
Allelopathic Effects of Foliar Epichloë Endophytes on Belowground Arbuscular Mycorrhizal Fungi: A Meta-Analysis

1
College of Resources and Environmental Sciences, National Academy of Agriculture Green Development, Key Laboratory of Plant-Soil Interactions, Ministry of Education, China Agricultural University, Beijing 100193, China
2
State Key Laboratory of Grassland Agro-Ecosystems, College of Pastoral Agriculture Science and Technology, Lanzhou University, Lanzhou 730020, China
*
Authors to whom correspondence should be addressed.
Agriculture 2022, 12(11), 1768; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12111768
Submission received: 6 September 2022
/
Revised: 20 October 2022
/
Accepted: 20 October 2022
/
Published: 25 October 2022
(This article belongs to the Special Issue Plant–Soil–Microorganism Interaction in Grassland Agroecosystem)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Many grasses are simultaneously symbiotic with Epichloë fungal endophytes and arbuscular mycorrhizal fungi (AMF). Epichloë endophytes are a group of filamentous fungi that colonize and grow within aerial plant tissues, such as leaves and stems. Infection and hyphal growth of Epichloë endophytes confer fitness advantages to the host plants. In addition to producing fungal alkaloids and altering host metabolic/genetic profiles, it is proven that symbiosis of plants with root/foliar endophytes affects the plant–soil relationship. We propose that the Epichloë presence/infection results in variations of soil and root AMF through allelopathic effects. We performed a meta-analysis that integrated the allelopathic effects of Epichloë endophytes on grass–AMF development. In the pre-symbiotic phase of grass–AMF symbiosis, root exudation from Epichloë-infected plants positively affected AMF growth, whereas the shoot exudates of Epichloë-infected plants inhibited AMF growth. In the symbiotic phase of grass–AMF symbiosis, the Epichloë infection was found to reduce root mycorrhizal colonization in plants. No pattern in the response of soil AMF to Epichloë presence was found. This study should improve our understanding of the impact of Epichloë endophytes on belowground microbial symbionts within the same host plant. Grass–Epichloë–AMF symbiosis may become an important model for studying above–belowground interactions.
1. Introduction
Above–belowground (ABG) interaction is one of the frontiers in studying and understanding the key biochemical and ecological processes in agricultural and natural ecosystems [1,2]. Plants interact with a wide range of microbes—both above- and belowground—that influence plant fitness through nutrient cycling and a wide array of signaling compounds [3,4,5,6]. Grass species are the main components of grassland and agricultural ecosystems, and their fungal symbionts may exert an important effect on plant fitness and biological processes [7,8,9,10]. Many types of grass in Poaceae are symbiotic with Epichloë (Ascomycota, Clavicipitaceae) fungal endophytes [7,9,10]. Epichloë fungal endophytes grow asymptomatically within the intercellular spaces of the aerial tissues of the grass hosts and vertically transmit their progeny through seeds. Epichloë endophytes are a source of secondary metabolites [4,6,11], such as protective fungal alkaloids that are toxic to herbivores and inhibit pathogen colonization, including lolines, indole diterpenes, ergot alkaloids, and peramines. The infection and growth of Epichloë endophytes modify plant immune and metabolic profiles to establish a hyphal network within aerial tissues to promote plant persistence and fitness [10,12].
Epichloë-infected grass plants may offer a competitive advantage to non-infected grass by influencing the root-associated (bulk soil, rhizosphere soil, and root endosphere) microbes [13,14,15]. A previous study suggested that aboveground Epichloë endophytes also generate shifts and variations in belowground biochemical processes and components [8]. Arbuscular mycorrhizal fungi (AMF), belonging to the subphylum Glomeromycotina, can form complex associations with host plants and play important roles in promoting plant fitness and productivity [5]. The symbiosis of AMF with roots begins with spore germination, and the development processes of pre-symbiotic and symbiotic phases are also determined by the host plants. Previous studies on Epichloë-infected grass plants reported shifts in the soil AMF community structure and root AMF colonization [15,16]. Some studies have assessed the potential pathways of Epichloë infection/presence on the developmental status of grass–AMF symbiosis, including root/Epichloë exudation and shoot litter [17,18,19]. Epichloë infection also changed the root exudation profiles [20,21] and shoot decomposition. However, the Epichloë-facilitated soil microbial processes and the subsequent soil microbial responses that contribute to greater tolerance or resistance to grass hosts are unknown. Determining this may carry significant economic and ecological importance in the development of sustainable agricultural practices [13,22].
Understanding the influences of grass–Epichloë interactions on the plant–soil–microbe interactions is of ecological and agricultural importance in grassland and agricultural ecosystems. Based on the transmission of Epichloë endophytes and AMF, we propose a model that integrates the effects of grass–Epichloë-AMF association. Here, we took complementary approaches to validate the multiple tripartite interactions and the predictions of our model. We reviewed the literature related to the interactions between Epichloë endophytes and AMF. A meta-analysis critically analyzed the potential pathways of Epichloë infection on pre-symbiotic and symbiotic phases of grass–AMF development, including (1) exploring the allelopathic effects of Epichloë endophytes on the pre-symbiotic phase of grass–AMF symbiosis, and (2) accessing the effects of Epichloë infection on the symbiotic phase of belowground grass–AMF symbiosis.
2. Material and Methods
We performed a quantitative analysis of published results about the effects of Epichloë endophyte infection on the pre-symbiotic and symbiotic phases of grass–AMF symbiosis. We obtained and reviewed the related literature reporting the multiple interactions between Epichloë and AMF, and performed a standard meta-analysis. Papers in journals were collected from the Web of Science and the China National Knowledge Infrastructure (CNKI) databases using the keywords “Epichloë/Neotyphodium and mycorrhiza” in May 2022.
These papers were screened based on the following criteria: (1) these fungal endophytes belong to the Epichloë (formerly Neotyphodium) genus, and the host plants of Epichloë endophytes are the cool-season grass species; (2) the experiments had to examine the effects of Epichloë endophyte infection/presence on growth and/or colonization of AMF; (3) the references of these selected papers were also checked to search for further appropriate papers; (4) the selected papers had to include the means of AMF growth and colonization for both the endophyte-infected and endophyte-free grasses. Here a total of 32 papers (21 papers in English and 11 papers in Chinese) were obtained from the databases and references of the selected papers for the subsequent meta-analysis.
The extracted values were obtained from the graphs using GetData Graph Digitizer 2.22 and tables in the selected papers. The mean and standard deviations (= standard errors × or the unreported standard deviations were estimated at 10% of the mean), and the number of repetitions (n) of Epichloë-infected (experimental group) and non-infected (control group) grass plants were extracted from the selected papers. The data were grouped into the following categories: (1) exudation resources (shoot, root, and Epichloë strains) or AMF growth (spore germination and hyphal growth) indices in the pre-symbiotic phase of grass–AMF development, and (2) grass classification (grass genus level) or root/soil AMF (root AMF colonization, root AMF concentration, and length of the extraradical hyphae of soil AMF) in the symbiotic phase of grass–AMF symbiosis. For each group as a categorical variable, this group had to be reported in at least five cases. The AMF growth indices in the pre-symbiotic phase of grass–AMF symbiosis were determined by laboratory experiments, and the root/soil AMF indices in the symbiotic phase of grass–AMF symbiosis were determined by controlled pot and field experiments. The meta-analysis was performed using MetaWin software (version 2.1, an intellectual property of Michael S. Rosenberg, Dean C. Adams and Jessica Gurevitch), and these effects were calculated using the log of the response ratio (lnR) using the following Equation according to a study [23].
where and represent the mean AMF growth and colonization of the experimental and control groups, respectively.
The variance of the natural logarithm of the response ratio (lnR (v)) was approximated using the following Equation according to a study [24].
where and indicate the repetitions of the experimental and control groups, respectively, and and indicate the standard deviations of the experimental and control groups, respectively [24]. The lnR means of the effects of Epichloë infection/presence on AMF growth and root/soil AMF were calculated using the combination of lnR and lnR(v), and lnR(v) was weighted by the inverse variance of lnR for each observation. The 95% bootstrap confidence intervals (95% CI) for the mean lnR were calculated using 9999 iterations of bootstrapping, according to a previous study [24].
3. Results
From 142 articles obtained from the Web of Science (124 articles) and CNKI (18 articles) databases, only 32 articles were selected and included in the meta-analysis. A total of 198 cases were obtained from these selected references, including: (1) allelopathic effects of Epichloë fungal endophytes on AMF growth (78 cases in 5 studies), and (2) Epichloë infection on soil and root AMF colonization in plants (120 cases of 31 studies). The present meta-analysis aimed to access whether foliar Epichloë presence/infection could result in some changes to the belowground AMF components (spore germination, hyphal growth, root AMF colonization/concentration, and soil AMF length in pre-symbiotic and symbiotic phases of grass–AMF development) of the grass hosts.
3.1. Does Epichloë Presence Affect the Pre-Symbiotic Phase of Grass-Arbuscular Mycorrhizal Fungi Development?
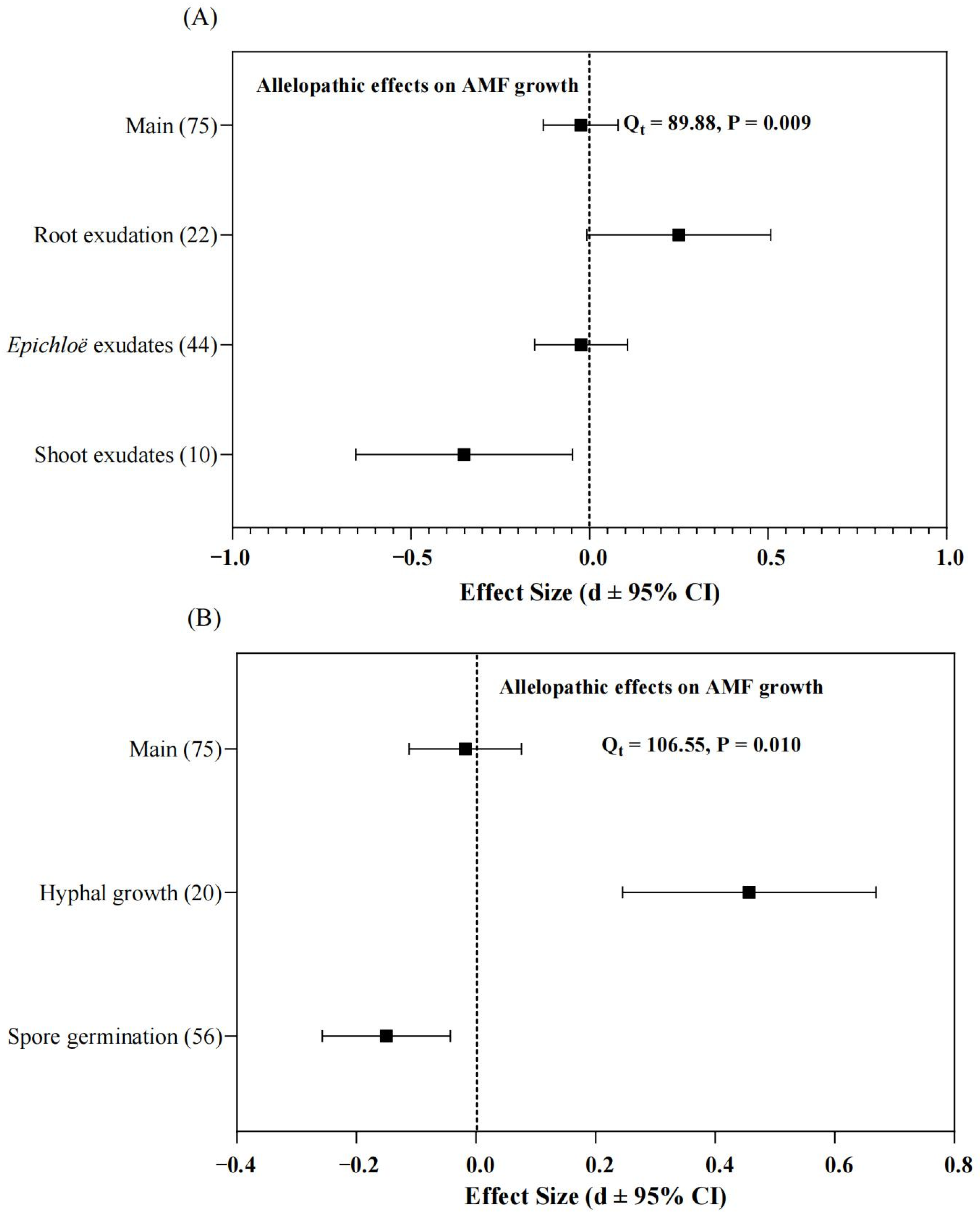
The results of this present meta-analysis showed there were significant (Qt = 89.88, p = 0.009, df = 75) and negative (effect size = −0.025, 95% CI = −0.129 to 0.008) effects of Epichloë fungal endophytes on the pre-symbiotic phase of grass–AMF development within a grass plant (Figure 1A,B). Exudates of Epichloë fungal strains had a weakly negative (effect size = −0.024, 95% CI = −0.154 to 0.106) effect on AMF growth (spore germination, hyphal length, and branch numbers) compared to the control (equal volume of medium) group (Figure 1A). Meanwhile, the shoot exudation from Epichloë-infected grass plants negatively (effect size = −0.351, 95% CI = −0.1655 to −0.048) affected the AMF growth in the pre-symbiotic phase of grass–AMF development than that of non-infected grass plants (Figure 1A). However, root exudation of Epichloë-infected grass plants stimulated (effect size = 0.250, 95% CI = −0.007 to 0.508) AMF growth compared to that of non-infected plants (Figure 1A).
The present meta-analysis indicated there was an overall negative effect (Qt = 106.55, p = 0.010, df = 75, effect size = 0.018, 95% CI = −0.112 to 0.076) of Epichloë-associated (shoot, root and Epichloë strains) exudates on the spore germination and hyphal growth of AMF (Figure 1B). The Epichloë-associated exudates reduced (effect size = 0.150, 95% CI = −0.256 to −0.043) the spore germination of AMF (Figure 1B). However, the Epichloë-associated exudates increased (effect size = 0.457, 95% CI = 0.245 to 0.669) the hyphal growth of AMF (Figure 1B).
3.2. Does Epichloë Infection Affect the Symbiotic Phase of Grass–Arbuscular Mycorrhizal Fungi Development?
The present meta-analysis showed an overall negative effect of Epichloë infection on belowground AMF (Qt = 203.69, p = 0.000, df = 119, effect size = −0.075, 95% CI = −0.128 to −0.022) and root AMF colonization (Qt = 173.21, p = 0.000, df = 102, effect size = −0.087, 95% CI = −0.143 to −0.030) of grass plants (Figure 2A,B). The Epichloë infection reduced root AMF colonization (effect size = −0.088, 95% CI = −0.151 to −0.025) and root AMF concentration (effect size = −0.082, 95% CI = −0.232 to 0.067) of grass plants (Figure 2A). In line with the results of allelopathic effects on AMF growth via root exudations and shoot litter, there was no change (effect size = 0.005, 95% CI = −0.151 to 0.161) in the length of extraradical AMF hyphae in the growing soil of Epichloë-infected versus non-infected plants (Figure 2A).
The present meta-analysis also indicated that the root AMF colonization was significantly (Qt = 203.69, p = 0.000, df = 119) decreased (effect size = −0.087, 95% CI = −0.143 to −0.030) in the roots of Epichloë-infected versus non-infected plants (Figure 2B). In line with the main effects, the same effects were observed in the Lolium (effect size = −0.228, 95% CI = −0.322 to −0.133) and Leymus (effect size = −0.031, 95% CI = −0.149 to 0.088) grass plants (Figure 2B). However, the opposite results were found in Festuca (effect size = 0.012, 95% CI = −0.190 to 0.214) and Achnatherum (effect size = 0.029, 95% CI = −0.126 to 0.183) grass plants (Figure 2B).
3.3. Synthesis of Epichloë Effects on the Development of Grass and Arbuscular Mycorrhizal Fungi
We addressed two perspectives to understand or explain the effects of Epichloë-infection on AMF from changes of root exudation and shoot litter mediated by Epichloë endophytes, and these variations of AMF in root and soil could contribute to the improvement of plant growth and resistance (Figure 3). The root exudation and shoot litter from the Epichloë infected grass plants positively and negatively affected the growth of AMF in the pre-symbiotic phase of grass AMF development compared with the non-infected grass plants (Figure 3). Therefore, there were no obvious changes in the growing soil of Epichloë infected versus non-infected grass plants (Figure 3). Collectively, the presence of foliar Epichloë reduced root colonization by AMF (Figure 3).
4. Discussion
The ABG interaction considers the spatiotemporal scales involved in the interactions between aboveground Epichloë fungal endophytes and belowground AMF within a plant [25]. The spatiotemporal scales could help to understand the coexistence interactions between foliar Epichloë endophyte and root AMF in a plant [25]. Epichloë systemic endophytes are found in the aerial tissues of 20% ~ 30% of all grasses [7], and AMF colonizes the roots of 80% of angiosperms [26], suggesting that these three-way plant–fungal interactions are ecologically common. Based on the transmission modes of Epichloë (vertically transmitted through seeds) and AMF (horizontally colonized grass roots), Epichloë endophytes had higher spatiotemporal priority to modify host metabolic and genetic profiles than co-infected AMF [25]. Here, we addressed two perspectives to investigate how foliar Epichloë endophyte presence/infection affects AMF growth and the development of grass–AMF symbiosis by altering (1) root exudates and (2) shoot litter decomposition (Figure 3).
4.1. The Effects of Epichloë Presence on the Pre-Symbiotic Phase of Grass–AMF Development
The development of AMF symbiosis with plants begins with spore germination, and the related developmental processes depend on the host plants, such as root exudation and plant litter. Root exudates provide chemical signaling to regulate rhizosphere microbial biomass and diversity [3]. Previous meta-analysis and experiment cases showed that the presence of Epichloë endophytes stimulated root exudation [8] and altered the composition of root exudates of the host grass [20,21]. For example, Epichloë-infected grass roots release more carbon (in the form of phenolic compounds and amines) into the rhizosphere through root exudation and volatiles than non-infected grasses [20,27]. Some secondary metabolites (such as lipids, flavonoids, and phenolic compounds) could regulate the pre-symbiotic phase of plant-AMF symbiosis [28,29] at the start of the connection of AMF initial mycelium with plant roots [30]. Our meta-analysis showed that root exudation of Epichloë-infected plants stimulated AMF growth (Figure 1). Additionally, higher AMF diversity in the rhizosphere has been reported in Achnatherum inebrians plants hosting the endophyte Epichloë gansuensis in field experiments under drought stress [14]. However, the shoot litter of Epichloë-infected plants also affected soil microbes compared to non-infected plants [25,31]. Shoot exudates of Epichloë-infected grasses inhibited AMF growth in the present meta-analysis (Figure 1). There was a reasonable explanation that Epichloë fungal alkaloids were present in the litter of Epichloë-infected plants [32,33] and the soil for more than 50 days [34]. Additionally, flavonoids (recognized as inhibitors of AMF [35]) were higher in the roots of Epichloë-infected than in non-infected Lolium multiflorum plants [36]. This study showed that shoot and root exudation had opposite effects on AMF growth; therefore, there was no obvious change in soil AMF in Epichloë-infected versus non-infected grasses (Figure 1, Figure 2 and Figure 3). This is in line with a study showing that aboveground Epichloë-grass associations do not affect belowground AMF symbionts [37]. The shifts of root-associated soil fungal and bacterial community structures in the Epichloë-infected versus non-infected grass plants had been reported in many field and pot cases [8,38,39,40,41]. There were different Epichloë-associated effects on the AMF growth—Epichloë presence reduced the spore germination of AMF strains while promoting the hyphal growth of AMF (Figure 1). This could help explain the variations of soil AMF associated with the presence of an Epichloë endophyte. However, the response mechanism of the root-associated microbiome of grass plants to the infection of Epichloë endophyte is still unclear. Further research should consider the potential mechanisms underlying these selections of soil microbiome by the infection of Epichloë endophytes.
4.2. The Effects of Epichloë Infection on the Symbiotic Phase of Grass–AMF Development
The symbiotic phase in which the AMF mycelium contacts and starts to grow along the root surface and enters the root endosphere is determined by the plant’s nutritional status and defenses [30]. Root mycorrhizal colonization was lower in Epichloë-infected plants than in non-infected plants (Figure 2). In line with two meta-studies, AMF negatively colonized the grass roots of Epichloë-infected plants [8,13]. Epichloë infection promotes auxin content [42], nutrients [42,43], and biomass accumulation [4,44] in the roots of grass hosts. Therefore, we propose that the presence of Epichloë may reduce the beneficial effects of AMF in grass hosts. A previous study supports our suggestion that Epichloë endophytes facilitate the invasion of grass hosts by reducing the benefits derived from other mutualisms [13]. Additionally, the hyphal growth of Epichloë endophytes in aerial tissues also modified the metabolic profiles (promoting the accumulation of flavonoids, benzoic acids, and cinnamic acids) of the roots of grass hosts [36,45], which may inhibit root colonization by AMF. We proposed that the decrease of AMF colonization in the roots of Epichloë-infected versus non-infected grass plants was partly explained by the higher bioactive compounds in the roots of plants infected with Epichloë fungal endophytes. Some indirect evidence has been reported that Epichloë infection altered the root exudation [20,21], root metabolic profiles [36,42], and root/rhizosphere AMF community [14,37], while whether the relationship among root exudation, root metabolic profiles and root/rhizosphere AMF are shifted in the root–soil relations of Epichloë-infected versus non-infected grass plants were sill not clear. Generally, the aboveground Epichloë endophytes could result in changes to belowground AMF, especially AMF growth and root colonization. This meta-analysis also provides insight into the connection between chemical changes in root exudates and shoot litter for plant–soil interactions in ecosystem processes.
5. Conclusions and Further Perspectives
In conclusion, we propose that the presence/infection and hyphal growth of Epichloë endophytes in aboveground tissues negatively affected belowground AMF. Root exudation of Epichloë-infected plants stimulated AMF growth in the pre-symbiotic phase, whereas shoot exudates of Epichloë-infected grass plants inhibited AMF growth. Additionally, AMF negatively colonized grass hosts hosting an Epichloë endophyte. There were no obvious changes in soil AMF. This study should help improve the understanding of the multiple plant–microbe interactions in ABG interactions. This study provides insights and evidence of the capability of the belowground plant and soil processes to be altered by aboveground Epichloë endophytes. There was a positive relationship between the AMF colonization rate and the Epichloë infection rate of the Poa bonariensis population [46]. The Epichloë-infected Hordeum comosum population had greater AMF colonization than non-infected plants [47]. The concentration of aboveground Epichloë endophyte is positively related to AMF concentration in the individual plant [48]. This means that the concentration of both Epichloë fungal endophytes and AMF within a grass plant was decreased compared with grass plants infected with either Epichloë endophytes or AMF. Some studies also reported that AMF colonization also reduced the concentration of Epichloë endophytes and their fungal alkaloid production [49]. Future research should consider (1) how variations in belowground processes from the Epichloë endophytes present feedback to the Epichloë–grass associations; (2) genetic and metabolic profiles of grass hosts to respond to simultaneous infections and environmental stresses; and (3) simultaneous infections on plant growth and defenses and the applications of multiple plant–microbe interactions in sustainable agriculture.
Author Contributions
R.Z. and L.Z. developed the concept; R.Z. collected and analyzed the data, R.Z., L.Z. and X.Z. wrote this manuscript, and all authors revised this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by the China Postdoctoral Science Foundation (2021M703515) and the Fundamental Research Funds for the Central Universities (lzujbky-2022-ey21), Lanzhou University.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Acknowledgments
Thanks to Michael Christensen from AgResearch Ltd., Grasslands Research Centre, New Zealand, for his valuable suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, K.S.; Geisen, S.; Morriën, E.; Snoek, B.L.; van der Putten, W. Network analyses can advance above-belowground ecology. Trends Plant Sci. 2018, 23, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Bastias, D.A.; Martínez-Ghersa, M.A.; Ballaré, C.L.; Gundel, P.E. Epichloë fungal endophytes and plant defenses: Not just alkaloids. Trends Plant Sci. 2017, 22, 939–948. [Google Scholar] [CrossRef]
- De Vries, F.T.; Griffiths, R.I.; Knight, C.G.; Nicolitch, O.; Williams, A. Harnessing rhizosphere microbiomes for drought-resilient crop production. Science 2020, 368, 270–274. [Google Scholar] [CrossRef]
- Schardl, C.L.; Florea, S.; Pan, J.; Nagabhyru, P.; Bec, S.; Calie, P.J. The epichloae: Alkaloid diversity and roles in symbiosis with grasses. Curr. Opin. Plant Biol. 2013, 16, 480–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leuchtmann, A. Systematics, distribution, and host specificity of grass endophytes. Nat. Toxins 1992, 1, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Omacini, M.; Semmartin, M.; Perez, L.I.; Gundel, P.E. Grass-endophyte symbiosis: A neglected aboveground interaction with multiple belowground consequences. Appl. Soil Ecol. 2012, 61, 273–279. [Google Scholar] [CrossRef]
- Kauppinen, M.; Saikkonen, K.; Helander, M.; Pirttilä, A.M.; Wäli, P.R. Epichloë grass endophytes in sustainable agriculture. Nat. Plants 2016, 2, 15224. [Google Scholar] [CrossRef]
- Schardl, C.L.; Leuchtmann, A.; Spiering, M.J. Symbioses of grasses with seedborne fungal endophytes. Annu. Rev. Plant Biol. 2004, 55, 315–340. [Google Scholar] [CrossRef]
- Ju, Y.; Sacalis, J.N.; Still, C.C. Bioactive flavonoids from endophyte-infected blue grass (Poa ampla). J. Agric. Food Chem. 1998, 46, 3785–3788. [Google Scholar] [CrossRef]
- Scott, B.; Green, K.; Berry, D. The fine balance between mutualism and antagonism in the Epichloë festucae-grass symbiotic interaction. Curr. Opin. Plant Biol. 2018, 44, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Perez, L.I.; Gundel, P.E.; Parisi, P.A.G.; Moyano, J.; Fiorenza, J.E.; Omacini, M.; Nunez, M.A. Can seed-borne endophytes promote grass invasion by reducing host dependence on mycorrhizas? Fungal Ecol. 2021, 52, 101077. [Google Scholar] [CrossRef]
- Zhong, R.; Xia, C.; Ju, Y.W.; Zhang, X.X.; Duan, T.Y.; Nan, Z.B.; Li, C.J. A foliar Epichloë endophyte and soil moisture modified belowground arbuscular mycorrhizal fungal biodiversity associated with Achnatherum inebrians. Plant Soil 2021, 458, 105–122. [Google Scholar] [CrossRef]
- Arrieta, A.M.; Iannone, L.J.; Scervino, J.M.; Vignale, M.V.; Novas, M.V. A foliar endophyte increases the diversity of phosphorus-solubilizing rhizospheric fungi and mycorrhizal colonization in the wild grass Bromus auleticus. Fungal Ecol. 2015, 17, 146–154. [Google Scholar] [CrossRef]
- Johnson, L.J.; de Bonth, A.C.M.; Briggs, L.R.; Caradus, J.R.; Finch, S.C.; Fleetwood, D.J.; Fletcher, L.R.; Hume, D.E.; Johnson, R.D.; Popay, A.J.; et al. The exploitation of epichloae endophytes for agricultural benefit. Fungal Divers. 2013, 60, 171–188. [Google Scholar] [CrossRef]
- Wu, R.H.; Liu, H.; Wu, M.; Ren, A.Z.; Gao, Y.B. Effects of Epichloë endophytes of Achnatherum sibiricum on spore germination of arbuscular mycorrhizal fungi. Chin. J. Appl. Ecol. 2018, 29, 4145–4151, (In Chinese with English Abstract). [Google Scholar]
- Novas, M.V.; Iannone, L.J.; Godeas, A.M.; Scervino, J.M. Evidence for leaf endophyte regulation of root symbionts: Effect of Neotyphodium endophytes on the pre-infective state of mycorrhizal fungi. Symbiosis 2011, 55, 19–28. [Google Scholar] [CrossRef]
- Vignale, M.V.; Iannone, L.J.; Scervino, J.M.; Novas, M.V. Epichloë exudates promote in vitro and in vivo arbuscular mycorrhizal fungi development and plant growth. Plant Soil 2018, 422, 267–281. [Google Scholar] [CrossRef]
- Guo, J.Q.; Rebecca, M.; McNear, D.H. Tall fescue cultivar and fungal endophyte combinations influence plant growth and root exudate composition. Front. Plant Sci. 2015, 6, 183. [Google Scholar] [CrossRef] [Green Version]
- Patchett, A.; Newman, J.A. Comparison of plant metabolites in root exudates of Lolium perenne infected with different strains of the fungal endophyte Epichloë festucae var. lolii. J. Fungi 2021, 7, 148. [Google Scholar] [CrossRef] [PubMed]
- Hager, H.; Gailis, M.; Newman, J. Allelopathic effects of Epichloë fungal endophytes: Experiment and meta-analysis. Plant Soil 2022, 1–16. [Google Scholar] [CrossRef]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The meta-analysis of response ratios in experimental ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Gurevitch, J.; Curtis, P.S.; Jones, M.H. Meta-analysis in ecology. Adv. Ecol. Res. 2001, 32, 199–247. [Google Scholar]
- Mack, K.M.L.; Rudgers, J.A. Balancing multiple mutualists: Asymmetric interactions among plants, arbuscular mycorrhizal fungi, and fungal endophytes. Oikos 2008, 117, 310–320. [Google Scholar] [CrossRef]
- Brundrett, M.C. Coevolution of roots and mycorrhizas of land plants. New Phytol. 2002, 154, 275–304. [Google Scholar] [CrossRef] [PubMed]
- Rostás, M.; Cripps, M.G.; Silcock, P. Aboveground endophyte affects root volatile emission and host plant selection of a belowground insect. Oecologia 2015, 177, 487–497. [Google Scholar] [CrossRef]
- Scervino, J.M.; Ponce, M.A.; Erra-Bassells, R.; Vierheilig, H.; Ocampo, J.A.; Godeas, A. Flavonoids exhibit fungal species and genus specific effects on the presymbiotic growth of Gigaspora and Glomus. Mycol. Res. 2005, 109, 789–794. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.N.; Wang, W.X.; Xie, Q.J.; Liu, N.; Liu, L.X.; Wang, D.P.; Zhang, X.W.; Yang, C.; Chen, X.Y.; Tang, D.Z.; et al. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef] [Green Version]
- Souza, T. Handbook of Arbuscular Mycorrhizal Fungi; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Antunes, P.M.; Miller, J.; Carvalho, L.M.; Klironomos, J.N.; Newman, J.A. Even after death the endophytic fungus of Schedonorus phoenix reduces the arbuscular mycorrhizas of other plants. Funct. Ecol. 2008, 22, 912–918. [Google Scholar] [CrossRef] [Green Version]
- Siegrist, J.; McCulley, R.; Bush, L.; Phillips, T. Alkaloids may not be responsible for endophyte-associated reductions in tall fescue decomposition rates. Funct. Ecol. 2010, 24, 460–468. [Google Scholar] [CrossRef]
- Gundel, P.E.; Helander, M.; Garibaldi, L.A.; Vázquez-de-Aldana, B.R.; Zabalgogeazcoa, I.; Saikkonen, K. Direct and indirect effects of the fungal endophyte Epichloë uncinatum on litter decomposition of the host grass, Schedonorus pratensis. Plant Ecol. 2017, 218, 1107–1115. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Hill, N.S. Soil carbon, nitrogen, and ergot alkaloids with short- and long-term exposure to endophyte-infected and endophyte-free tall fescue. Soil Sci. Soc. Am. J. 2005, 69, 404–412. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, P.L.N.; Silva, E.O.; Chagas-Paula, D.A.; Hortolan Luiz, J.H.; Ikegaki, M. Importance and implications of the production of phenolic secondary metabolites by endophytic fungi: A mini-review. Mini Rev. Med. Chem. 2016, 16, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Ponce, M.A.; Bompadre, M.J.; Scervino, J.M.; Ocampo, J.A.; Chaneton, E.J.; Godeas, A.M. Flavonoids, benzoic acids and cinnamic acids isolated from shoots and roots of Italian rye grass (Lolium multiflorum Lam.) with and without endophyte association and arbuscular mycorrhizal fungus. Biochem. Syst. Ecol. 2009, 37, 245–253. [Google Scholar] [CrossRef]
- Slaughter, L.C.; McCulley, R.L. Aboveground Epichloë coenophiala-grass associations do not affect belowground fungal symbionts or associated plant, soil parameters. Microb. Ecol. 2016, 72, 682–691. [Google Scholar] [CrossRef]
- Bell-Dereske, L.; Takacs-Vesbach, C.; Kivlin, S.N.; Emery, S.M.; Rudgers, J.A. Leaf endophytic fungus interacts with precipitation to alter belowground microbial communities in primary successional dunes. FEMS Microb. Ecol. 2017, 93, fix036. [Google Scholar] [CrossRef] [Green Version]
- Ding, N.; Guo, H.; Kupper, J.V.; McNear, D.H. Phosphorus source and Epichloe coenophiala strain interact over time to modify tall fescue rhizosphere microbial community structure and function. Soil Biol. Biochem. 2021, 154, 208125. [Google Scholar] [CrossRef]
- van Hecke, M.M.; Treonis, A.M.; Kaufman, J.R. How does the fungal endophyte Neotyphodium coenophialum affect tall fescue (Festuca arundinacea) rhizodeposition and soil microorganisms? Plant Soil 2005, 275, 101–109. [Google Scholar] [CrossRef]
- Chen, J.; Deng, Y.K.; Yu, X.H.; Wu, G.H.; Gao, Y.B.; Ren, A.Z. Epichloë endophyte infection changes the root endosphere microbial community composition of Leymus chinensis under both potted and field growth conditions. Microb. Ecol. 2022, 1–13. [Google Scholar] [CrossRef]
- Cui, X.L.; Zhang, X.X.; Shi, L.; Christensen, M.J.; Nan, Z.B.; Xia, C. Effects of Epichloë endophyte and transgenerational effects on physiology of Achnatherum inebrians under drought stress. Agriculture 2022, 12, 761. [Google Scholar] [CrossRef]
- Xia, C.; Christensen, M.J.; Zhang, X.X.; Nan, Z.B. Effect of Epichloë gansuensis endophyte and transgenerational effects on the water use efficiency, nutrient and biomass accumulation of Achnatherum inebrians under soil water deficit. Plant Soil 2018, 424, 555–571. [Google Scholar] [CrossRef]
- Zhong, R.; Bastías, D.A.; Zhang, X.X.; Li, C.J.; Nan, Z.B. Vertically transmitted Epichloë systemic endophyte enhances drought tolerance of Achnatherum inebrians host plants through promoting photosynthesis and biomass accumulation. J. Fungi 2022, 8, 512. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, D.P.; Alloush, G.A.; Belesky, D.P. Evidence for chemical changes on the root surface of tall fescue in response to infection with the fungal endophyte Neotyphodium coenophialum. Plant Soil 1998, 205, 1–12. [Google Scholar] [CrossRef]
- Novas, M.V.; Iannone, L.J.; Godeas, A.M.; Cabral, D. Positive association between mycorrhiza and foliar endophytes in Poa bonariensis, a native grass. Mycol. Progress 2008, 8, 75. [Google Scholar] [CrossRef]
- Casas, C.; Gundel, P.E.; Deliens, E.; Iannone, L.J.; Martinez, G.G.; Vignale, M.V.; Schnyder, H. Loss of fungal symbionts at the arid limit of the distribution range in a native Patagonian grass-Resource eco-physiological relations. Funct. Ecol. 2022, 36, 583–594. [Google Scholar] [CrossRef]
- Liu, H.; Wu, M.; Liu, J.M.; Qu, Y.B.; Gao, Y.B.; Ren, A.Z. Tripartite interactions between endophytic fungi, arbuscular mycorrhizal fungi, and Leymus chinensis. Microb. Ecol. 2020, 79, 98–109. [Google Scholar] [CrossRef]
- Liu, Q.H.; Parsons, A.J.; Xue, H.; Fraser, K.; Ryan, G.D.; Newman, J.A.; Rasmussen, S. Competition between foliar Neotyphodium lolii endophytes and mycorrhizal Glomus spp. fungi in Lolium perenne depends on resource supply and host carbohydrate content. Funct. Ecol. 2011, 25, 910–920. [Google Scholar] [CrossRef]
Figure 1.
The cumulative effect size of allelopathic effects of Epichloë endophytes on the pre-symbiotic phase of the grass and arbuscular mycorrhizal fungi (AMF) development among (A) different allelopathic (exudates from root, shoot, and Epichloë strains) effects, or (B) AMF germination and hyphal growth. Squares indicate the mean values and bars denote the 95% confidence interval (CI).
Figure 1.
The cumulative effect size of allelopathic effects of Epichloë endophytes on the pre-symbiotic phase of the grass and arbuscular mycorrhizal fungi (AMF) development among (A) different allelopathic (exudates from root, shoot, and Epichloë strains) effects, or (B) AMF germination and hyphal growth. Squares indicate the mean values and bars denote the 95% confidence interval (CI).

Figure 2.
The cumulative effect size of Epichloë endophytes infection on the symbiotic phase of the grass and arbuscular mycorrhizal fungi (AMF) development among (A) soil and root AMF effects or (B) root AMF colonization of different plant genera. The squares indicate the mean values and the bars denote the 95% confidence interval (CI).
Figure 2.
The cumulative effect size of Epichloë endophytes infection on the symbiotic phase of the grass and arbuscular mycorrhizal fungi (AMF) development among (A) soil and root AMF effects or (B) root AMF colonization of different plant genera. The squares indicate the mean values and the bars denote the 95% confidence interval (CI).

Figure 3.
The synthesis of potential mechanisms for the effects of Epichloë endophytes infection/presence on the developmental processes of grass roots and soil arbuscular mycorrhizal fungi (AMF).
Figure 3.
The synthesis of potential mechanisms for the effects of Epichloë endophytes infection/presence on the developmental processes of grass roots and soil arbuscular mycorrhizal fungi (AMF).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhong, R.; Zhang, L.; Zhang, X. Allelopathic Effects of Foliar Epichloë Endophytes on Belowground Arbuscular Mycorrhizal Fungi: A Meta-Analysis. Agriculture 2022, 12, 1768. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12111768
AMA Style
Zhong R, Zhang L, Zhang X. Allelopathic Effects of Foliar Epichloë Endophytes on Belowground Arbuscular Mycorrhizal Fungi: A Meta-Analysis. Agriculture. 2022; 12(11):1768. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12111768
Chicago/Turabian StyleZhong, Rui, Lin Zhang, and Xingxu Zhang. 2022. "Allelopathic Effects of Foliar Epichloë Endophytes on Belowground Arbuscular Mycorrhizal Fungi: A Meta-Analysis" Agriculture 12, no. 11: 1768. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12111768
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.