
Introgression of QTLs for Drought Tolerance into Farmers’ Preferred Sorghum Varieties

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Location
2.2. Markers Used for Drought Tolerance
2.3. Introgression of STG QTLs from Donor Parents to the Recurrent Parents and Genotyping
2.4. The Development of F1, BC1F1 and BC2F1 Populations
2.5. Development of BC2F2 and BC2F3
2.6. Genotyping of BC2F1 and BC2F3 Populations
2.7. Molecular Data Analysis
3. Results
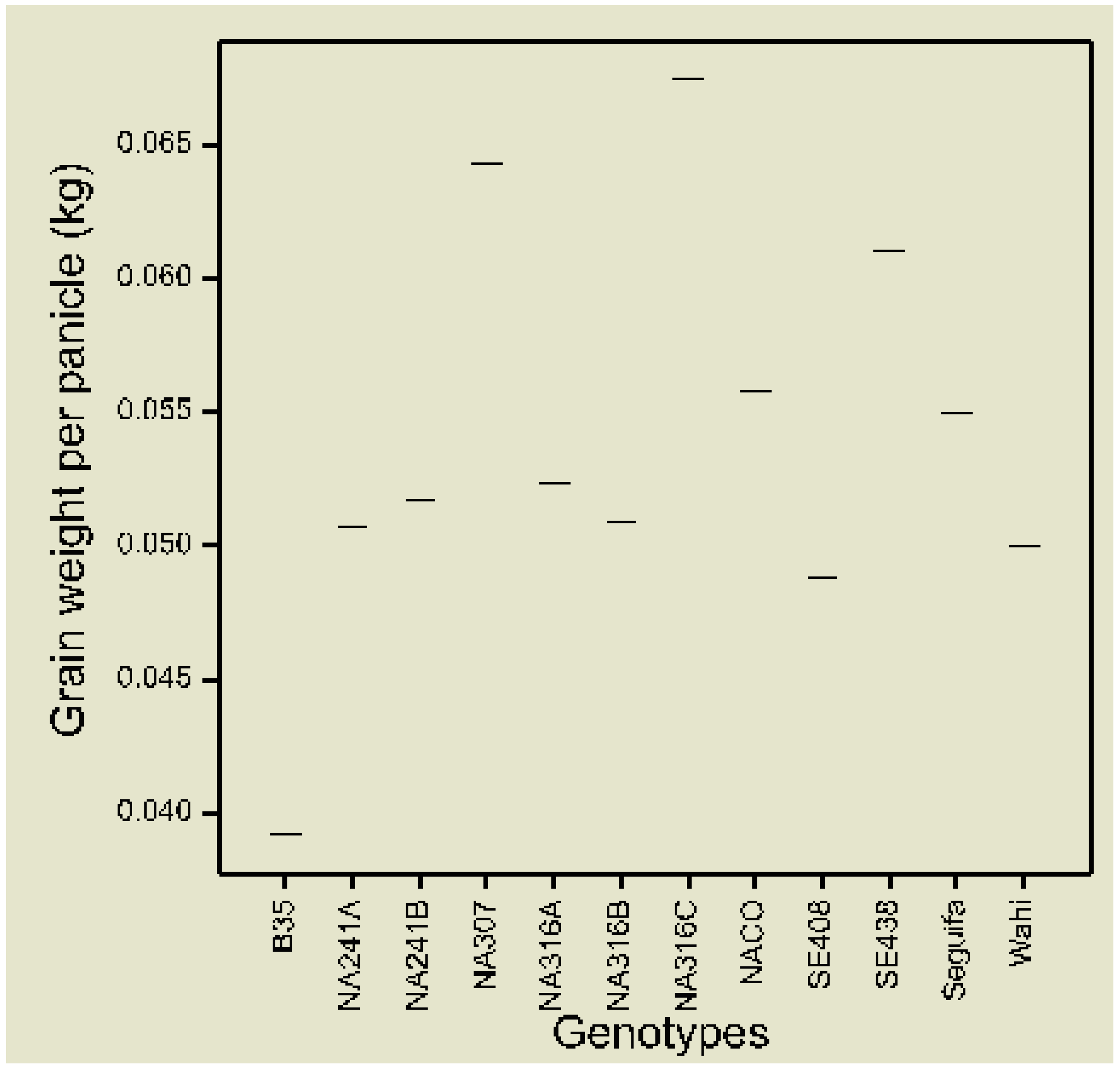
3.1. Phenotypic Data
3.2. BC2F1 Genotyping
3.3. Phenotypic Data of BC2F3 Populations’ Results
3.4. BC2F3 Genotyping Results
4. Discussion
5. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barron, J.; Rockström, J.; Gichuki, F.; Hatibu, N. Dry spell analysis and maize yields for two semi-arid locations in East Africa. Agric. Meteorol. 2003, 117, 23–37. [Google Scholar] [CrossRef]
- Chander, S.; Garcia-Oliveira, A.L.; Gedil, M.; Shah, T.; Otusanya, G.O.; Asiedu, R.; Chigeza, G. Genetic diversity and population structure of soybean lines adapted to sub-Saharan Africa using single nucleotide polymorphism (SNP) markers. Agronomy 2021, 11, 604. [Google Scholar] [CrossRef]
- Premachandra, G.S.; Hahn, D.T.; Joly, R.J. Leaf water relations and gas exchange in two grain sorghum genotypes differing in their pre- and post-flowering drought tolerance. J. Plant Physiol. 1994, 143, 96–101. [Google Scholar] [CrossRef]
- Kamal, S.M.; Gorafi, Y.S.A.; Tsujimoto, H.; Ghanim, A.M.A. Stay-Green QTLs response in adaptation to post flowering drought depends on the drought deverity. BioMed Res. Int. 2018, 2018, 1–15. [Google Scholar] [CrossRef]
- Borrell, A.K.; van Oosterom, E.J.; Mullet, J.E.; George-Jaeggli, B.; Jordan, D.R.; Klein, P.E.; Hammer, G.L. Stay-green alleles individually enhance grain yield in sorghum under drought by modifying canopy development and water uptake patterns. New Phytol. 2014, 203, 817–830. [Google Scholar] [CrossRef]
- Rosenow, D.T.; Clark, L.E. Drought tolerance in sorghum. In Proceedings of the 36th Annual Corn and Sorghum Industry Research Conference; Loden, H.D., Wilkinson, D., Eds.; American Seed trade Association: Washington, DC, USA, 1981; pp. 18–31. [Google Scholar]
- Slegers, M.F.W. Exploring Farmers’ Perceptions of Drought in Tanzania and Ethiopia. Ph.D Thesis, Wageningen University, Wageningen, The Netherlands, 2008. [Google Scholar]
- Velazco, J.G.; Jordan, D.R.; Mace, E.S.; Hunt, C.H.; Malosetti, M.; van Eeuwijk, F.A. Genomic Prediction of Grain Yield and Drought-Adaptation Capacity in Sorghum is Enhanced by Multi-Trait Analysis. Front. Plant Sci. 2003, 10, 997. [Google Scholar] [CrossRef] [Green Version]
- Sory, S. Marker Assisted Selection for Post-Flowering Drought Tolerance in Sorghum bicolor [L.] Moench. Ph.D. Thesis, University of Ghana, Accra, Ghana, December 2015. [Google Scholar]
- Serraj, R.; Hash, T.C.; Buhariwalla, H.K.; Bidinger, F.R.; Folkertsma, R.T.; Chandra, S.; Gaur, P.; Kashiwagi, J.; Nigam, S.N.; Rupakula, A.; et al. Marker-assisted breeding for crop drought tolerance at ICRISAT: Achievements and prospects. In Proceedings of the International Congress; In the Wake of the Double Helix: From the Green Revolution to the Gene Revolution; Tuberosa, R., Phillips, R.L., Gale, M., Eds.; Avenue Media: Bologna, Italy, 2003; pp. 217–238. [Google Scholar]
- Boyles, R.E.; Brenton, Z.W.; Kresovich, S. Genetic and genomic resources of sorghum to connect genotype with phenotype in contrasting environments. Plant J. 2019, 97, 19–39. [Google Scholar] [CrossRef] [Green Version]
- Kamal, N.M.; Gorafi, Y.S.A.; Abdeltwab, H.; Abdalla, I.; Tsujimoto, H.; Ghanim, A.M.A. A New Breeding Strategy towards Introgression and Characterization of Stay-Green QTL for Drought Tolerance in Sorghum. Agriculture 2021, 11, 598. [Google Scholar] [CrossRef]
- Vadez, V.; Deshpande, S.P.; Kholova, J.; Hammer, G.L.; Borrell, A.K.; Talwar, H.S.; Hash, C.T. Stay-green quantitative trait loci’s effects on water extraction, transpiration efficiency and seed yield depend on recipient parent background. Funct. Plant Biol. 2011, 38, 553–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribaut, J.M.; Betran, J. Single large-scale marker assisted selection (SLS-MAS). Mol. Breed. 1999, 5, 531–541. [Google Scholar] [CrossRef]
- Xu, W.; Subudhi, P.K.; Crasta, O.R.; Rosenow, D.T.; Mullet, J.E.; Nguyen, H.T. Molecular mapping of QTLs conferring staygreen in grain sorghum (Sorghum bicolor L. Moench). Genome 2000, 43, 461–469. [Google Scholar] [CrossRef]
- Kebede, H.; Subudhi, P.K.; Rosenow, D.T.; Nguyen, H.T. Quantitative trait loci influencing drought tolerance in grain sorghum (Sorghum bicolor L. Moench). Theor. Appl. Genet. 2001, 103, 266–276. [Google Scholar] [CrossRef]
- Ayalew, K.; Adugna, A.; Fetene, M.; Sintayehu, S. Evaluation of bio-physiological and yield responses of stay-green QTL introgression sorghum lines to post-flowering drought stress. Afr. Crop Sci. J. 2018, 26, 447–462. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahman, M.; El-Sayed, M.; Jogaiah, S.; Burritt, D.J.; Tran, L.P. The “STAY-GREEN” trait and phytohormone signaling networks in plants under heat stress. Plant Cell Rep. 2017, 36, 1009–1025. [Google Scholar] [CrossRef] [PubMed]
- Ngugi, K.; Kimani, W.; Kiambi, D.; Mutitu, E.W. Improving Drought Tolerance in Sorghum bicolor L. Moench: Mar ker-Assisted transfer of the Stay-Green Quantitative Trait Loci (Q T L) from a Characterized Donor Source into a Local Farmer Variety. Int. J. Sci. 2013, 1, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Gimode, D.; Odeny, D.A.; de Villiers, E.P.; Wanyonyi, S.; Dida, M.M.; Mneney, E.E.; Muchugi, A.; Machuka, J.; de Villiers, S.M. Identification of SNP and SSR Markers in Finger Millet Using Next Generation Sequencing Technologies. PLoS ONE 2016, 11, e0159437. [Google Scholar] [CrossRef] [Green Version]
- Mammadov, J.; Aggarwal, R.; Buyyarapu, R.; Kumpatla, S. SNP markers and their impact on plant breeding. Int. J. Plant Genom. 2012, 2012, 728398. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Babu, R.; Hearne, S.; Olsen, M. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Overview of the technology and its application in crop improvement. Mol. Breed. 2013, 33, 1–14. [Google Scholar] [CrossRef]
- Thomson, M.J. High-throughput SNP genotyping to accelerate crop improvement. Plant Breed Biotech. 2014, 2, 195–212. [Google Scholar] [CrossRef]
- Meade, F.; Byrne, S.; Griffin, D.; Kennedy, C.; Mesiti, F.; Dan Milbourne, D. Rapid Development of KASP Markers for Disease Resistance Genes Using Pooled Whole-Genome Re-sequencing. Potato Res. 2020, 63, 57–73. [Google Scholar] [CrossRef]
- Kiranmayee, K.N.S.U.; Hash, C.T.; Sivasubramani, S.; Ramu, P.; Amindala, B.P.; Rathore, A.; Kishor, P.B.K.; Gupta, R.; Deshpande, S.P. Fine-Mapping of Sorghum Stay-Green QTL on Chromosome10 Revealed Genes Associated with Delayed Senescence. Genes 2020, 11, 1026. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, S.; Rakshit, S.; Manasa, K.G.; Pandey, S.; Gupta, R. Genomic Approaches for Abiotic Stress Tolerance in Sorghum. In the Sorghum Genome; Springer: Cham, Switzerland, 2016; pp. 169–187. [Google Scholar]
- Meru, G.M. Genotyping BC3F2 Populations of Four Ethiopian Sorghum Varieties for Stay Green QTLs Introgression through Marker Assisted Selection with SSRs. Master’s Thesis, Kenyata University, Nairobi, Kenya, 2010. [Google Scholar]
- International Board for Plant Genetic Resources (IBPGR); International Crops Research Institute for the Semi-Arid Tropics (ICRISAT). Descriptors for Sorghum [Sorghum bicolor (L.) Moench]; IBPGR: Rome, Italy; ICRISAT: Patancheru, India, 1993. [Google Scholar]
- Burow, G.; Chopra, R.; Sattler, S.; Burke, J.; Acosta-Martinez, V.; Xin, Z. Deployment of SNP (CAPS and KASP) markers for allelic discrimination and easy access to functional variants for brown midrib genes bmr6 and bmr12 in Sorghum bicolor. Mol. Breed. 2019, 39, 115. [Google Scholar] [CrossRef]
- Da Silva, M.J.; Ribeiro, P.C.O.; Silva, R.A.; Silva, K.J.; Schaffert, R.E.; Parrella, R.A.C. Multi-trait selection of sweet sorghum (Sorghum bicolor (L.) Moench) genotypes for bioenergy production. J. Bioenergy Food Sci. 2020, 7, e2952020. [Google Scholar] [CrossRef]
- Schertz, K.F.; Clark, L.E. Controlling dehiscence with plastic bags for hand crosses in Sorghum 1. Crop Sci. 1967, 7, 540–542. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Park, S.; Kim, Y.; Wang, S.; Yoo, S.; Hörtensteiner, S.; Paek, N. Arabidopsis STAY-GREEN2 Is a Negative Regulator of Chlorophyll Degradation during Leaf Senescence. Mol. Plant 2014, 7, 1288–1302. [Google Scholar] [CrossRef] [Green Version]
- Sukumaran, S.; Li, X.; Xianran Li, X.; Zhu, C.; Bai, G.; Perumal, R.; Tuinstra, M.R.; Prasad, P.V.; Mitchell, S.E.; Tesso, T.T.; et al. QTL Mapping for Grain Yield, Flowering Time, and Stay-Green Traits in Sorghum with Genotyping-by-Sequencing Markers. Crop Sci. 2016, 56, 1429–1442. [Google Scholar] [CrossRef]
- Kimani, W.; Ngugi, K.; Kiambi, D.; MutituI, E.W.; De Villiers, S. Marker-assisted introgression of stay-green trait in a Kenyan sorghum variety. E. Afr. Agric. For. J. 2012, 78, 109–115. [Google Scholar]
- Reddy, N.R.; Ragimasalawada, M.; Sabbavarapu, M.M.; Nadoor, S.; Patil, J.V. Detection and va Perrierlidation of stay-green QTL in post-rainy sorghum involving widely adapted cultivar, M35-1 and a popular stay-green genotype B35. BMC Genom. 2014, 15, 909. [Google Scholar] [CrossRef] [Green Version]
- Kamal, N.M.; Gorafi, Y.S.A.; Ghanim, A.M.A. Performance of Sorghum stay-green introgression lines under post-flowering drought. Int. J. Plant Res. 2017, 7, 65–74. [Google Scholar]
- Kholova, J.; Deshpande, S.; Madhusudhana, R.; Blummel, M.; Borrell, A.; Hammer, G. Improving Post-Rainy Sorghum Varieties to Meet the Growing Grain and Fodder Demand in India—Phase 2; Australian Centre for International Agricultural Research: Canberra, Australia, 2019; p. 29.
- Adu-Boakyewaa, G.A.; Badu-Apraku, B.; Akromah, R.; Garcia-Oliveira, A.L.; Awuku, F.J.; Gedil, M. Genetic diversity and population structure of early-maturing tropical maize inbred lines using SNP markers. PLoS ONE 2019, 14, e021481. [Google Scholar]
- Nelimor, C.; Badu-Apraku, B.; Garcia-Oliveira, A.L.; Tetteh, A.; Paterne, A.; N’guetta, A.S.; Gedil, M. Genomic Analysis of Selected Maize Landraces from Sahel and Coastal West Africa Reveals Their Variability and Potential for Genetic Enhancement. Genes 2020, 11, 1054. [Google Scholar] [CrossRef] [PubMed]
- Platten, J.D.; Cobb, J.N.; Zantua, R.E. Criteria for evaluating molecular markers: Comprehensive quality metrics to improve marker-assisted selection. PLoS ONE 2019, 14, e0210529. [Google Scholar] [CrossRef] [Green Version]
- Azzouz-Olden, F.; Hunt, A.G.; Dinkins, R. Transcriptome analysis of drought-tolerant sorghum genotype SC56 in response to water stress reveals an oxidative stress defense strategy. Mol. Biol. Rep. 2020, 47, 3291–3303. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| S.No. | Physical_Mbp | SNP Ids | Alleles | Donor Allele | Alternate Allele | Remarks/Possible |
|---|---|---|---|---|---|---|
| 1 | S2_56112177 | snpSB0035 | C/T | C | T | grain types |
| 2 | S2_56918230 | snpSB0037 | C/T | C | T | |
| 3 | S2_56918233 | snpSB0038 | A/T | A | T | |
| 4 | S2_57495274 | snpSB0039 | A/G | A | G | PCD triggering |
| 5 | S2_59000770 | snpSB0040 | C/T | C | T | |
| 6 | S2_59047449 | snpSB0041 | C/G | C | G | |
| 7 | S2_59255001 | snpSB0042 | T/C | T | C | |
| 8 | S2_59281892 | snpSB0043 | T/C | T | C | |
| 9 | S2_59472716 | snpSB0044 | A/T | A | T | |
| 10 | S2_59821923 | snpSB0049 | G/A | G | A | N2 mobilisation/ SAG/SGR1 |
| 11 | S2_60059010 | snpSB0053 | A/G | A | G | |
| 12 | S2_60098184 | snpSB0054 | G/A | G | A | |
| 13 | S2_61811307 | snpSB0072 | G/A | G | A | APETALA2 and EREBPs |
| 14 | S2_62145285 | snpSB0075 | A/G | A | G | Recurrent parent-specific |
| 15 | S2_62155735 | snpSB0076 | T/A | T | A | Heat shock domain |
| 16 | S2_62155778 | snpSB0077 | T/C | T | C | |
| 17 | S2_62378269 | snpSB0080 | C/T | C | T | Recurrent parent-specific |
| 18 | S2_63375987 | snpSB0083 | T/C | T | C | Chloroplast precursor/ubiquitionus |
| 19 | S2_63690795 | snpSB0087 | A/G | A | G | |
| 20 | S2_65453155 | snpSB0089 | C/A | C | A | SGR2 |
| 21 | S2_67306935 | snpSB0091 | A/C | A | C | panicle compactness |
| 22 | S2_67357372 | snpSB0092 | C/G | G | C | Salt responsive |
| 23 | S2_67423749 | snpSB0093 | A/T | A | T | |
| 24 | S2_67664290 | snpSB0094 | T/C | T | C | |
| 25 | S2_67710384 | snpSB0095 | A/G | A | G | |
| 26 | S2_69739036 | snpSB0098 | C/G | G | C | Aspartic proteases through SA |
| 27 | S2_69859850 | snpSB0099 | C/T | C | T | |
| 28 | S2_70523721 | snpSB0101 | C/G | C | G | SGR3 |
| 29 | S2_71360153 | snpSB0102 | A/G | A | G | |
| 30 | S2_71419274 | snpSB0103 | C/G | G | C |
| Population | PH | STG | NGLM | PL | PW | EX | GWT |
|---|---|---|---|---|---|---|---|
| B35*Wahi | 117.4 c | 4.83 ab | 2.167 ef | 32.22 f | 7.5 fg | 2.5 b | 0.096 e |
| S35*Hakika | 135 defg | 5.5 cd | 2 de | 30.25 e | 7.25 ef | 2 a | 0.08 d |
| B35*NACO | 144.1 gh | 4.67 a | 3 g | 24.92 d | 8.17 g | 3 c | 0.127 f |
| B35*Macia | 132.7 def | 6.67 f | 1 bc | 23.88 d | 5.07 cd | 3 c | 0.027 a |
| B35*Seguifa | 141.7 fgh | 5 abc | 2.833 fg | 20.75 c | 6.58 e | 2.83 bc | 0.11 e |
| S35*Pato | 146.4 h | 6.17 ef | 1 bc | 21.75 c | 8.17 g | 2.5 b | 0.112 e |
| B35 DP | 88 a | 8 g | 0.33 ab | 17.17 ab | 3.5 a | 3 c | 0.013 a |
| Pato RP | 131.3 de | 5 abc | 1.67 cde | 15.83 a | 6.67 e | 2.67 bc | 0.064 c |
| Seguifa RP | 139.7 efgh | 5.33 bcd | 1.67 cde | 21.33 c | 5.67 d | 2.67 bc | 0.067 cd |
| Hakika RP | 131.2 de | 5.33 bcd | 1 bc | 29.17 e | 5.5 d | 3 c | 0.028 a |
| Macia RP | 113.4 bc | 7.67 g | 0.0 a | 21.33 c | 4 ab | 3 c | 0.019 a |
| NACO RP | 113.8 bc | 5.67 de | 1.33 cd | 21.47 c | 5.13 cd | 3 c | 0.047 b |
| S35 DP | 104.5 b | 5.67 de | 2 de | 18.33 b | 4 ab | 3 c | 0.024 a |
| Wahi RP | 128.3 d | 5 abc | 1 bc | 30.17 e | 4.5 bc | 3 c | 0.021 a |
| SE | 2.72 | 0.16 | 0.22 | 0.5 | 0.2 | 0.11 | 4.43 |
| LSD | 5.37 | 0.32 | 0.43 | 0.98 | 0.4 | 0.33 | 8.72 |
| CV (%) | 6.5 | 8.4 | 43.2 | 6.3 | 10.5 | 11.7 | 22.3 |
| NS | SNP Markers | H. Allele | DP Alleles | RP Alleles | NS | SNP Markers | H. Allele | DP Alleles | RP Alleles | |
|---|---|---|---|---|---|---|---|---|---|---|
| BC2F1 (B35*Wahi) | BC2F1 (B35*Seguifa) | |||||||||
| 82 | snpSB0102 | G/A | AA | GA | 567 | snpSB0103 | G/C | CC | CC | |
| 90 | snpSB0102 | G/A | AA | GA | 569 | snpSB0103 | G/C | CC | CC | |
| 94 | snpSB0102, | G/A, | AA, | GA, | 581 | snpSB0103 | G/C | CC | CC | |
| BC2F1 (S35*Hakika) | BC2F1 (S35*Pato) | |||||||||
| 127 | snpSB0103 | G/C | CC | CC | 595 | snpSB0049, | G/A, | GG, | AA, | |
| 131 | snpSB0102, | G/A, | AA, | GA, | 601 | snpSB0103 | G/C | CC | CC | |
| 138 | snpSB0103 | G/C | CC | CC | 610 | snpSB0049, | G/A, | GG, | CC | |
| BC2F1 (NACO Mtama 1) | 624 | snpSB0102 | G/A | AA | AA | |||||
| 213 | snpSB0103 | G/C | CC | CC | snpSB0103 | G/C | GG | CC | ||
| 276 | snpSB0103 | G/C | CC | CC | 645 | snpSB0102 | G/A | AA | AA | |
| 304 | snpSB0102 | G/A | AA | AA | 656 | snpSB0103 | G/C | CC | CC | |
| 307 | snpSB0102, | G/A, | AA, | AA, | 659 | snpSB0102 | G/A | AA | AA | |
| 319 | snpSB0103 | G/C | CC | CC | snpSB0103 | G/C | CC | CC | ||
| 320 | snpSB0103 | G/C | CC | CC | 663 | snpSB0103 | G/C | CC | CC | |
| BC2F1 (B35* Macia) | 668 | snpSB0103 | C/T,G/C | CC | CC | |||||
| 347 | snpSB0103 | G/C | CC | CC | 671 | snpSB0103 | G/C | CC | CC | |
| 348 | snpSB0102, | G/A, | AA, | AA, | 679 | snpSB0103 | G/C | CC | CC | |
| 352 | snpSB0103 | G/C | CC | CC | 685 | snpSB0102, | G/A, | AA, | AA, | |
| 391 | snpSB0103 | G/C | CC | CC | snpSB0103 | G/C | CC | CC | ||
| 395 | snpSB0103 | G/C | CC | CC | 703 | snpSB0102 | G/A | AA | AA | |
| SNP Markers | FA | QTL | Physical Location | Role |
|---|---|---|---|---|
| snpSB00039 | A | STG 3A | S2_57495274 | Retains STG and triggers PCD |
| snpSB00040 | C | STG 3A | S2_59000770 | Retains STG and triggers PCD |
| snpSB00041 | C | STG 3A | S2_59047449 | Retains STG and triggers PCD |
| snpSB00043 | T | STG 3A | S2_59281892 | Retains STG and triggers PCD |
| snpSB00044 | A | STG 3A | S2_59472716 | Retains STG and triggers PCD |
| snpSB00049 | G | STG 3A | S2_59821923 | Retains STG and enhance grain yield |
| snpSB00053 | A | STG 3A | S2_60059010 | Retains STG and enhance grain yield |
| snpSB00076 | T | STG 3A | S2_62155735 | Retains STG and regulate heat shock domain |
| snpSB00077 | T | STG 3A | S2_62155778 | Retains STG and regulate heat shock domain |
| snpSB00083 | T | Interval | S2_63375987 | Retains STG and regulates chloroplast precursors |
| snpSB00087 | A | Interval | S2_63690795 | Retains STG and regulates chloroplast precursors |
| snpSB00089 | C | STG 3B | S2_65453155 | Retains STG and enhances grain yield |
| snpSB00093 | A | STG 3B | S2_67423749 | Retains STG and regulates salt |
| snpSB00094 | T | STG 3B | S2_67664290 | Retains STG and regulates salt |
| snpSB00095 | A | STG 3B | S2_67710384 | Retains STG and regulates salt |
| snpSB000101 | C | STG 3B | S2_70523721 | Retains STG and enhances grain yield |
| snpSB00102 | A | STG 3B | S2_71360153 | Retains STG and enhances grain yield |
| snpSB00103 | G | STG 3B | S2_71419274 | Retains STG and enhances grain yield |
| Genotype | SNP Markers | FA | No. of FA | Genotype | SNP Markers | FA | No. of FA |
|---|---|---|---|---|---|---|---|
| BC2F3 (B35*NACO) | BC2F3 (B35*Seguifa) | ||||||
| NA241A | snSB00049 | GG | 115 | SE408 | snSB00049 | GG | 55 |
| snSB00072 | AA | 6 | snSB00089 | CC | 73 | ||
| snSB00089 | CC | 124 | snSB00101 | CC | 73 | ||
| snSB000102 | AA | 123 | SE438 | snSB00049 | GG | 23 | |
| NA241B | snSB00049 | GG | 113 | snSB00053 | AA | 51 | |
| snSB00101 | CC | 122 | snSB000102 | AA | 52 | ||
| NA316A | snSB00049 | GG | 116 | Seguifa | snSB00049 | GG | 3 |
| snSB00053 | AA | 120 | (Recurrent P) | snSB00053 | AA | 3 | |
| snSB00101 | CC | 121 | snSB000102 | AA | 3 | ||
| NA316B | snSB00049 | GG | 70 | B35 | snSB00042 | TT | 2 |
| snSB00053 | AA | 115 | (Donor P) | snSB00049 | GG | 2 | |
| snSB000102 | AA | 115 | snSB00101 | CC | 2 | ||
| NA316C | snSB00049 | GG | 92 | snSB000102 | AA | 2 | |
| snSB00053 | AA | 122 | snSB00103 | GG | 2 | ||
| snSB00072 | AA | 27 | Wahi | snSB00049 | GG | 3 | |
| snSB000102 | AA | 121 | snSB00101 | CC | 2 | ||
| NACO Mtama 1 | snSB00053 | AA | 3 | ||||
| (Recurrent P) | snSB00054 | GG | 3 | ||||
| SNP Markers | FA | No. of FA | QTL | Physical Location | Function |
|---|---|---|---|---|---|
| snpSB0042 | T | 2 | STG 3A | S2_59255001 | Retain STG and PCD triggering |
| snpSB00049 | G | 584 | STG 3A | S2_59821923 | Retain STG and N mobilisation |
| snpSB00053 | A | 707 | STG 3A | S2_60059010 | Retain STG and N mobilisation |
| snpSB00054 | G | 709 | STG 3A | S2_60098184 | Retain STG and N mobilisation |
| snpSB00072 | A | 105 | STG 3A | S2_61811307 | EREBPs |
| snpSB00089 | C | 729 | STG 3B | S2_65453155 | Retain STG |
| snpSB00101 | C | 728 | STG 3B | S2_70523721 | Retain STG |
| snpSB00102 | A | 727 | STG 3B | S2_71360153 | Retain STG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mwamahonje, A.; Eleblu, J.S.Y.; Ofori, K.; Feyissa, T.; Deshpande, S.; Garcia-Oliveira, A.L.; Bohar, R.; Kigoni, M.; Tongoona, P. Introgression of QTLs for Drought Tolerance into Farmers’ Preferred Sorghum Varieties. Agriculture 2021, 11, 883. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11090883
Mwamahonje A, Eleblu JSY, Ofori K, Feyissa T, Deshpande S, Garcia-Oliveira AL, Bohar R, Kigoni M, Tongoona P. Introgression of QTLs for Drought Tolerance into Farmers’ Preferred Sorghum Varieties. Agriculture. 2021; 11(9):883. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11090883
Chicago/Turabian StyleMwamahonje, Andekelile, John Saviour Yaw Eleblu, Kwadwo Ofori, Tileye Feyissa, Santosh Deshpande, Ana Luísa Garcia-Oliveira, Rajaguru Bohar, Milcah Kigoni, and Pangirayi Tongoona. 2021. "Introgression of QTLs for Drought Tolerance into Farmers’ Preferred Sorghum Varieties" Agriculture 11, no. 9: 883. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11090883