooled in an ice bath with the addition of deionized water. The carotenoids were extracted with hexane, which was pipetted into a separate tube after centrifugation at 2200× g for 10 min (Centric 322A, Tehtnica, Slovenia). The extraction procedure was repeated until the colorless upper hexane layer. The collected supernatants were evaporated using rotary vacuum concentrator (RVC 2-25CD plus, Martin Christ, Germany) and dissolved in 200 µL acetonitrile:dichloromethane:methanol (45:20:35, v/v/v) containing 0.1% BHT.
2.4. Extraction of Carotenoids from Micellar Fraction
After digestion, 8 mL of digesta was centrifuged for 1 h at 3200× g at 4 °C, and the clear supernatant was filtered through 0.22 μm nylon membrane syringe filters; 5 mL of the filtered aqueous phase was used for extraction of the bioaccessible carotenoids. The extraction started with the addition of 5 mL of ethanol, 3 mL of hexane and 100 μL of β-apo-carotenal solution as internal standard. After mixing and centrifugation (2500× g, 5 min, 4 °C), the upper hexane phase was collected, and the extraction procedure was repeated until colorless upper hexane layer. The combined extracts were dried using the rotary vacuum concentrator (RVC 2-25 CD), and dissolved in 200 μL of acetonitrile:methanol:methylene chloride (45:20:35, v/v/v) containing 0.1% BHT.2.5. Carotenoid analysis using HPLC.
Lutein, zeaxanthin, α- and β-cryptoxanthin and β-carotene in the extracts of whole maize grain and micellar fraction were quantified according to the reversed-phase HPLC method described by Kurilich and Juvik [28]. Carotenoids were separated and quantified using a SpectraSystem HPLC instrument (Thermo Separation Products, Inc., Waltham, MA, USA) equipped with a quaternary gradient pump, an autosampler and a UV–vis detector. Compounds were separated on two sequentially connected C18 reversed-phase columns Vydac 201TP54 column (5 μm, 4.6 × 150 mm; Hichrom, Reading, UK), followed by a Zorbax RX-C18 column (5 μm, 4.6 × 150 mm; Agilent Technologies, Santa Clara, CA, USA). The separation columns were protected by a Supelguard Discovery C18 guard column (5 μm, 4 × 20 mm; Supelco, Bellefonte, PA, USA). The mobile phase consisted of acetonitrile:methanol:dichloromethane (75:25:5, v/v/v) containing 0.1% BHT and 0.05% triethylamine. An aliquot of 30 μL was injected, and the flow rate was 1.8 mL/min. The separations were performed at room temperature, and carotenoids were monitored at 450 nm.
Carotenoids [lutein (purity 99%), zeaxanthin (purity 99%), α- and β- cryptoxanthin (purity of both 99%), and β-carotene (purity 98%)] were identified by comparing their retention times and quantified by external standardization with calibration curves using commercially available standards (Extrasynthese, France; r2 ≥ 0.99 for all carotenoids). The total carotenoid content was calculated by summing the contents of the individual carotenoids. For total carotenoid content in maize, each hybrid was analyzed in triplicate, and the mean value was taken as the result. Digestion was carried out for each hybrid in triplicate on two separate days, and the mean values of micellar carotenoids, both individual and total, were taken as a result.
2.5. Statistical Analysis
Statistical analyses of the obtained results were performed using SAS statistical software (version 9.4; SAS Institute Inc., Cary, NC, USA). Based on the total carotenoid content, hybrids were classified into five following groups: G1 (<15 µg/g DM), G2 (15–20 µg/g DM), G3 (20–25 µg/g DM), G4 (25–30 µg/g DM), and G5 (>30 µg/g DM). Differences between hybrid groups were subjected to analysis of variance using the MIXED procedure. Means were defined by the least squares means statement and compared using the PDIFF option; letter groups were determined using the PDMIX macro procedure. Contents of the whole grain and micellar carotenoids were assessed using Pearson correlation as implemented in the CORR procedure. Further relationships between carotenoid fractions were assessed using the REG procedure. The threshold for statistical significance was defined as p < 0.05.
3. Results and Discussion
3.1. Carotenoid Content in Tested Commercial Maize Hybrids
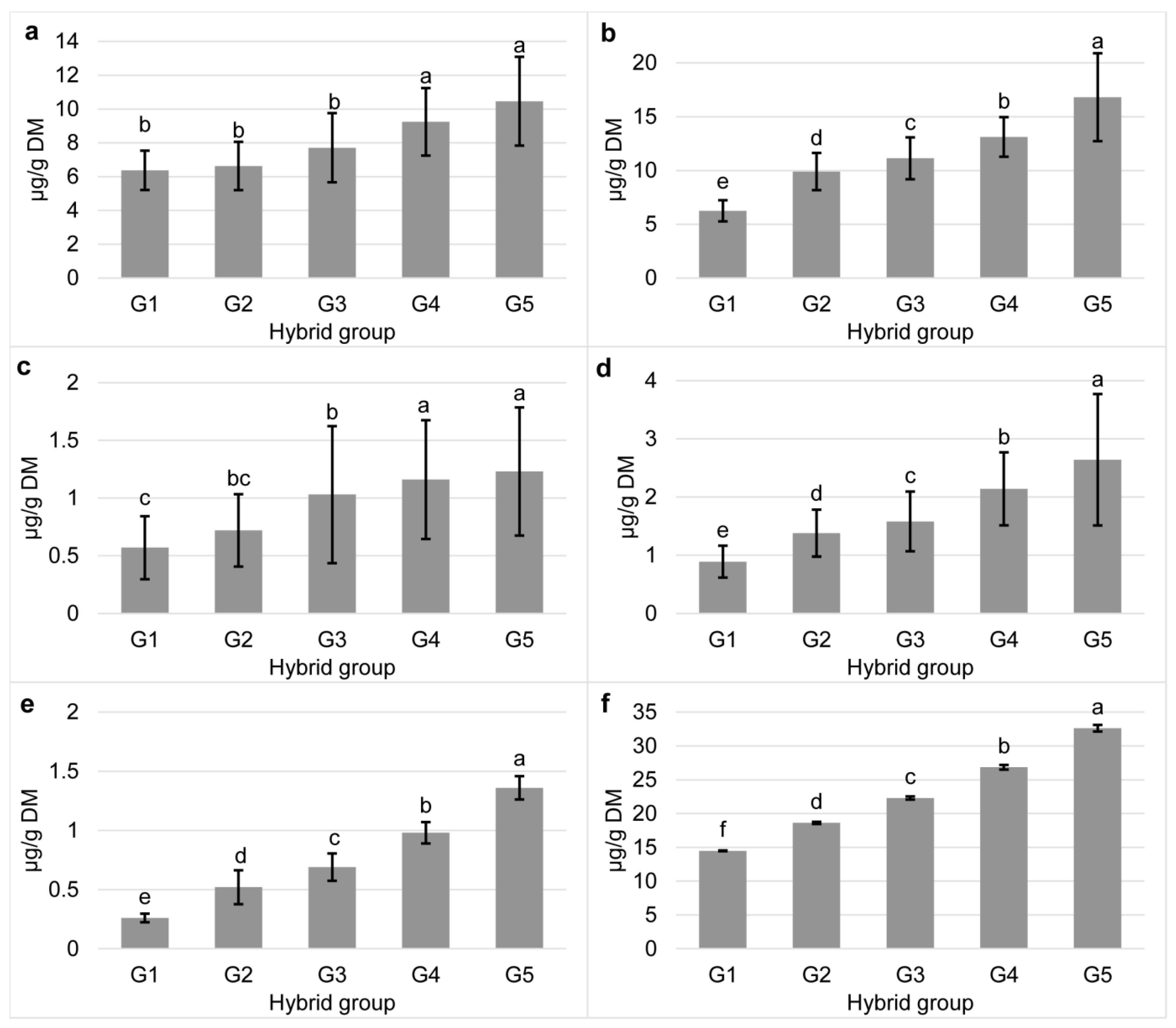
The commercial hybrids analyzed in the present study showed a wide range of total carotenoid content (14.48 to 32.61 µg/g DM), which provided the basis for the classification into five groups. The majority of the hybrids had total carotenoid content in the range of 15 to 25 µg/g DM (34%) and 20 to 25 µg/g DM (41%), while 15% had content in the range of 25 to 30 µg/g DM (Supplementary Table S1). A content of less than 15 µg/g DM was determined in 5% of the hybrids, and the same proportion of hybrids had contents above 30 µg/g DM. Carotenoid content in maize depends on factors such as genotype, climatic conditions and agronomic factors including nitrogen fertilization [29,30]. In total, 88–97% of the variation is due to differences between genotypes in their carotenoid content [28]. To minimize effects other than genotype, the maize hybrids investigated in the present study were grown in the same test field. Compared to the hybrids used in the present study, 24 inbred lines in the study by Tiwari et al. [31] had a similar range of total carotenoids (12.20–30.10 µg/g DM), with content in more than half of the lines analyzed (54%) in the range of G3 group from the present study.
Determined high diversity in total carotenoid content indicated high genotypic variability of tested maize hybrids for all determined carotenoids. With ranges of 6.25–16.81 µg/g DM for zeaxanthin and 6.37–10.46 µg/g DM for lutein, results were consistent with previous reports on the predominance of these carotenoids over other present in maize grain (83–84% depending on the group [32]). Commercial hybrids in the present study had a small prevalence of zeaxanthin over lutein, similarly to the hybrids in studies of Egesel et al. [29] and Halilu et al. [6] and opposite to the inbred lines in studies of Weber [33] and Thakkar and Failla [16] who reported lutein as the most abundant individual carotenoid. However, Saenz et al. [34] showed that the predominance of lutein or zeaxanthin was related to grain vitreousness; in 18 temperate commercial genotypes differing in hardness, lutein negatively while zeaxanthin positively correlated with grain vitreousness. The majority of the hybrids in the present study were dent, but their vitreousness could vary [35], implying that the majority of tested hybrids had higher vitreous over floury endosperm content in the grain. The contents of the other carotenoids were comparable with the reports for different maize genotypes by Kurillich and Juvik [27], Egesel et al. [24], Kean et al. [15], Kandianis [36], and Song et al. [37]. Among the provitamin A carotenoids, obtained amounts of β-cryptoxanthin (0.89–2.64 µg/g DM) exceeded the content of β-carotene (0.26–1.36 µg/g DM), which was consistent with other studies [28,29,38,39].
In general, the content of all carotenoids increased linearly with increasing carotenoid group (p < 0.001; Figure 1), indicating that the content of all individual carotenoids in commercial maize hybrids increases linearly with total carotenoid content. In addition, moderate to high correlations were found between carotenoids of the same biosynthetic pathway (r = 0.63 for lutein—α-cryptoxanthin, r = 0.71 for zeaxanthin—β-cryptoxanthin, r = 0.62 for zeaxanthin—β-carotene, r = 0.61 for β-cryptoxanthin—β-carotene, p < 0.001 for all), while traits in different branches showed low or no correlation. The results were in agreement with previous studies in which correlation analyses showed simultaneous increases in the contents of carotenoids produced in the same pathway branch, and this observation could be used for genetic improvement of provitamin A carotenoids in maize [6,40]. Moreover, a recent study by Wang et al. [41] showed that inbred lines containing a nonfunctional Ven1 allele exhibited a decrease in polar and an increase in nonpolar carotenoids in the amyloplasts, which provides insight into breeding vitreous grain varieties and high vitamin A content in maize.
3.2. Bioaccessibility of Carotenoids in Commercial Maize Hybrids
Micellarization efficiency is a measure of bioaccessibility and takes into account factors such as carotenoid species, food matrix, chemical state, ingested amount, and absorption modifiers [12,42]. Once carotenoids are released from the food matrix, they dissolve in the oily phase of lipid droplets, which is an important step preceding their incorporation into mixed micelles [43]. Three main factors affecting the transfer of carotenoids from emulsion lipid droplets into micelles are the type of carotenoid, pH, and concentration in bile lipids [44]. Since the above parameters were constant in this study, the micellarization efficiency of carotenoids was related to their polarity and the ratio of their concentrations in feed matrix [16,27].
In general, the amount of bioaccessible carotenoids increased with the amount of carotenoids in the maize hybrid; the correlation coefficients were 0.79 for lutein, 0.70 for zeaxanthin, 0.85 for α-cryptoxanthin, 0.78 for β-cryptoxanthin, 0.51 for β-carotene, and 0.62 for total carotenoids (p < 0.001, respectively). Consistent with this positive correlation, the amount of bioaccessible carotenoids increased linearly with the increasing carotenoid content in hybrid groups (p < 0.05). These results imply that increased carotenoid content in maize hybrid translates to increased bioaccessibility, leading to increased bioavailability when fed to animals. When the content of bioaccessible carotenoids was plotted against the content in maize grain (Figure 2), the linear regression yielded coefficients of determination between 0.26 and 0.73. Among the carotenoids detected in the tested maize hybrids, the best-fitting models were found for lutein, and α- and β-cryptoxanthin and these models were able to explain 68%, 73%, and 64% of the variability, respectively, in predicting the content of bioaccessible carotenoids from the content in the grain.
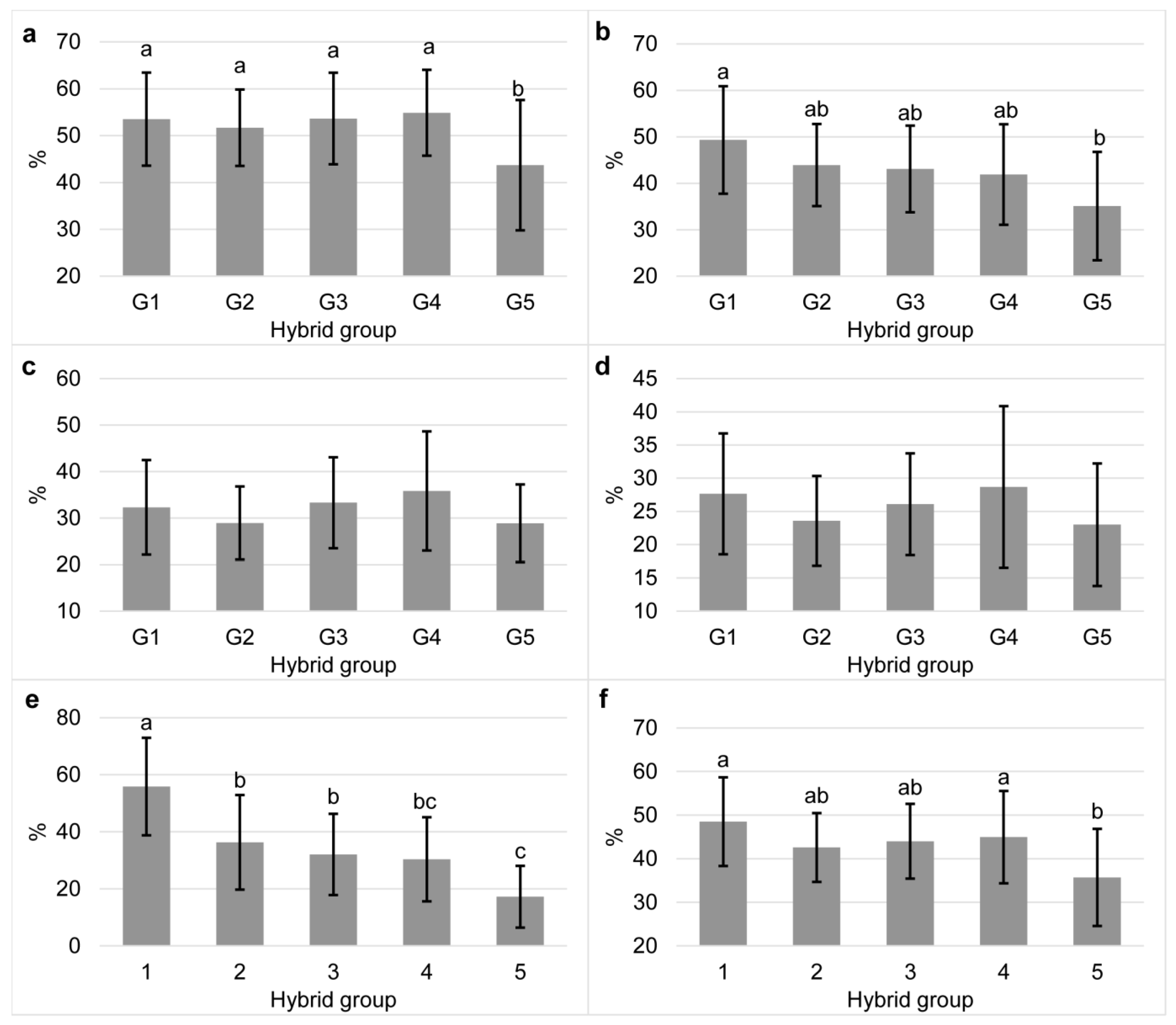
When the amount of bioaccessible carotenoids was expressed relative to the content in the grain, i.e., as bioaccessibility, the difference between groups was less pronounced, and the range of values within groups was more similar (Figure 3). Furthermore, the difference between the groups was not found for bioaccessibility of α- and β-cryptoxanthin. A proportionality was observed only for zeaxanthin, β-carotene and total carotenoids (p < 0.05), and this effect was opposite to the content in the maize grain or the content of bioaccessible carotenoids. This finding suggests that the bioaccessibility of those carotenoids decreased with increasing content in grain, and it is possible that competition for incorporation into micelles occurred at higher concentrations [45]. This decreasing proportionality was most pronounced for β-carotene, where the group of hybrids with <15 μg/g DM of total carotenoids had bioaccessibility of 55.9% and the group with >30 μg/g DM had 17.2%.
The average bioaccessibility of lutein, zeaxanthin, β-cryptoxanthin, and β-carotene determined in the present study exceeded the average efficiency of incorporation into synthetic micelles reported in the study by Thakkar and Faillla [16] for raw maize samples (range of 16–30%). In contrast, Dube et al. [46] reported higher average bioaccessibility of these carotenoids (62 ± 5.3%, 65 ± 4.7%, 54 ± 9.5%, and 49 ± 7.5%, respectively) for 10 maize genotypes, but the authors did not use a standardized in vitro digestibility procedure. Nonetheless, the determined broad micellarization efficiency could also be related to the maize grain matrix characteristics, particularly lipid content. Since lipids are required to incorporate carotenoids into micelles [45], the differences in lipid content among the tested maize hybrids could influence the bioaccessibility of carotenoids in the present study.
Among the carotenoids in the tested maize hybrids, the highest values of bioaccessibility were found for lutein (average 51.5%), followed by zeaxanthin (average 42.7%), β-carotene (average 34.4%), α-cryptoxanthin (average 26.5%), and β-cryptoxanthin (average 25.8%). The predominance of lutein and zeaxanthin incorporating in micelles over β-carotene agrees with previous results reported by Thakkar and Failla [16] and Dube et al. [46]. The obtained results indicate the preferential incorporation of xanthophylls in mixed micelles due to their polarity, which has been previously reported [15,16,43,47]. Due to their polar nature and location on the surface of emulsions, they can be spontaneously transferred from lipid droplets to micelles, unlike carotenes which are located in the core [43].
The bioaccessibility of β-carotene in commercial maize hybrids was higher than that of α- and β-cryptoxanthin, and this finding was surprising since β-carotene is the least polar carotenoid among those determined in the hybrids tested. Results opposite to higher bioaccessibility of β-carotene over α- and β-cryptoxanthin in the present study have been reported for raw and cooked maize [17], whole grain and degermed maize meal products [15] and maize porridge [16]. A possible reason for the high average bioaccessibility of β-carotene is its interaction with lutein. Thakkar and Failla [16] found that when the molar ratio of lutein to β-carotene is ≥7, the micellarization efficiency of β-carotene increases, which they explain by the retention of β-carotene within the core of oil droplets when lipase-mediated hydrolysis of tri-glycerides is limited, reducing the possibility of transferring hydrocarbon carotenoid into micelles. In the present study, this molar ratio was above 7 in G1, G2, G3, and G4 (23, 12, 11, and 9, respectively). The highest bioaccessibility of β-carotene was in G1 (55.86%), where possibly the enhancing effect of lutein was highest (molar ratio of lutein to β-carotene of 23). In contrast, the lowest bioaccessibility was in G5 (17.22%), where the molar ratio of lutein to β-carotene was 7.
4. Conclusions
The results obtained in the present study show a considerable variation in the carotenoid composition of maize samples in agreement with a wide genotype variability among the commercial hybrids. The amount of bioaccessible carotenoids increased with carotenoid content in the tested hybrids, which translates to increased bioavailability when fed to animals. However, the bioaccessibility decreased with increasing content in the hybrids, suggesting that high carotenoid in grain does not imply efficient micellarization during digestion. Only 43% of the total grain carotenoids is bioaccessible and thus available for absorption by the monogastric animal. This result suggests that although maize grain is the only cereal with significant content of carotenoids, knowledge of their content is not sufficient to predict potential utilization in animals and this should be considered in the formulation of animal diets. As maize could contribute substantially to carotenoid content of animal diets due to its high proportion, further steps should be taken to evaluate factors that could increase bioaccessibility of carotenoids.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/agriculture11070586/s1, Table S1: Classification of tested hybrids into groups based on total carotenoid content in the grain.
Author Contributions
Conceptualization, K.K. and D.G.; Methodology, K.K. and M.D.; Validation, K.K., D.G. and G.K.; Formal analysis, D.Z.; Investigation, D.Z., K.K, D.G. and G.K.; Resources, K.K., M.D. and T.M.; Data curation, K.K.; Writing—original draft preparation, D.Z.; Writing—review and editing, K.K., D.G., M.D., G.K. and T.M.; Visualization, D.Z.; Supervision, K.K.; Project administration, K.K.; Funding acquisition, K.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been fully supported by Croatian Science Foundation under the project ColourMaize (“Bioavailability of maize carotenoids in laying hens: effect of grain microstructure and diet composition”; IP-2019-04-9063). The work of doctoral student Dora Zurak has been fully supported by the “Young researchers’ career development project—training of doctoral students” of the Croatian Science Foundation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to the use of commercial maize hybrids in the study and intellectual property rights of the hybrid producers.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Mezzomo, N.; Ferreira, S.R.S. Carotenoids functionality, sources, and processing by supercritical technology: A review. J. Chem. 2016, 3164312. [Google Scholar] [CrossRef] [Green Version]
- Menkir, A.; Pixley, K.; Maziya-Dixon, B.; Gedil, M. Recent advances in breeding maize for enhanced provitamin A content. In Meeting the Challenges of Global Climate Change and Food Security through Innovative Maize Research, Proceedings of the Third National Maize Workshop of Ethiopia, Addis Ababa, Ethiopia, 18–20 April 2011; Worku, M., Twumasi Afriyie, S., Legesse, W., Berhanu Tadesse, E., Demisie, G., Bogale, G., Dagne Wegary, G., Prasanna, B.M., Eds.; International Maize and Wheat Improvement Center: Texcoco, Mexico, 2012; p. 290. [Google Scholar]
- Ashokkumar, K.; Govindaraj, M.; Karthikeyan, A.; Shobhana, V.G.; Warkentin, T.D. Genomics-integrated breeding for carotenoids and folates in staple cereal grains to reduce malnutrition. Front. Genet. 2020, 11, 414. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, A.J.; Mandić, A.I.; Bantis, F.; Böhm, V.; Borge, G.I.A.; Brnčić, M.; Bysted, A.; Cano, M.P.; Graça Dias, M.; Elgersma, A.; et al. A comprehensive review on carotenoids in foods and feeds: Status quo, applications, patents, and research needs. Crit. Rev. Food Sci. Nutr. 2021. [Google Scholar] [CrossRef] [PubMed]
- Muzhingi, T.; Yeum, K.J.; Russell, R.M.; Johnson, E.; Qin, J.; Tang, G. Determination of carotenoids in yellow maize, the effects of saponification and food preparations. Int. J. Vitam. Nutr. Res. 2008, 78, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Halilu, A.D.; Ado, S.G.; Aba, A.D.; Usman, S.I. Genetics of carotenoids for provitamin A biofortification in tropical-adapted maize. Crop. J. 2016, 4, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.S.; Saltzman, A.; Virk, P.S.; Pfeiffer, W.H. Progress update: Crop development of biofortified staple food crops under HarvestPlus. Afr. J. Food Nutr. Dev. 2017, 17, 11905–11935. [Google Scholar] [CrossRef]
- Berardo, N.; Brenna, O.V.; Amato, A.; Valoti, P.; Pisacane, V.; Motto, M. Carotenoids concentration among maize genotypes measured by near infrared reflectance spectroscopy (NIRS). Innov. Food Sci. Emerg. Technol. 2004, 5, 393–398. [Google Scholar] [CrossRef]
- Schieber, A.; Weber, F. Carotenoids. In Handbook on Natural Pigments in Food and Beverages, Industrial Applications for Improving Food Color, 1st ed.; Carle, R., Schweiggert, R.M., Eds.; Woodhead Publishing Series in Food Science, Technology and Nutrition: Cambridge, UK, 2016; pp. 101–123. [Google Scholar] [CrossRef]
- Carbonell-Capella, J.M.; Buniowska, M.; Barba, F.J.; Esteve, M.J.; Frìgola, A. Analytical methods for determining bioavailability and bioaccessibility of bioactive compounds from fruits and vegetables: A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 155–171. [Google Scholar] [CrossRef]
- van Het Hof, K.H.; West, C.E.; Weststrate, J.A.; Hautvast, J.G. Dietary factors that affect the bioavailability of carotenoids. J. Nutr. 2000, 130, 503–506. [Google Scholar] [CrossRef]
- Dima, C.; Assadpour, E.; Dima, S.; Mahdi Jafari, S. Bioavailability of nutraceuticals: Role of the food matrix, processing conditions, the gastrointestinal tract, and nanodelivery systems. Compr. Rev. Food Sci. Food Saf. 2020, 19, 954–994. [Google Scholar] [CrossRef]
- Sugiura, M.; Nakamura, M.; Ikoma, Y.; Yano, M.; Ogawa, K.; Matsumoto, H.; Kato, M.; Ohshima, M.; Nagao, A. Serum carotenoid concentrations are inversely associated with serum aminotransferases in hyperglycemic subjects. Diabetes Res. Clin. Pract. 2006, 71, 82–91. [Google Scholar] [CrossRef]
- Zhai, S.; Xia, X.; He, Z. Carotenoids in staple cereals: Metabolism, regulation, and genetic manipulation. Front. Plant Sci. 2016, 7, 1197. [Google Scholar] [CrossRef] [Green Version]
- Kean, E.G.; Hamaker, B.R.; Ferruzzi, M.G. Carotenoid bioaccessibility from whole grain and degermed maize meal products. J. Agric. Food Chem. 2008, 56, 9918–9926. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, S.K.; Failla, M.L. Bioaccessibility of pro-vitamin A carotenoids is minimally affected by non pro-vitamin A xanthophylls in maize (Zea mays sp.). J. Agric. Food Chem. 2008, 56, 11441–11446. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Jayadeep, P.A. Comparison of total carotenoids, lutein, zeaxanthin, and β-carotene content in maize employing solvent extraction and in vitro physiological methods. J. Food Biochem. 2018, 42, 1–9. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Balance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Estévez-Santiago, R.; Olmedilla-Alonso, B.; Fernández-Jalao, I. Bioaccessibility of provitamin A carotenoids from fruits: Application of a standardised static in vitro digestion method. Food Funct. 2016, 7, 1354–1366. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.B.; Mariutti, L.R.B.; Mercadante, A.Z. An in vitro digestion method adapted for carotenoids and carotenoid esters: Moving forward towards standardization. Food Funct. 2016, 7, 4992–5001. [Google Scholar] [CrossRef] [PubMed]
- Petry, F.C.; Mercadante, A.Z. Impact of in vitro digestion phases on the stability and bioaccessibility of carotenoids and their esters in mandarin pulps. Food Funct. 2017, 8, 3951–3963. [Google Scholar] [CrossRef]
- Tan, Y.; Zhang, Z.; Zhou, H.; Xiao, H.; McClements, D.J. Factors impacting lipid digestion and β-carotene bioaccessibility assessed by standardized gastrointestinal model (INFOGEST): Oil droplet concentration. Food Funct. 2020, 11, 7126–7137. [Google Scholar] [CrossRef]
- Egger, L.; Schlegel, P.; Baumann, C.; Stoffers, H.; Guggisberg, D.; Brügger, C.; Dürr, D.; Stoll, P.; Vergères, G.; Portmann, R. Physiological comparability of the harmonized INFOGEST in vitro digestion method to in vivo pig digestion. Food Res. Int. 2017, 102, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 46, S33–S50. [Google Scholar] [PubMed]
- Weurding, R.E.; Veldman, A.; Veen, W.A.; van der Aar, P.J.; Verstegen, M.W. In vitro starch digestion correlates well with 414 rate and extent of starch digestion in broiler chickens. J. Nutr. 2001, 131, 2336–2342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinarov, Z.; Petkova, Y.; Tcholakova, S.; Denkov, N.; Stoyanov, S.; Pelan, E.; Lips, A. Effects of emulsifier charge and concentration on pancreatic lipolysis. 1. In the absence of bile salts. Langmuir 2012, 28, 8127–8139. [Google Scholar] [CrossRef]
- Mashurabad, P.C.; Palika, R.; Jyrwa, Y.W.; Bhaskarachary, K.; Pullakhandam, R. Dietary fat composition, food matrix and 420 relative polarity modulate the micellarization and intestinal uptake of carotenoids from vegetables and fruits. J. Food Sci. Technol. 2017, 54, 333–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurilich, A.C.; Juvik, J.A. Quantification of carotenoid and tocopherol antioxidants in Zea mays. J. Agric. Food Chem. 1999, 47, 1948–1955. [Google Scholar] [CrossRef]
- Egesel, C.O.; Wong, J.C.; Lambert, R.J.; Rocheford, T.R. Combining ability of maize inbreds for carotenoids and tocopherols. Crop. Sci. 2003, 43, 818–823. [Google Scholar] [CrossRef]
- Ortiz-Covarrubias, Y.; Dhliwayo, T.; Palacios-Rojas, N.; Ndhlela, T.; Magorokosho, C.; Aguilar-Rincón, V.H.; Cruz-Morales, A.S.; Trachsel, S. Effects of drought and low nitrogen stress on provitamin A carotenoid content of biofortified maize hybrids. Crop. Sci. 2019, 59, 2521–2532. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, A.; Prasanna, B.M.; Hossain, F.; Guruprasad, K.N. Analysis of genetic variability for kernel carotenoid concentration in selected maize inbred lines. Indian J. Genet. Plant Breed. 2012, 72, 1–6. [Google Scholar]
- Kumar, S.; Sangwan, S.; Yadav, R.; Langyan, S.; Singh, M. Maize carotenoid composition and biofortification for provitamin A activity. In Maize: Nutrition Dynamics and Novel Uses; Chaudhary, D.P., Kumar, S., Langyan, S., Eds.; Springer: New Delhi, India, 2014; pp. 83–91. [Google Scholar]
- Weber, E.J. Carotenoids and tocols of corn grain determined by HPLC. J. Am. Oil Chem. Soc. 1987, 64, 1129–1134. [Google Scholar] [CrossRef]
- Saenz, E.; Borrás, L.; Gerde, J.A. Carotenoid profiles in maize genotypes with contrasting kernel hardness. J. Cereal Sci. 2021, 99, 103206. [Google Scholar] [CrossRef]
- Kljak, K.; Grbeša, D. Carotenoid content and antioxidant activity of hexane extracts from selected Croatian corn hybrids. Food Chem. 2015, 167, 402–408. [Google Scholar] [CrossRef]
- Kandianis, C. Genetic dissection of carotenoid concentration and compositional traits in maize grain. Doctoral Dissertation, University of Illinois, Urbana, IL, USA, 2010. Available online: https://www.ideals.illinois.edu/handle/2142/14699 (accessed on 9 May 2021).
- Song, J.; Li, D.; He, M.; Chen, J.; Liu, C. Comparison of carotenoid composition in immature and mature grains of corn (Zea Mays L.) varieties. Int. J. Food Prop. 2016, 19, 351–358. [Google Scholar] [CrossRef]
- Menkir, A.; Gedil, M.; Tanumihardjo, S.; Adepoju, A.; Bossey, B. Carotenoid accumulation and agronomic performance of maize hybrids involving parental combinations from different marker-based groups. Food Chem. 2014, 148, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Menkir, A.; Liu, W.; White, W.S.; Maziya-Dixon, B.; Rocheford, T. Carotenoid diversity in tropical-adapted yellow maize inbred lines. Food Chem. 2008, 109, 521–529. [Google Scholar] [CrossRef]
- Menkir, A.; Maziya-Dixon, B.; Mengesha, W.; Rocheford, T.; Oladeji Alamu, E. Accruing genetic gain in pro-vitamin A enrichment from harnessing diverse maize germplasm. Euphytica 2017, 213, 105. [Google Scholar] [CrossRef]
- Wang, H.; Huang, Y.; Xiao, Q.; Huang, X.; Li, C.; Gao, X.; Wang, Q.; Xiang, X.; Zhu, Y.; Wang, J.; et al. Carotenoids modulate kernel texture in maize by influencing amyloplast envelope integrity. Nat. Commun. 2020, 11, 5346. [Google Scholar] [CrossRef]
- Rodriguez Concepcion, M.; Avalos, J.; Luisa Bonet, M.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Carmen Limon, M.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [Green Version]
- Perera, C.O.; Mei Yen, G. Functional properties of carotenoids in human health. Int. J. Food Prop. 2007, 10, 201–230. [Google Scholar] [CrossRef]
- Tyssandier, V.; Lyan, B.; Borel, P. Main factors governing the transfer of carotenoids from emulsion lipid droplets to micelles. Biochim. Biophys. Acta 2001, 1533, 285–292. [Google Scholar] [CrossRef]
- Priyadarshani, A.M.B. A review on factors influencing bioaccessibility and bioefficacy of carotenoids. Crit. Rev. Food Sci. Nutr. 2017, 57, 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- Dube, N.; Mashurabad, P.C.; Hossain, F.; Pullakhandam, R.; Thingnganing, L.; Kumar Bharatraj, D. β-Carotene bioaccessibility from biofortified maize (Zea mays) is related to its density and is negatively influenced by lutein and zeaxanthin. Food Funct. 2018, 9, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.B.; Chitchumroonchokchai, C.; Mariutti, L.R.B.; Mercadante, A.Z.; Failla, M. Comparison of two static in vitro digestion methods for screening the bioaccessibility of carotenoids in fruits, vegetables, and animal products. J. Agric. Food Chem. 2017, 65, 11220–11228. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Content of individual (a), lutein; (b), zeaxanthin; (c), α-cryptoxanthin; (d), β-cryptoxanthin; (e), β-carotene) and total carotenoids (f) of commercial maize hybrids classified into five groups according to the total carotenoid content (G1, <15; G2, 15–20; G3, 25–25; G4, 25–30, and G5, >30 µg/g DM). Different small letters represent statistically significant differences (p < 0.05) between hybrid groups. Error bars represent SD.
Figure 1.
Content of individual (a), lutein; (b), zeaxanthin; (c), α-cryptoxanthin; (d), β-cryptoxanthin; (e), β-carotene) and total carotenoids (f) of commercial maize hybrids classified into five groups according to the total carotenoid content (G1, <15; G2, 15–20; G3, 25–25; G4, 25–30, and G5, >30 µg/g DM). Different small letters represent statistically significant differences (p < 0.05) between hybrid groups. Error bars represent SD.

Figure 2.
Linear regression of carotenoids, individual (a), lutein; (b), zeaxanthin; (c), α-cryptoxanthin; (d), β-cryptoxanthin; (e), β-carotene and total (f), in whole maize grain and bioaccessible after INFOGEST in vitro digestibility analysis.
Figure 2.
Linear regression of carotenoids, individual (a), lutein; (b), zeaxanthin; (c), α-cryptoxanthin; (d), β-cryptoxanthin; (e), β-carotene and total (f), in whole maize grain and bioaccessible after INFOGEST in vitro digestibility analysis.

Figure 3.
Bioaccessibility of individual (a), lutein; (b), zeaxanthin; (c), α-cryptoxanthin; (d), β-cryptoxanthin; (e), β-carotene and total carotenoids (f) of commercial maize hybrids classified into five groups according to the total carotenoid content (G1, <15; G2, 15–20; G3, 25–25; G4, 25–30, and G5, >30 µg/g DM). Different small letters represent statistically significant differences (p < 0.05) between hybrid groups. Error bars represent SD.
Figure 3.
Bioaccessibility of individual (a), lutein; (b), zeaxanthin; (c), α-cryptoxanthin; (d), β-cryptoxanthin; (e), β-carotene and total carotenoids (f) of commercial maize hybrids classified into five groups according to the total carotenoid content (G1, <15; G2, 15–20; G3, 25–25; G4, 25–30, and G5, >30 µg/g DM). Different small letters represent statistically significant differences (p < 0.05) between hybrid groups. Error bars represent SD.


{kind=link}
{kind=link}
{kind=link}
Table 1.
List of tested commercial maize hybrids.
| Hybrid | Hybrid | Hybrid | Hybrid |
|---|---|---|---|
| Bc Institut Agram | KWS Kapitolis | Pioneer P0164 | PIO Os 522 |
| Bc Institut Alibi | KWS Kollegas | Pioneer P0200 | PIO Os 3850 |
| Bc Institut Bc 323 | KWS Kolumbaris | Pioneer P0216 | PIO Posavac 36 |
| Bc Institut Bc 344 | KWS Konfites | Pioneer P0217 | PIO Velimir |
| Bc Institut Bc 415 | KWS Kashmir | Pioneer P0412 | RWA ES Inventive |
| Bc Institut Bc 418 | KWS Orlando | Pioneer P0725 | RWA Ajowan |
| Bc Institut Bc 424 | KWS KxB 8386 | Pioneer P9241 | RWA Inclusiv |
| Bc Institut Bc 525 | KWS KxB 8453 | Pioneer P9300 | RWA Persic |
| Bc Institut Bc 572 | KWS Smaragd | Pioneer P9363 | RWA Gladiator |
| Bc Institut Instruktor | LG 30.3115 | Pioneer P9415 | RWA Glumanda |
| Bc Institut Kekec | LG 30.315 | Pioneer P9757 | RWA Ulyxxe |
| Bc Institut Majstor | LG 31.322 | Pioneer P9889 | RWA Hexagon |
| Bc Institut Pajdaš | LG 31.377 | Pioneer P9903 | RWA Tweetor |
| Bc Institut Tesla | LG 31.545 | Pioneer P9911 | RWA Urbanix |
| Bc Institut Thriler | LG 368/08 | Pioneer P9978 | Syngenta Sy Andromeda |
| DKC 4670 | LG Shannon | PIO 1 Tomasov | Syngenta Sy Atomic |
| DKC 4920 | MAS 34B | PIO Jablan | Syngenta Sy Bilbao |
| DKC 4943 | MAS 48L | PIO Kulak | Syngenta Sy Carioca |
| DKC 5031 | MAS 64P | PIO Os 3114 | Syngenta Sy Chorintos |
| DKC 5068 | NS seme 3022 | PIO Os 3150 | Syngenta Sy Kreon |
| DKC 5075 | NS seme 4015 | PIO Os 3450 | Syngenta Sy Lucius |
| DKC 5093 | NS seme 4051 | PIO Os 378 | Syngenta Sy Photon |
| DKC 5182 | NS seme 6102 | PIO Os 398 | Syngenta Sy Premeo |
| DKC 5685 | NS seme 6102 | PIO Os 4014 | Syngenta Sy Sandoro |
| DKC 5830 | NS seme Haris | PIO Os 4015 | Syngenta Sy Senko |
| KWS Balasco | P0023 | Os 403 | Syngenta Sy Zoan |
1 PIO—Agricultural Institute Osijek.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zurak, D.; Grbeša, D.; Duvnjak, M.; Kiš, G.; Međimurec, T.; Kljak, K. Carotenoid Content and Bioaccessibility in Commercial Maize Hybrids. Agriculture 2021, 11, 586. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070586
AMA Style
Zurak D, Grbeša D, Duvnjak M, Kiš G, Međimurec T, Kljak K. Carotenoid Content and Bioaccessibility in Commercial Maize Hybrids. Agriculture. 2021; 11(7):586. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070586
Chicago/Turabian StyleZurak, Dora, Darko Grbeša, Marija Duvnjak, Goran Kiš, Tatjana Međimurec, and Kristina Kljak. 2021. "Carotenoid Content and Bioaccessibility in Commercial Maize Hybrids" Agriculture 11, no. 7: 586. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070586
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.