
Structure and Function of the Soil Rhizosphere Fungal Communities in Medicinal Plants—A Preliminary Study


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Researched Plants—Ecophysiology and Medicinal Use
2.3. Soil Sample Collection
2.4. Soil Analysis
2.5. Molecular–Taxonomy Determination
2.6. Data Processing and Analysis
2.7. Statistical Analysis
3. Results
3.1. Abiotic Variables
3.2. Taxonomic Analysis
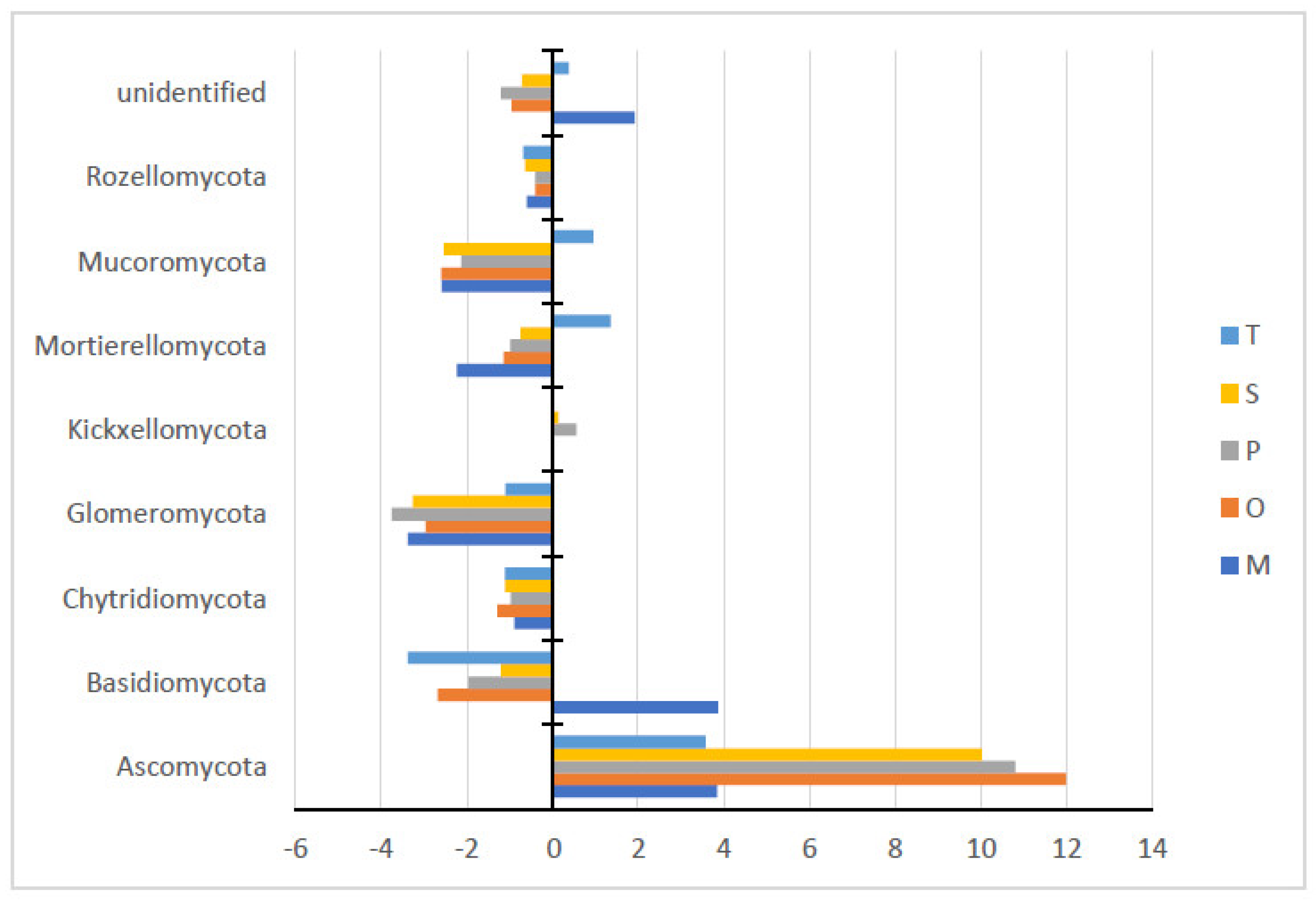
3.2.1. Phyla
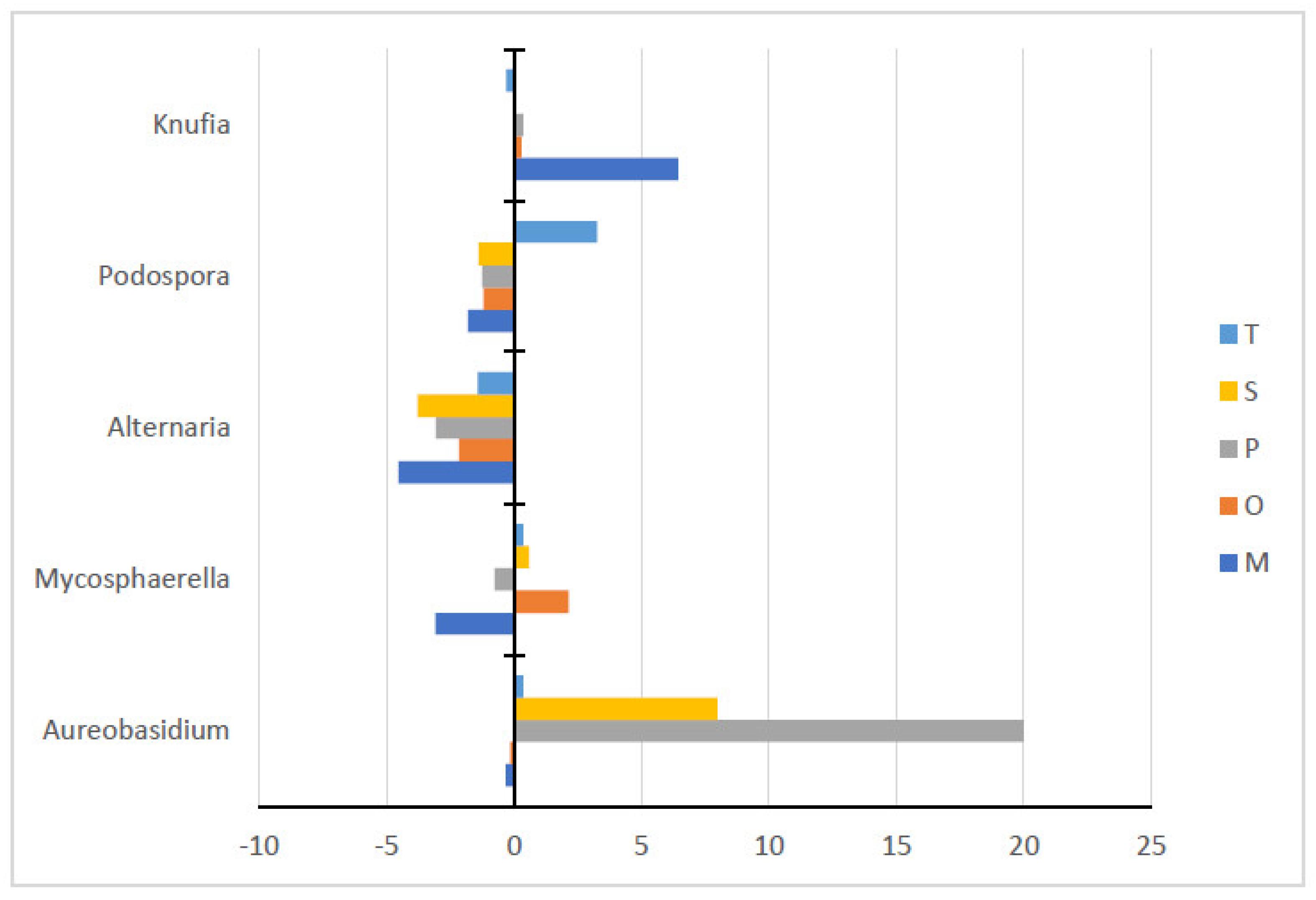
3.2.2. Genera
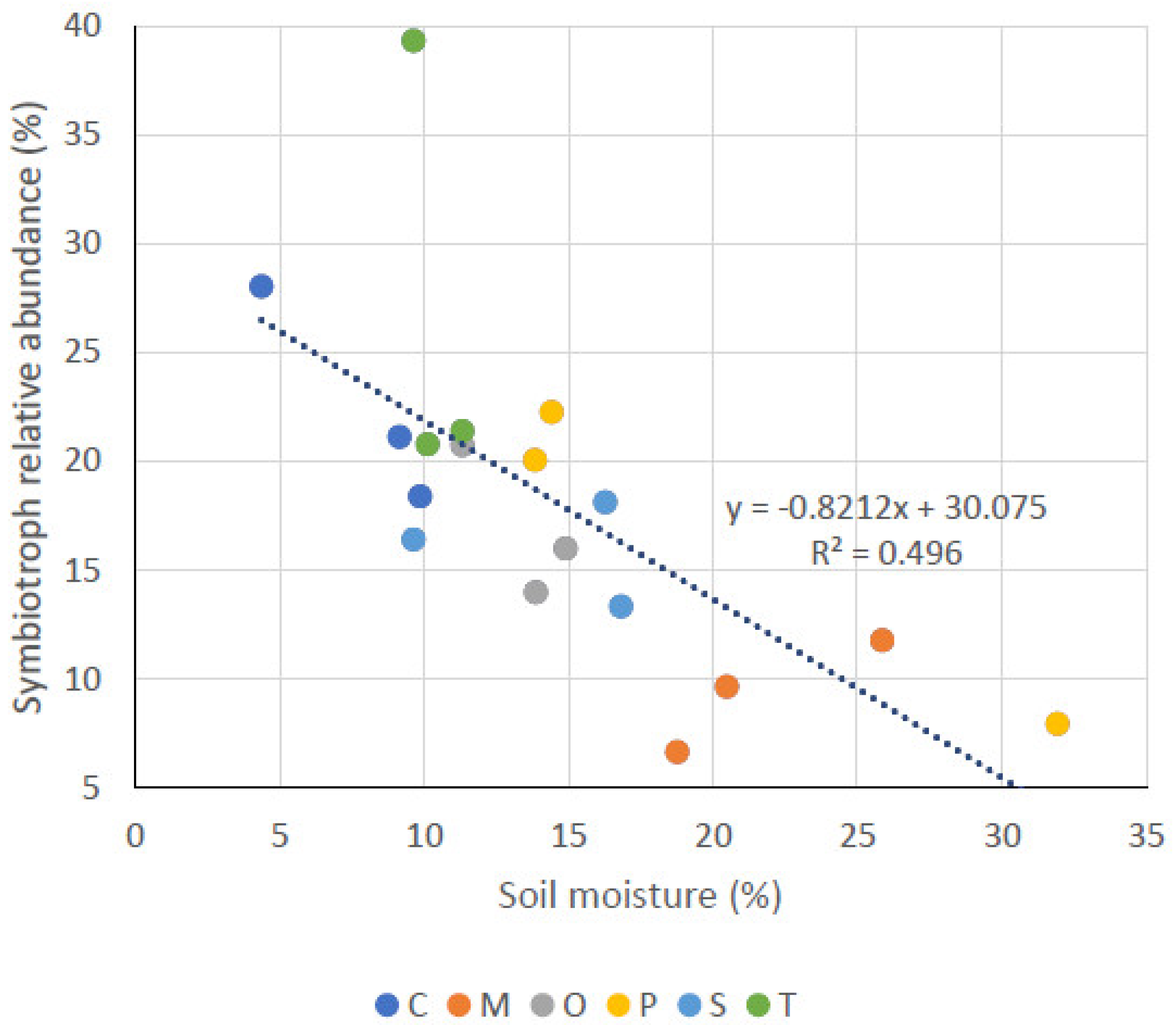
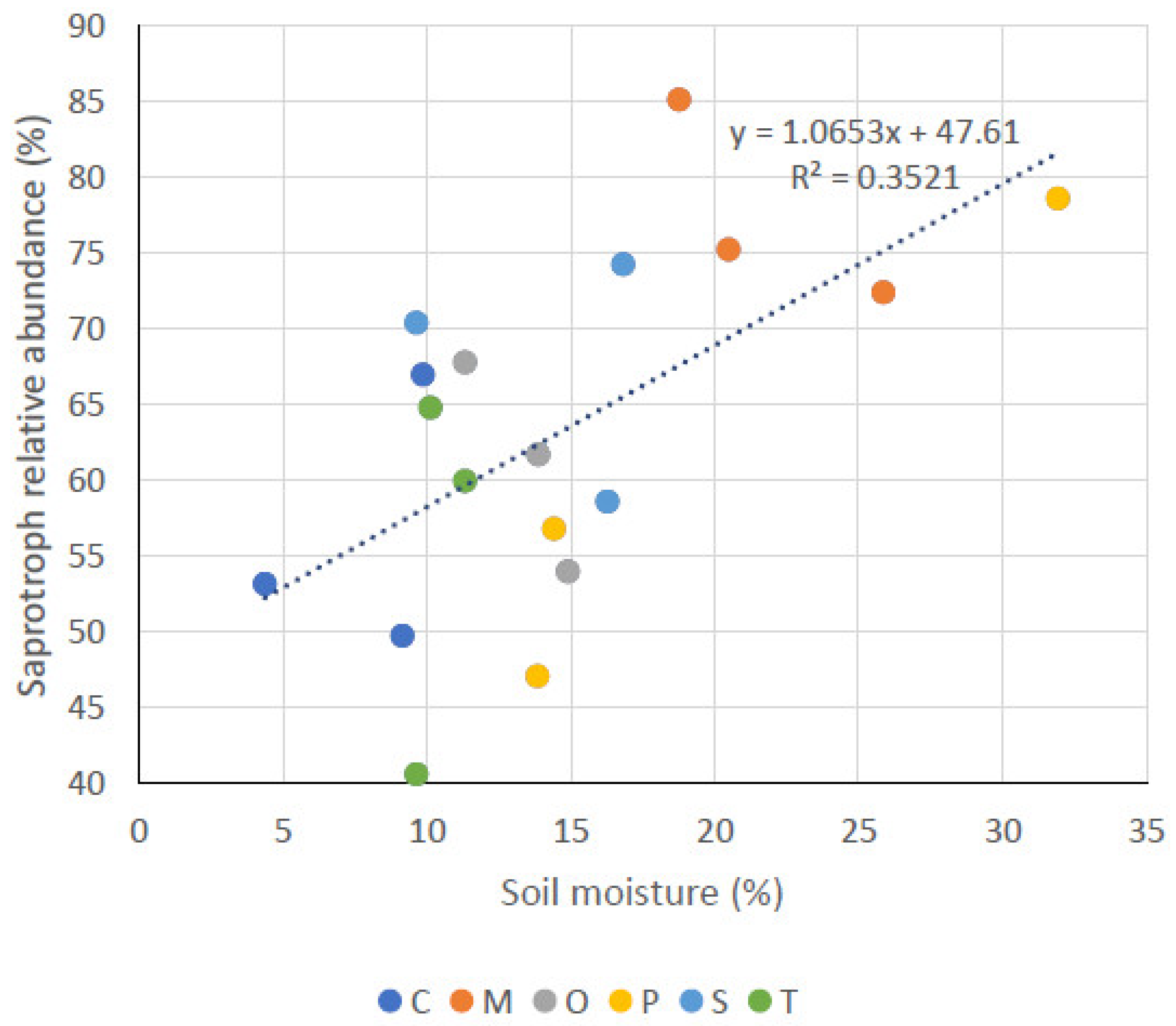
3.3. Fungal Functional Modes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Philippot, L.; Raiijmakers, J.M.; Lemanceau, P.; Van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root Exudates Regulate Soil Fungal Community Composition and Diversity. Appl. Environ. Microbiol. 2020, 74, 738–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veen, G.F.; Fry, E.L.; ten Hooven, F.C.; Kardol, P.; Morriën, E.; De Long, J.R. The Role of Plant Litter in Driving Plant-Soil Feedbacks. Front. Environ. Sci 2019, 7, 168. [Google Scholar] [CrossRef]
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Amsterdam, The Netherlands, 2008; pp. 1–8. [Google Scholar]
- van der Wal, A.; Geydan, T.D.; Kuyper, T.W.; de Boer, W. A thready affair: Linking fungal diversity and community dynamics to terrestrial decomposition processes. FEMS Microbiol. Rev. 2013, 37, 477–494. [Google Scholar] [CrossRef]
- Doehlemann, G.; Ökmen, B.; Zhu, W.; Sharon, A. Plant Pathogenic Fungi. Microbiol. Spectr. 2017, 5, 703–726. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, S.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Musilova, L.; Ridl, J.; Polivkova, M.; Macek, T.; Uhlik, O. Effects of Secondary Plant Metabolites on Microbial Populations: Changes in Community Structure and Metabolic Activity in Contaminated Environments. Int. J. Mol. Sci. 2016, 17, 1205. [Google Scholar] [CrossRef] [Green Version]
- Prajapati, N.D.; Purohit, S.S.; Sharma, A.K.; Kumar, T. A Handbook of Medicinal Plants a Complete Source Book; Jodhpur (India) Agrobios: Jodhpur, India, 2003; pp. 1–3. [Google Scholar]
- Petrovska, B.B. Historical review of medicinal plants’ usage. Pharmacogn. Rev. 2012, 6, 1–5. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Traditional Medicine Strategy: 2014–2023; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- De Smet, P.A.G.M. The Role of Plant-Derived Drugs and Herbal Medicines in Healthcare. Drugs 1997, 54, 801–840. [Google Scholar] [CrossRef]
- Köberl, M.; Schmidt, R.; Ramadan, E.M.; Bauer, R.; Berg, G. The microbiome of medicinal plants: Diversity and importance for plant growth, quality and health. Front. Microbiol. 2013, 4, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; de Boer, W.; Zhang, Y.; Ding, C.; Zhang, T.; Wang, X. Suppression of soil-borne Fusarium pathogens of peanut by intercropping with the medicinal herb Atractylodes lancea. Soil Biol. 2018, 116, 120–130. [Google Scholar] [CrossRef]
- Khan, A.L.; Asaf, S.; Abed, R.M.M.; Chai, Y.N.; Al-Rawahi, A.N.; Mohanta, T.K.; Al-Rawahi, A.; Schachtman, D.P.; Al-Harrasi, A. Rhizosphere Microbiome of Arid Land Medicinal Plants and Extra Cellular Enzymes Contribute to Their Abundance. Microorganisms 2020, 8, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villalobos-Flores, L.E.; Espinosa-Torres, S.D.; Hernández-Quiroz, F.; Piña-Escobedo, A.; Cruz-Narváez, Y.; Velázquez-Escobar, F.; Süssmuth, R.; García-Mena, J. The Bacterial and Fungal Microbiota of the Mexican Rubiaceae Family Medicinal Plant Bouvardia ternifolia. Microb. Ecol. 2021. [Google Scholar] [CrossRef]
- Kushwaha, R.K.; Rodrigues, V.; Kumar, V.; Patel, H.; Raina, M.; Kumar, D. Soil Microbes-Medical Plants Interactions: Ecological Diversity and Future Prospects. In Plant Microbiome Symbiosis; Varma, A., Tripathi, S., Prasad, R., Eds.; Springer: Cham, Switzerland, 2020; pp. 263–286. [Google Scholar]
- Feinburn-Dothan, N.; Danin, A. Analytical Flora of Eretz-Israel, 2nd ed.; CANA Publishing House Ltd.: Jerusalem, Israel, 1998; p. 583. [Google Scholar]
- Mayer-Chissick, U.; Lev, E. Wild Edible Plants in Israel Tradition Versus Cultivation. In Medicinal and Aromatic Plants of the Middle-East; Yaniv, Z., Dudai, N., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 12–14. [Google Scholar]
- Dudai, N.; Yaniv, Z. Endemic Aromatic Medicinal Plants in the Holy Land Vicinity. In Medicinal and Aromatic Plants of the Middle-East; Yaniv, Z., Dudai, N., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 37–44. [Google Scholar]
- Perfumi, M.; Arnold, N.; Tacconi, R. Hypoglycemic activity of Salvia fruticose Mill. From Cyprus. J. Ethnopharmacol. 1991, 34, 135–140. [Google Scholar] [CrossRef]
- Boukhary, R.; Raafat, K.; Ghoneim, A.I.; Aboul-Ela, M.; El-Lakany, A. Anti-Inflammatory and Antioxidant Activities of Salvia fruticosa: An HPLC Determination of Phenolic Contents. Evid. Based Complement. Altern. Med. 2016, 2016, 7178105. [Google Scholar] [CrossRef] [Green Version]
- Nawash, O.; Shudiefat, M.; Al-Tabini, R.; Al-Khalidi, K. Ethnobotanical study of medicinal plants commonly used by local Bedouins in the badia region of Jordan. J. Ethnopharmacol. 2013, 148, 921–925. [Google Scholar] [CrossRef]
- Maccioni, A.; Falconieri, D.; Porcedda, S.; Piras, A.; Gonçalvese, M.J.; Alves-Silva, J.M.; Salgueiro, L.; Maxia, A. Antifungal activity and chemical composition of the essential oil from the aerial parts of two new Teucrium capitatum L. chemotypes from Sardinia Island, Italy. Nat. Prod. Res. 2020, 35, 6007–6013. [Google Scholar] [CrossRef]
- Ghareeb, B.A.; Weld-Ali, D.H. Assessment of antibacterial virtues of Teucrium capitatum (Ja’da) on pathogenic bacteria (From Folk to Complementary Medicine). IJMPAM 2015, 3, 1–9. [Google Scholar]
- Chabane, S.; Boudjelal, A.; Keller, M.; Doubakh, S.; Potterat, O. Teucrium polium—Wound healing potential, toxicity and polyphenolic profile. S. Afr. J. Bot. 2020, 137, 228–235. [Google Scholar] [CrossRef]
- Yaniv, Z.; Dudai, N. (Eds.) Myrtle (Myrtus communis L.)—A Native Mediterranean and Cultured Crop Species. In Medicinal and Aromatic Plants of the Middle-East; Springer: Dordrecht, The Netherlands, 2014; pp. 253–267. [Google Scholar]
- Alipour, G.; Dashti, S.; Hosseinzadeh, H. Review of Pharmacological Effects of Myrtus communis L. and its Active Constituents. Phytother. Res. 2014, 28, 1125–1136. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Milella, F.; Mari, M.; Roberti, R. A preliminary investigation into Aureobasidium pullulans as a potential biocontrol agent against Phytophthora infestans of tomato. Biol. Control. 2017, 114, 144–149. [Google Scholar] [CrossRef]
- Landau, S.; Muklada, H.; Markovics, A.; Azaizeh, H. Traditional Uses of Pistacia lentiscus in Veterinary and Human Medicine. In Medicinal and Aromatic Plants of the Middle-East; Yaniv, Z., Dudai, N., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 163–197. [Google Scholar]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoch, C.L.; Sung, G.H.; López-Giráldez, F.; Townsend, J.P.; Miadlikowska, J.; Hofstetter, V.; Robbertse, B.; Matheny, P.B.; Kauff, F.; Wang, Z.; et al. The Ascomycota tree of life: A phylum-wide phylogeny clarifies the origin and evolution of fundamental reproductive and ecological traits. Syst. Biol. 2009, 58, 224–239. [Google Scholar] [CrossRef] [PubMed]
- Bozoudi, D.; Tsaltas, D. The Multiple and Versatile Roles of Aureobasidium pullulans in the Vitivinicultural Sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Di Francesco, A.; Sciubba, L.; Becivenni, M.; Marzadori, C.; Baraldi, E. Application of Aureobasidium pullulans in iron-poor soil. Can the production of siderophores improve iron bioavailability and yeast antagonistic activity? Ann. Appl. Biol. 2021. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J. Alternaria spp.: From general saprophyte to specific parasite. BSPP 2003, 4, 225–236. [Google Scholar]
- Soković, M.; Tzakou, O.; Pitarokili, D.; Couladis, M. Antifungal activities of selected aromatic plants growing wild in Greece. Mol. Nutr. Food Res. 2002, 46, 317–320. [Google Scholar]
- Kordali, S.; Usanmaz, A.; Cakir, A.; Komaki, A.; Ercisli, S. Antifungal and herbicidal effects of fruit essential oils of four Myrtus communis genotypes. Chem. Biodivers. 2016, 13, 77–84. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E. Growth of saprotrophic fungi and bacteria in soil. FEMS Microbiol. Ecol. 2011, 78, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Azcón-Aguilar, C.; Palenzuela, J.; Roldán, A.; Bautista, S.; Vallejo, R.; Bera, J.M. Analysis of the mycorrhizal potential in the rhizosphere of representative plant species from desertification-threatened Mediterranean shrublands. Appl. Soil Ecol. 2003, 22, 29–37. [Google Scholar] [CrossRef]
- Marouf, B.K.; Ighilhariz, Z.; de Lajudie, P.; Duponnois, R.; Bekki, A. Assessing the Native Arbuscular Mycorrhizal Symbioses to Rehabilitate a Degraded Coastal Sand Dune In Algeria. Int. J. Agric. Crop Sci. 2015, 8, 194–202. [Google Scholar]
- Alguacil, M.M.; Torres, M.P.; Torrecillas, E.; Díaz, G.; Roldán, A. Plant type differently promote the arbuscular mycorrhizal fungi biodiversity in the rhizosphere after revegetation of a degraded, semiarid land. Soil Biol. Biochem. 2011, 43, 167–173. [Google Scholar] [CrossRef]
- Vaz, A.B.M.; Sampedro, I.; Siles, J.A.; Vasquez, J.A.; García-Romera, I.; Vierheilig, H.; Roas, C.A.; Ocampo, J.A. Arbuscular mycorrhizal colonization of Sorghum vulgare in presence of root endophytic fungi of Myrtus communis. Appl. Soil Ecol. 2012, 61, 288–294. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Control | Sampled Rhizosphere | ||||
|---|---|---|---|---|---|---|
| O. syriacum | S. fruticosa | T. capitatum | M. communis | P. lentiscus | ||
| SM (%) | 7.83 ± 2.99 b | 13.37 ± 1.86 ab | 14.23 ± 04.02 ab | 10.33 ± 0.87 b | 21.73 ± 03.71 a | 20.07 ± 10.25 a |
| OM (%) | 6.37 ± 10.84 a | 8.40 ± 0.66 a | 7.0 ± 01.57 a | 7.67 ± 2.57 a | 10.07 ± 5.84 a | 16.57 ± 12.11 a |
| pH | 8.0 ± 0.06 ab | 8.10 ± 0.05 a | 7.91 ± 0.07 bc | 7.92 ± 0.06 bc | 7.91 ± 0.08 bc | 7.79 ± 0.09 c |
| EC | 78.57 ± 1.18 a | 84.77 ± 12.18 a | 83.43 ± 04.65 a | 86.3 ± 6.59 a | 80.37 ± 05.06 a | 83.93 ± 20.40 a |
| Phyla | Control | Sampled Rhizosphere | ||||
|---|---|---|---|---|---|---|
| O. syriacum | S. fruticosa | T. capitatum | M. communis | P. lentiscus | ||
| Asco | 76.52 ± 8.54 b | 88.51 ± 1.0 a | 86.55 ± 6.95 ab | 80.1 ± 6.3 ab | 80.37 ± 05.5 ab | 87.33 ± 3.89 ab |
| Basidio | 8.87 ± 5.33 a | 6.20 ± 0.72 a | 7.67 ± 5.13 a | 5.49 ± 1.54 a | 12.74 ± 5.08 a | 6.91 ± 1.41 a |
| Chytridio | 1.36 ± 1.28 a | 0.07 ± 0.06 b | 0.26 ± 0.31 ab | 0.25 ± 0.14 ab | 0.48 ± 0.7 ab | 0.40 ± 0.32 ab |
| Glomero | 5.07 ± 3.69 a | 2.11 ± 1.03 ab | 1.81 ± 0.47 ab | 3.98 ± 1.41 ab | 1.69 ± 1.45 ab | 1.32 ± 1.39 b |
| Kickxello | 0.0 ± 0.0 b | 0.01 ± 0.02 b | 0.15 ± 0.02 b | 0.0 ± 0.0 b | 0.01 ± 0.02 b | 0.55 ± 0.45 a |
| Mortierel | 2.81 ± 2.58 a | 1.67 ± 1.23 a | 2.07 ± 2.4 a | 4.16 ± 2.55 a | 0.57 ± 0.33 a | 1.83 ± 1.23 a |
| Mucoro | 2.63 ± 3.31 a | 0.03 ± 0.04 a | 0.1 ± 0.14 a | 3.57 ± 5.73 a | 0.05 ± 0.05 a | 0.51 ± 0.32 a |
| Rozello | 0.75 ± 0.77 a | 0.35 ± 0.58 a | 0.12 ± 0.15 a | 0.07 ± 0.06 a | 0.15 ± 0.24 a | 0.36 ± 0.62 a |
| unid | 2.0 ± 1.21 ab | 1.04 ± 0.55 b | 1.28 ± 0.45 b | 2.37 ± 1.31 ab | 3.92 ± 1.98 a | 0.79 ± 0.57 b |
| Genera | Control | Sampled Rhizosphere | ||||
|---|---|---|---|---|---|---|
| O. syriacum | S. fruticosa | T. capitatum | M. communis | P. lentiscus | ||
| Aureobasi | 0.43 ± 0.22 b | 0.30 ± 0.19 b | 8.40 ± 13.96 ab | 0.80 ± 0.42 b | 0.10 ± 0.11 b | 20.42 ± 20.43 a |
| Mycospha | 4.17 ± 3.66 ab | 6.31 ± 3.79 a | 4.77 ± 0.62 ab | 4.54 ± 1.96 ab | 1.09 ± 0.5 b | 3.44 ± 2.91 ab |
| Alternaria | 4.73 ± 4.12 a | 2.59 ± 1.19 ab | 0.94 ± 0.35 b | 3.32 ± 0.35 ab | 0.21 ± 0.14 b | 1.68 ± 1.59 ab |
| Podospora | 2.27 ± 1.5 ab | 1.07 ± 0.37 b | 0.88 ± 0.99 b | 5.55 ± 4.69 a | 0.48 ± 0.71 b | 1.01 ± 1.55 b |
| Knufia | 0.3 ± 0.52 b | 0.60 ± 0.95 b | 0.28 ± 0.22 b | 0.0 ± 0.0 b | 6.74 ± 2.72 a | 0.66 ± 0.77 b |
| Trophic Group | Control | Sampled Rhizosphere | ||||
|---|---|---|---|---|---|---|
| O. syriacum | S. fruticosa | T. capitatum | M. communis | P. lentiscus | ||
| PA | 20.9 ± 7.47 a | 22.0 ± 9.53 a | 16.33 ± 6.04 a | 17.77 ± 2.91 a | 13.13 ± 4.2 a | 22.43 ± 9.79 a |
| SA | 56.63 ± 9.15 b | 61.17 ± 6.92 ab | 67.73 ± 8.13 ab | 55.10 ± 12.79 b | 77.57 ± 6.67 a | 60.83 ± 16.13 ab |
| SY | 22.5 ± 4.95 a | 16.87 ± 3.48 ab | 15.93 ± 2.43 ab | 27.17 ± 10.51 a | 9.30 ± 2.56 b | 16.73 ± 7.74 ab |
| SM (%) | OM (%) | pH | EC (µS* cm−1) | SA | PA | SY | |
|---|---|---|---|---|---|---|---|
| SM (%) | - | ||||||
| OM (%) | 0.75 ** | - | |||||
| pH | NS | NS | - | ||||
| EC (µS* cm−1) | NS | 0.58 | NS | - | |||
| SA | 0.59 * | NS | NS | NS | - | ||
| PA | NS | NS | NS | NS | −0.79 ** | - | |
| SY | −0.71 * | NS | NS | NS | −0.83 *** | NS | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Applebaum, I.; Jeyaraman, M.; Sherman, C.; Doniger, T.; Steinberger, Y. Structure and Function of the Soil Rhizosphere Fungal Communities in Medicinal Plants—A Preliminary Study. Agriculture 2022, 12, 152. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020152
Applebaum I, Jeyaraman M, Sherman C, Doniger T, Steinberger Y. Structure and Function of the Soil Rhizosphere Fungal Communities in Medicinal Plants—A Preliminary Study. Agriculture. 2022; 12(2):152. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020152
Chicago/Turabian StyleApplebaum, Itaii, Mareeswaran Jeyaraman, Chen Sherman, Tirza Doniger, and Yosef Steinberger. 2022. "Structure and Function of the Soil Rhizosphere Fungal Communities in Medicinal Plants—A Preliminary Study" Agriculture 12, no. 2: 152. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture12020152