
Identification of Drought-Resistant Genes in Shanlan Upland Rice

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
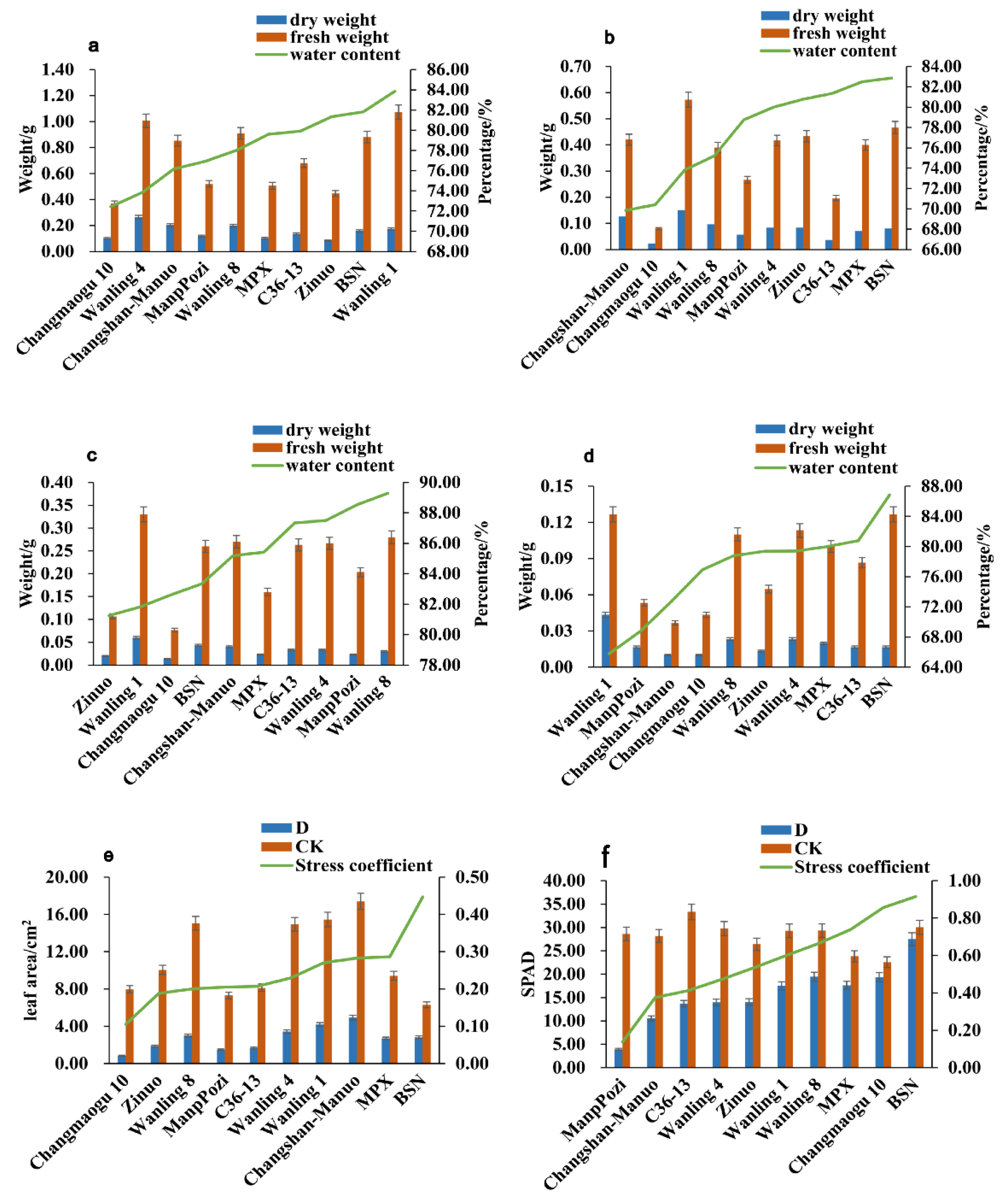
2.2.1. Drought Resistance Phenotype Identification
2.2.2. Library Preparation and RNA Sequencing
2.2.3. Transcriptomic Data and Gene Expression Level Analysis
2.2.4. Screening and Analysis of Differentially Expressed Genes
3. Results
3.1. Screening Drought Resistance of Different Shanlan Upland Rice Varieties
3.2. Sample Collection and cDNA Library Construction for Sequencing
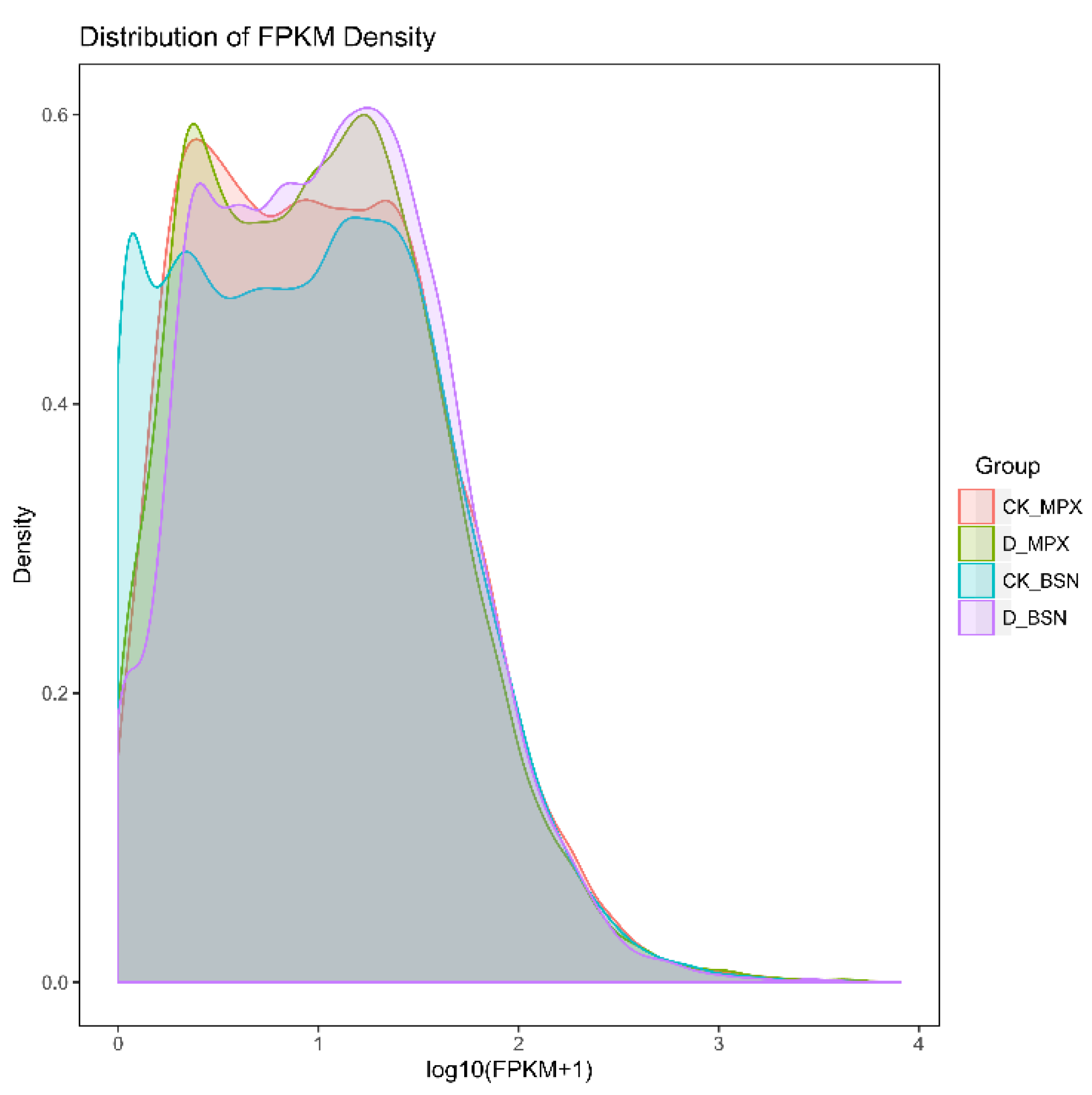
3.3. Transcriptomic Data Quality Analysis
3.4. Gene Expression Levels Analysis
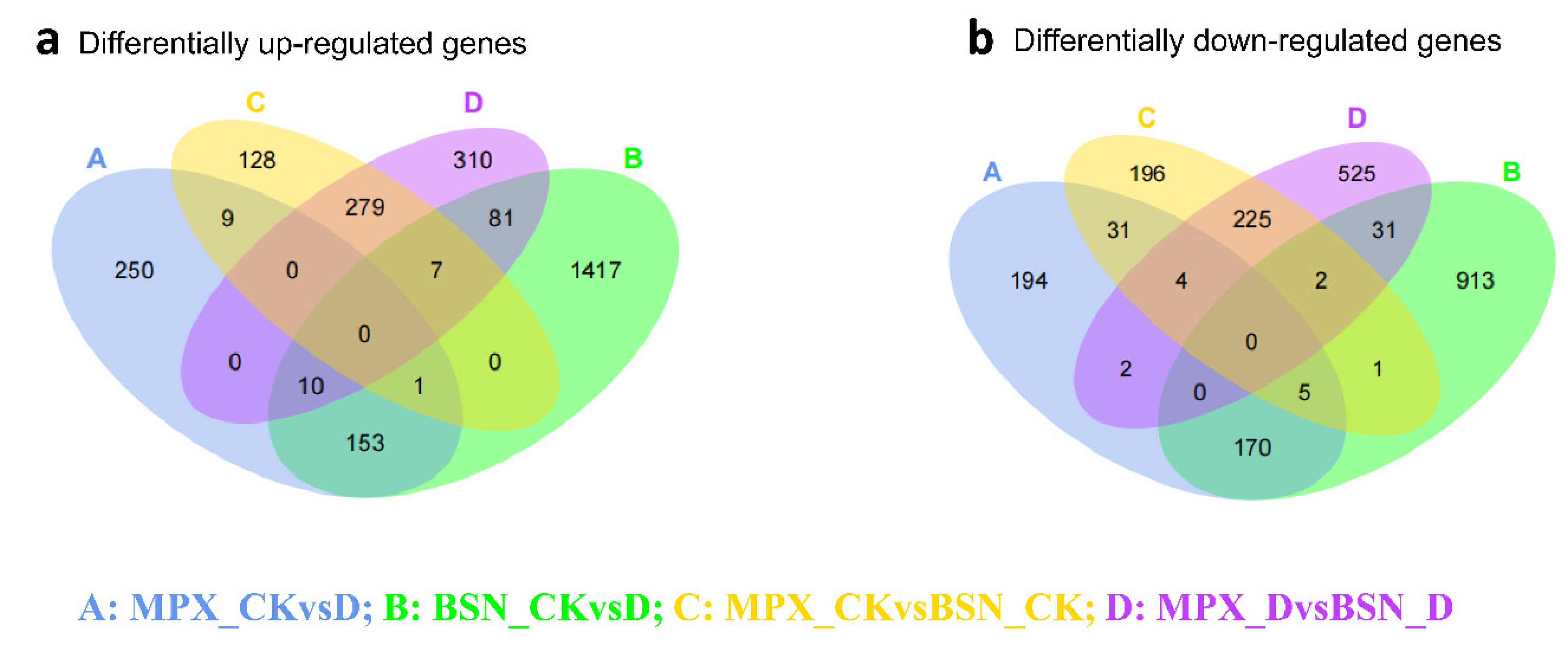
3.5. Comparison of Gene Expression
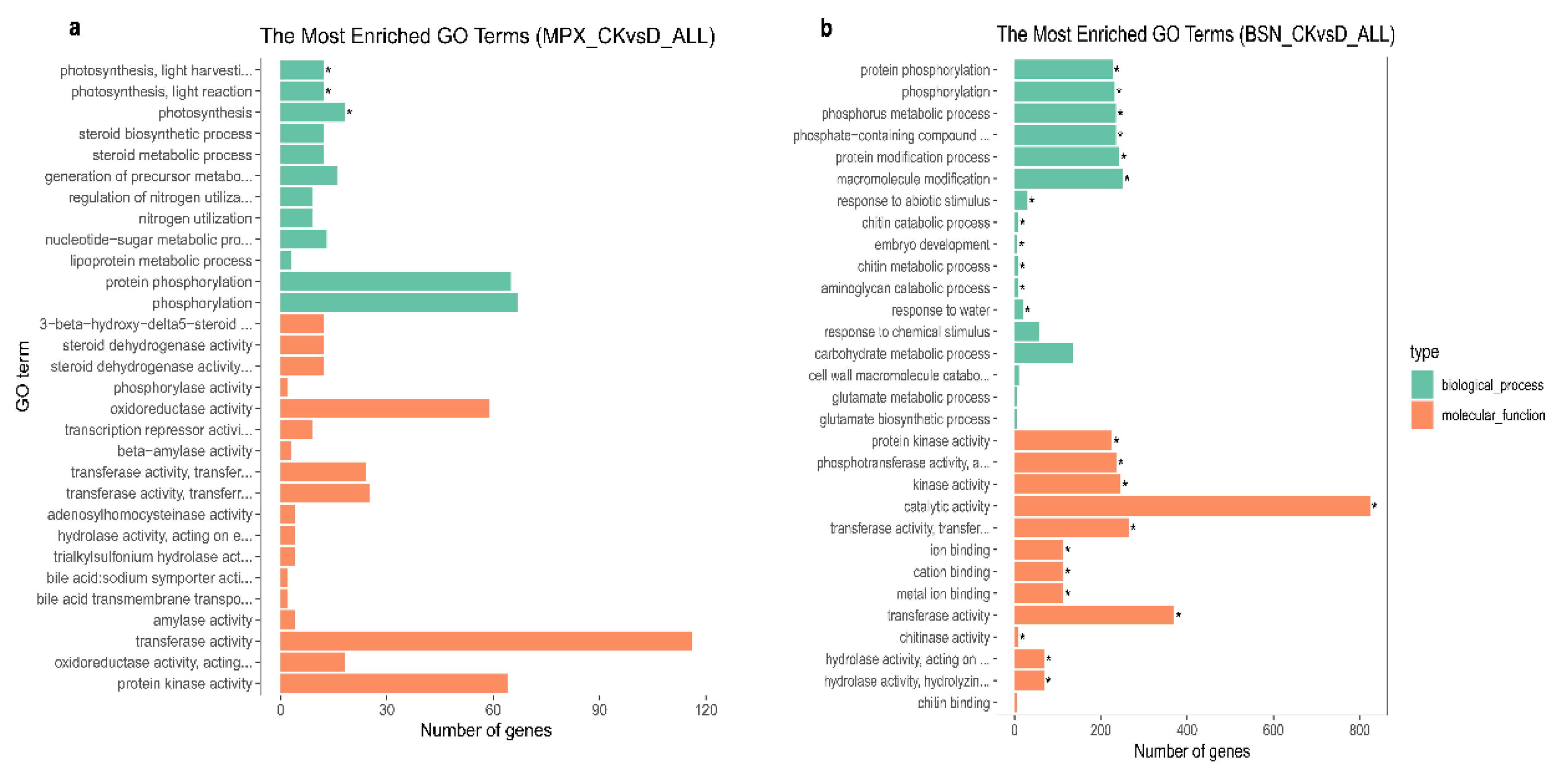
3.6. GO Enrichment Analysis of the DEGs
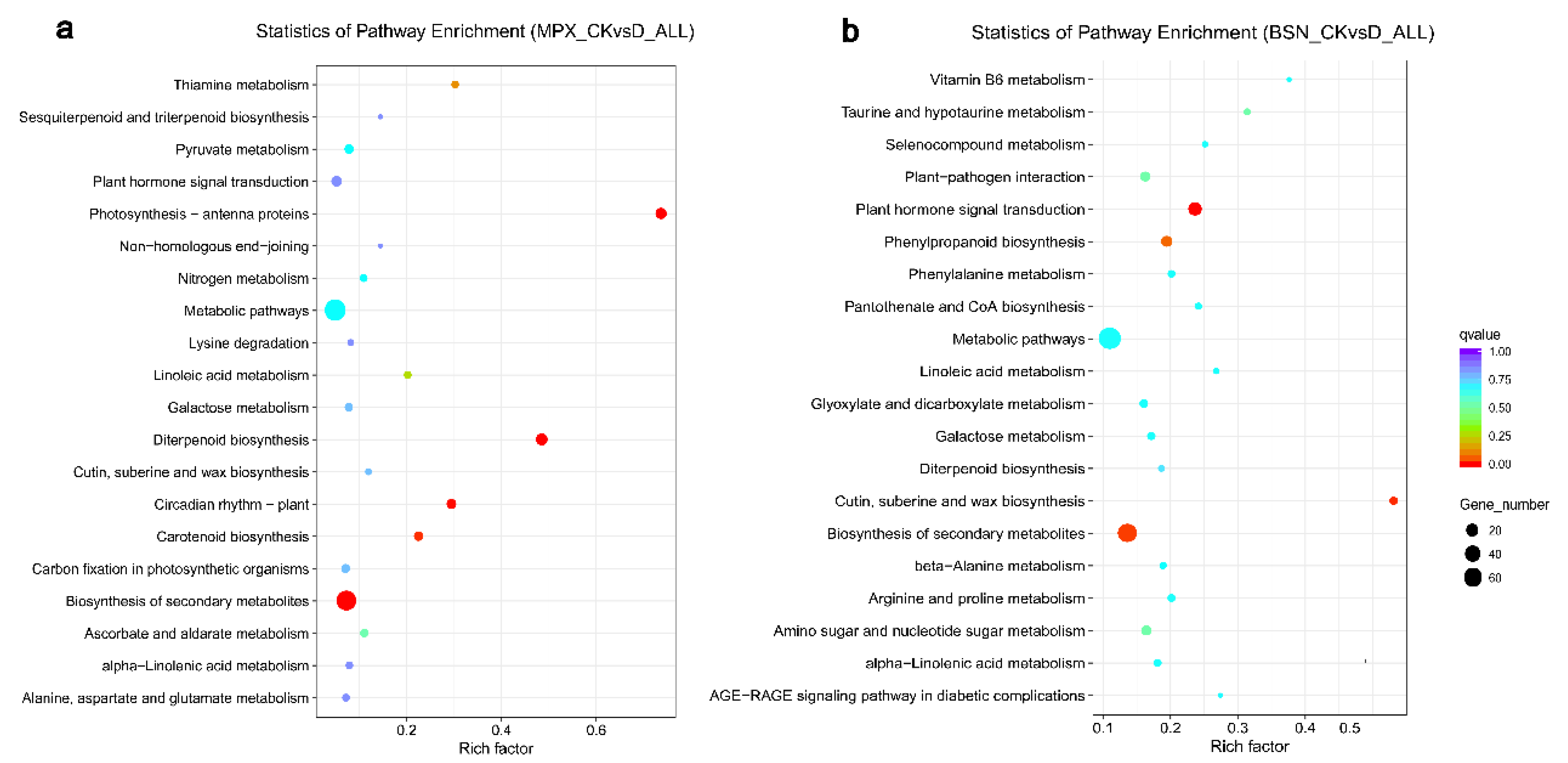
3.7. KEGG Enrichment Analysis of the DEGs
3.8. Differentially Expressed Transcription Factor Analysis
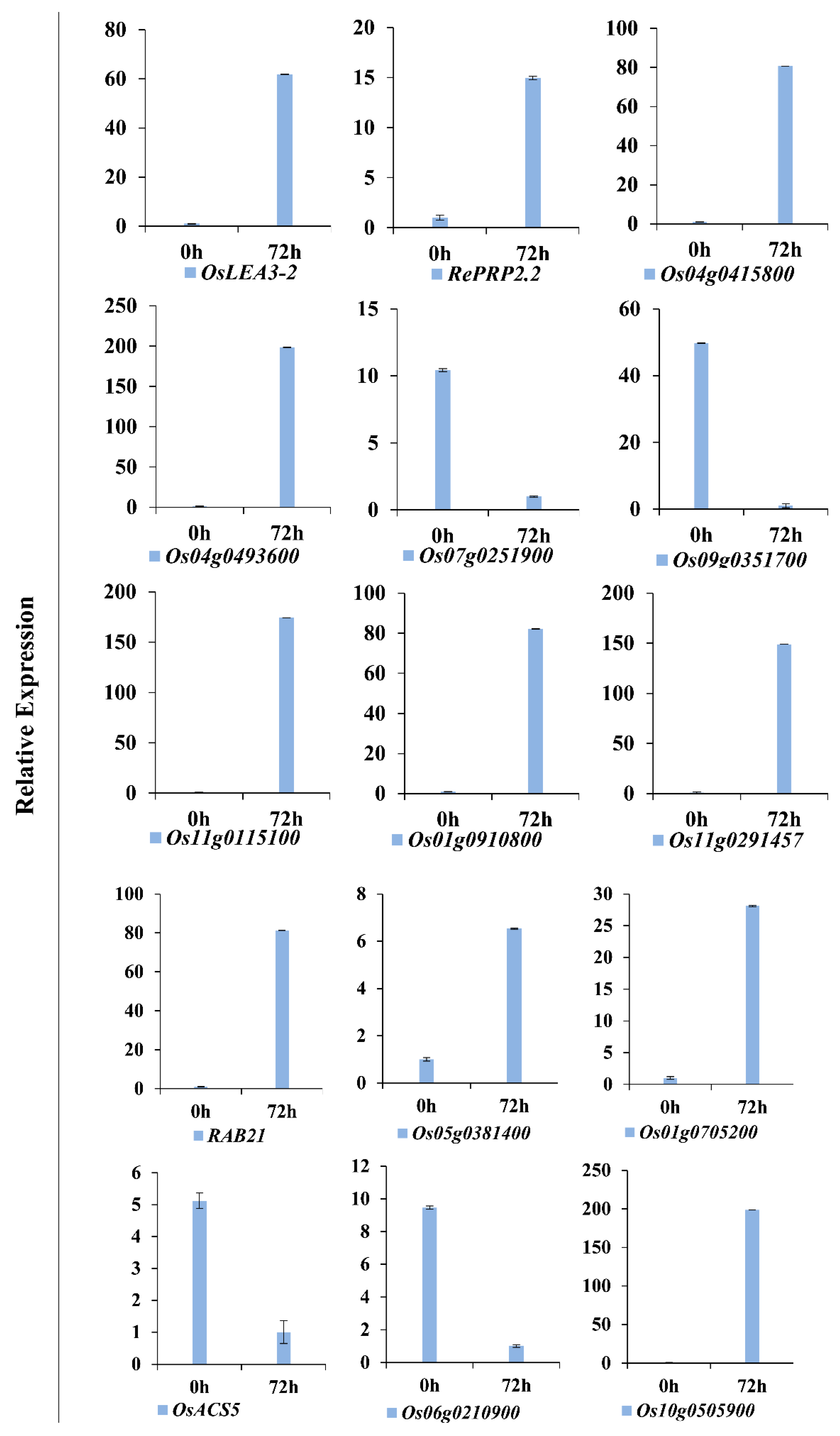
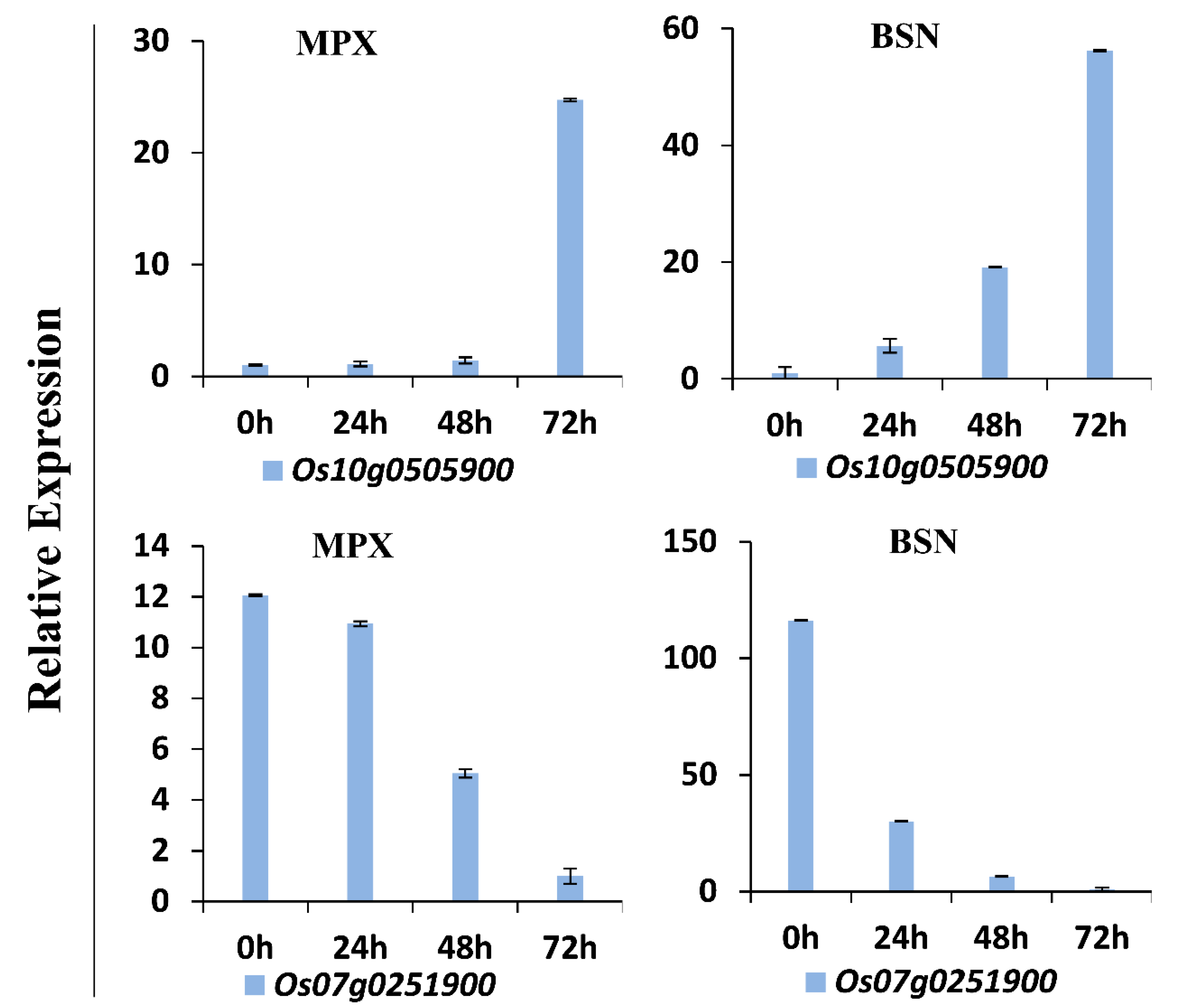
3.9. Verification of DEGs by Quantitative Real-Time PCR
4. Discussions
4.1. Phenotypic Analysis of Shanlan Upland Rice
4.2. Transcriptome Identification and Analysis of Shanlan Upland Rice Varieties
4.3. GO Analysis of Genes Related to Drought Resistance in Shanlan Upland Rice
4.4. KEGG Analysis of Drought Related Genes in Shanlan Upland Rice
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spracklen, D.V. Global warming: China’s contribution to climate change. Nature 2016, 531, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.E. Early climate models successfully predicted global warming. Nature 2020, 578, 45–46. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Oliveira, M.M. Mechanisms underlying plant resilience to water deficits: Prospects for water-saving agriculture. J. Exp. Bot. 2004, 55, 2365–2384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Zhao, Y.; Xu, P.; Yu, D. Microbial diversity of upland rice roots and their influence on rice growth and drought tolerance. Microorganisms 2020, 8, 1329. [Google Scholar] [CrossRef]
- Yuan, N.N.; Wei, X.; Xue, D.Y.; Yang, Q.W. The origin and evolution of upland rice in Li ethnic communities in Hainan province. J. Plant Genet. Resour. 2013, 14, 202–207. [Google Scholar]
- Liu, H.Z.; Ji, C.D. Conservation and utilization of Shanlan upland rice germplasm resources in Hainan province. Chin. J. Trop. Agric. 2016, 36, 49–51. [Google Scholar]
- Huang, C.Y.; Luo, W.Q.; Wang, B.; Yang, X.B. Germplasm resource and conservative model of upland rice (Shanlan rice) in south-central Hainan. Guihaia 2015, 35, 905–912. [Google Scholar]
- Yang, J.; Xu, J.X. Research progress on drought resistance of Shanlan upland rice in Hainan province. Chin. J. Trop. Agric. 2018, 38, 64–68. [Google Scholar]
- Liu, X.X.; Xu, L.X.; Yuan, Q.H. Cloning and sequence biological analysis of HKT2 gene from Shanlan upland rice in Hainan. Guangdong Agric. Sci. 2013, 40, 128–132. [Google Scholar]
- Liu, W.J.; Xu, L.X.; He, M.D.; Wen, Z.Z.; Zhang, J.N.; Yuan, Q.H. The differences in morphological and physiological traits between Shanlan upland rice and cultivated rice under drought stress. J. Trop. Biol. 2014, 5, 260–264. [Google Scholar]
- He, G.L.; Zhao, J.F.; He, M.D.; Ke, Z.; Yuan, Q.H. Effects of cultivation in different mulching patterns on the agronomic traits of Shanlan upland rice and weed control. J. Trop. Biol. 2018, 9, 344–349. [Google Scholar]
- Jo, K.; Kwon, H.B.; Kim, S. Time-series RNA-seq analysis package (TRAP) and its application to the analysis of rice, Oryza sativa L. ssp. Japonica, upon drought stress. Methods 2014, 67, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Ma, X.; Xu, K.; Wang, L.; Liu, H.; Chen, L.; Luo, L. Temporal transcriptomic differences between tolerant and susceptible genotypes contribute to rice drought tolerance. BMC Genom. 2020, 21, 776. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Xie, K.; Yao, J.; Qi, Z.; Xiong, L. A homolog of human ski-interacting protein in rice positively regulates cell viability and stress tolerance. Proc. Natl. Acad. Sci. USA 2009, 106, 6410–6415. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.F.; Li, Y.Y.; Xiao, B.Z. Comparative transcriptome analysis highlights the crucial roles of photosynthetic system in drought stress adaptation in upland rice. Sci. Rep. 2016, 6, 19349. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.J.; Long, Y.; Wang, J.; Zhang, J.W.; Wang, Y.Y.; Li, W.M.; Peng, Y.F.; Yuan, Q.H.; Pei, X.W. De novo transcriptome assembly of common wild rice (Oryza rufipogon griff.) and discovery of drought-response genes in root tissue based on transcriptomic data. PLoS ONE 2015, 10, e131455. [Google Scholar] [CrossRef] [PubMed]
- Yoo, Y.H.; Nalini Chandran, A.K.; Park, J.C.; Gho, Y.S.; Lee, S.W.; An, G.; Jung, K.H. OsPhyB-mediating novel regulatory pathway for drought tolerance in rice root identified by a global RNA-Seq transcriptome analysis of rice genes in response to water deficiencies. Front. Plant Sci. 2017, 8, 580. [Google Scholar] [CrossRef]
- Han, B.; Wang, J.; Li, Y.; Ma, X.; Jo, S.; Cui, D. Identification of quantitative trait loci associated with drought tolerance traits in rice (Oryza sativa L.) under PEG and field drought stress. Euphytica 2018, 214, 1–16. [Google Scholar] [CrossRef]
- Wang, B.; Xu, B.; Liu, Y.; Chen, X.; Liu, J.; Zhi, W.; Xing, Y.; Yang, B.; Li, J.; Chi, M.; et al. Variation of drought resistance of rice genotypes released in different years in China. J. Sci. Food Agric. 2019, 99, 4430–4438. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Zhang, L.; Luo, L.X.; Wang, H.; Guo, T.; Liu, Y.Z.; Zhou, J.Y.; Chen, Z.Q.; Xiao, W.M. Response analysis of rice gene NAL11 to abiotic stresses at the stage of seedling. Acta Agric. Boreali-Sin. 2020, 35, 120–128. [Google Scholar]
- Zhao, S.; Ye, Z.; Stanton, R. Misuse of RPKM or TPM normalization when comparing across samples and sequencing protocols. RNA 2020, 26, 903–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quail, M.A.; Kozarewa, I.; Smith, F.; Scally, A.; Stephens, P.J.; Durbin, R.; Swerdlow, H.; Turner, D.J. A large genome center’s improvements to the Illumina sequencing system. Nat. Methods 2008, 5, 1005–1010. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Cai, W. OsLEA3-2, an abiotic stress induced gene of rice plays a key role in salt and drought tolerance. PLoS ONE 2012, 7, e45117. [Google Scholar] [CrossRef] [Green Version]
- Tseng, I.C.; Hong, C.Y.; Yu, S.M.; Ho, T.H. Abscisic acid- and stress-induced highly proline-rich glycoproteins regulate root growth in rice. Plant Physiol. 2013, 163, 118–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mundy, J.; Chua, N.H. Abscisic acid and water-stress induce the expression of a novel rice gene. EMBO J. 1988, 7, 2279–2286. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, L.J.; Mei, H.W.; Yu, X.Q.; Xia, H.; Chen, L.; Liu, H.Y.; Zhang, A.N.; Xu, K.; Wei, H.B.; Liu, G.L.; et al. Water-saving and drought-resistance rice: From the concept to practice and theory. Mol. Breed. 2019, 39, 1–15. [Google Scholar] [CrossRef]
- Wang, L.; Yu, C.; Chen, C.; He, C.; Zhu, Y.; Huang, W. Identification of rice Di19 family reveals OsDi19-4 involved in drought resistance. Plant Cell Rep. 2014, 33, 2047–2062. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Xiong, J.; Xia, H.; Ma, X.; Gao, M.; Wang, L.; Liu, G.; Yu, X.; Luo, L. Transcriptomic divergence between upland and lowland ecotypes contributes to rice adaptation to a drought-prone agroecosystem. Evol. Appl. 2020, 13, 2484–2496. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Shiroto, Y.; Kishitani, S.; Ito, Y.; Toriyama, K. Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Rep. 2009, 28, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Lakra, N.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. A unique bZIP transcription factor imparting multiple stress tolerance in Rice. Rice 2019, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Zhang, H.; Huang, L.; Huang, L.; Li, D.; Song, F. Overexpression of a stress responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice. Front. Plant Sci. 2016, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef]
- Yuan, X.; Huang, P.; Wang, R.; HLi Lv, X.; Duan, M.; Tang, H.; Zhang, H.; Huang, J. A zinc finger transcriptional repressor confers pleiotropic effects on rice growth and drought tolerance by down-regulating stress-responsive genes. Plant Cell Physiol. 2018, 59, 2129–2142. [Google Scholar] [CrossRef]
- Tang, Y.; Bao, X.; Zhi, Y.; Wu, Q.; Guo, Y.; Yin, X.; Zeng, L.; Li, J.; Zhang, J.; He, W.; et al. Overexpression of a MYB family gene, OsMYB6, increases drought and salinity stress tolerance in transgenic rice. Front. Plant Sci. 2019, 10, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.Y. Comparative Transcriptome Analysis of Drought Tolerance and Function Characterization of OsDRAPI in Rice; Chinese Academy of Agricultural Sciences: Beijing, China, 2014. [Google Scholar]
- Gu, J.; Yin, X.; Struik, P.C.; Stomph, T.J.; Wang, H. Using chromosome introgression lines to map quantitative trait loci for photosynthesis parameters in rice (Oryza sativa L.) leaves under drought and well-watered field conditions. J. Exp. Bot. 2012, 63, 455–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.; Carvalho, I.; Faria, T.; Pinheira, C. How plants cope with water stress in the field? Photosynthesis and growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.T.; Ma, S.L.; Bai, L.P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef] [PubMed]
- Cletus, J.; Balasubramanian, V.; Vashisht Sakthivel, N.D. Transgenic expression of plant chitinases to enhance disease resistance. Biotechnol. Lett. 2013, 35, 1719–1732. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Othman Zaifuddin, F.A.M.; Hassan, N.M. Carotenoid biosynthesis regulatory mechanisms in plants. J. Oleo Sci. 2014, 63, 753–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.J.; Zhang, A.H.; Sun, J.B.; Zhang, L.X. Advances of research on allelopathic potencial of terpenoids in plants. Ecol. Environ. Sci. 2012, 21, 187–193. [Google Scholar]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones enhanced drought tolerance in plants: A coping strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Leaf Rolling Score | |||
|---|---|---|---|---|
| Day0 | Day7 | Day15 | Day21 | |
| BSN | - | - | - | + |
| MPX | - | - | - | ++ |
| Changshan-Manuo | - | - | + | ++ |
| C36-13 | - | - | + | ++ |
| Wanling1 | - | - | + | ++ |
| Wanling4 | - | - | ++ | +++ |
| Wanling8 | - | + | ++ | ++++ |
| ManPozi | - | + | +++ | +++++ |
| Changmaogu10 | - | ++ | ++++ | +++++ |
| Zinuo | - | ++ | ++++ | +++++ |
| Sample Name | Clean Reads | Total Mapped | Uniquely Mapped | Multiple Mapped | Non-Splice Reads | Splice Reads |
|---|---|---|---|---|---|---|
| CK-MPX_1 | 21,987,732 | 41,444,762 94.25% | 40,214,334 91.45% | 1,230,428 2.8% | 25,355,296 57.66% | 14,859,038 33.79% |
| CK-MPX_2 | 21,468,087 | 40,629,408 94.63% | 39,601,202 92.23% | 1,028,206 2.39% | 24,861,263 57.9% | 14,739,939 34.33% |
| CK-MPX_3 | 24,166,220 | 45,762,210 94.68% | 44,623,664 92.33% | 1,138,546 2.36% | 30,685,049 63.49% | 13,938,615 28.84% |
| D-MPX_1 | 24,485,259 | 46,524,502 95.01% | 45,551,600 93.02% | 972,902 1.99% | 28,555,559 58.31% | 16,996,041 34.71% |
| D-MPX_2 | 22,420,575 | 42,315,692 94.37% | 41,307,648 92.12% | 1,008,004 2.25% | 26,086,901 58.18% | 15,220,747 33.94% |
| D-MPX_3 | 24,358,311 | 46,429,204 95.3% | 45,500,000 93.4% | 929,204 1.91% | 29,353,793 60.25% | 16,146,207 33.14% |
| CK-BSN_1 | 32,875,666 | 61,508,950 93.55% | 59,799,194 90.95% | 1,709,756 2.6% | 38,504,957 58.56% | 21,294,237 32.39% |
| CK-BSN_2 | 29,814,310 | 56,666,020 95.03% | 55,275,174 92.7% | 1,390,846 2.33% | 35,184,648 59.01% | 20,090,526 33.69% |
| CK-BSN_3 | 23,114,058 | 42,871,330 92.74% | 41,749,732 90.31% | 1,121,598 2.43% | 26,794,172 57.96% | 14,955,560 32.35% |
| D-BSN_1 | 27,025,304 | 51,254,708 94.83% | 50,153,446 92.79% | 1,101,262 2.04% | 32,620,538 60.35% | 17,532,908 32.44% |
| D-BSN_2 | 24,713,673 | 47,035,774 95.16% | 46,077,454 93.22% | 958,320 1.94% | 29,253,860 59.19% | 16,823,594 34.04% |
| D-BSN_3 | 25,863,163 | 48,457,128 93.68% | 47,434,590 91.7% | 1,022,538 1.98% | 30,195,571 58.38% | 17,239,019 33.33% |
| TF Family | Upregulated | Downregulated | Percentage (%) | |||
|---|---|---|---|---|---|---|
| MPX_DvsMPX_CK | BSN_DvsBSN_CK | MPX_DvsMPX_CK | BSN_DvsBSN_CK | MPX_DvsMPX_CK | BSN_DvsBSN_CK | |
| Znf (RING-finger) | 6 | 22 | 6 | 16 | 20.65 | 20.69 |
| Znf | 14 | 41 | 12 | 48 | 14.13 | 20.69 |
| Znf (WRKY) | 0 | 1 | 2 | 24 | 13.59 | 3.45 |
| Aux/IAA | 1 | 16 | 2 | 6 | 11.96 | 5.17 |
| NAC | 4 | 14 | 1 | 7 | 11.41 | 8.62 |
| MYB/MYC | 5 | 17 | 5 | 1 | 9.78 | 17.24 |
| AP2/ERF | 7 | 14 | 2 | 3 | 9.24 | 15.52 |
| bZIP | 1 | 10 | 2 | 3 | 7.07 | 5.17 |
| HD-ZIP | 1 | 2 | 0 | 0 | 1.09 | 1.72 |
| HSF | 0 | 0 | 0 | 1 | 0.54 | 0.00 |
| DREB | 0 | 1 | 1 | 0 | 0.54 | 1.72 |
| Gene_ID | Description | Readcount | |
|---|---|---|---|
| CK | D | ||
| Os04g0415800 | Similar to OSIGBa0092M08.2 protein. | 0.28 | 992.69 |
| Os04g0493600 | Similar to Lectin-C precursor (PL-C). | 2.82 | 7596.22 |
| Os07g0251900 | Leucine-rich repeat, N-terminal domain-containing protein. | 829.35 | 20.05 |
| Os09g0351700 | Protein kinase, catalytic domain-containing protein. | 169.76 | 2.51 |
| Os11g0115100 | Similar to Lipid transfer protein. | 0.42 | 1909.17 |
| Os01g0910800 | Conserved hypothetical protein. | 2.58 | 3210.34 |
| Os11g0291475 | Hypothetical protein. | 2.04 | 676.33 |
| Os05g0381400 | AWPM-19-like protein, Stress tolerance through ABA-dependent pathway | 8.36 | 105.17 |
| Os01g0705200 | Late embryogenesis abundant protein repeat-containing protein. | 6.30 | 406.94 |
| Os01g0192900 | 1-aminocyclopropane-1-carboxylate synthase family protein. | 176.44 | 11.81 |
| Os06g0210900 | Lipase, class 3 family protein. | 155.16 | 6.43 |
| Os10g0505900 | Conserved hypothetical protein. | 18.46 | 828.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, X.; Zhai, N.; Yang, X.; Su, M.; Liu, C.; Wang, L.; Qu, P.; Liu, W.; Yuan, Q.; Pei, X. Identification of Drought-Resistant Genes in Shanlan Upland Rice. Agriculture 2022, 12, 150. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020150
Niu X, Zhai N, Yang X, Su M, Liu C, Wang L, Qu P, Liu W, Yuan Q, Pei X. Identification of Drought-Resistant Genes in Shanlan Upland Rice. Agriculture. 2022; 12(2):150. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020150
Chicago/Turabian StyleNiu, Xiaoling, Nanxin Zhai, Xinsen Yang, Meng Su, Caiyue Liu, Liu Wang, Pengzheng Qu, Wuge Liu, Qianhua Yuan, and Xinwu Pei. 2022. "Identification of Drought-Resistant Genes in Shanlan Upland Rice" Agriculture 12, no. 2: 150. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020150